 María Alejandra Gleisner1,2
María Alejandra Gleisner1,2 Mariela Navarrete1,2
Mariela Navarrete1,2 Francisca Hofmann1,2
Francisca Hofmann1,2 Flavio Salazar-Onfray1,2*
Flavio Salazar-Onfray1,2* Andrés Tittarelli1,2*
Andrés Tittarelli1,2*
- 1Disciplinary Program of Immunology, Faculty of Medicine, Institute of Biomedical Sciences, Universidad de Chile, Santiago, Chile
- 2Faculty of Medicine, Millennium Institute on Immunology and Immunotherapy, Universidad de Chile, Santiago, Chile
Gap junctions (GJs)-mediated intercellular communications (GJICs) are connexin (Cx)-formed plasma membrane channels that allow for the passage of small molecules between adjacent cells, and are involved in several physiopathological processes, including immune responses against cancer. In general, tumor cells are poorly coupled through GJs, mainly due to low Cx expression or reduced channel activity, suggesting that Cxs may have tumor suppressor roles. However, more recent data indicate that Cxs and/or GJICs may also in some cases promote tumor progression. This dual role of Cx channels in tumor outcome may be due, at least partially, to the fact that GJs not only interconnect cells from the same type, such as cancer cells, but also promote the intercellular communication of tumor cells with different types of cells from their microenvironment, and such diverse intercellular interactions have distinctive impact on tumor development. For example, whereas GJ-mediated interactions among tumor cells and microglia have been implicated in promotion of tumor growth, tumor cells delivery to dendritic cells of antigenic peptides through GJs have been associated with enhanced immune-mediated tumor elimination. In this review, we provide an updated overview on the role of GJICs in tumor immunity, focusing on the pro-tumor and antitumor effect of GJs occurring among tumor and immune cells. Accumulated data suggest that GJICs may act as tumor suppressors or enhancers depending on whether tumor cells interact predominantly with antitumor immune cells or with stromal cells. The complex modulation of immune-tumor cell GJICs should be taken into consideration in order to potentiate current cancer immunotherapies.
Introduction
Gap junctions (GJs) are intercellular channels found at the plasma membrane that allow direct communication between adjacent cells. Functional GJs are composed of connexin (Cx) proteins. Cxs form hexameric hemichannels (Cx-HCs) inserted into the membrane of one cell, which then docks with a Cx-HC from an adjacent cell to establish a GJ channel (1). When Cx-HCs acquire an open conformation, they allow for the bidirectional exchange of molecules between the cytoplasm and the extracellular fluid. The Cx gene family is comprised of 21 members in humans and 20 members in mice, and they are usually named after their predicted molecular weight, for example Cx43 corresponds to a Cx of 43 kDa. Whereas most Cx isoforms are strictly expressed in a tissue-specific manner, Cx43 is expressed almost ubiquitously, and is the main Cx member in the immune system (2). Each Cx-HC can be formed by one or more isoform of Cx proteins, which determines, at least partially, GJ permeability and their regulatory properties (1). GJs and Cx-HCs allow for the intercellular passage and the intake/uptake from or to the extracellular fluid of small (~1.4 nm) and immunologically relevant molecules, including adenosine triphosphate (ATP), cyclic adenosine monophosphate (cAMP), uridine diphosphate (UDP), cyclic guanosine monophosphate–adenosine monophosphate (cGAMP), inositol triphosphate (IP3), Ca2+, microRNAs (miRNAs), and small peptides (3).
Gap junction-mediated intercellular communications (GJICs) are critical for several physiological processes, including: electric current propagation in the heart and neurons (4, 5); embryonic development (6); cell differentiation (7); tissue homeostasis (8); autophagosome biogenesis (9); cell survival, proliferation, and cell death (10, 11); and the immune response (12). As GJs are involved in countless cellular and physiological processes, the cells need to establish delicate regulatory mechanisms of GJICs, which occurs at different levels, such as that of Cx gene expression, the life cycle of Cx protein level, or GJ assembly and permeability. These different mechanisms of GJIC regulation are at the same time highly responsive to environmental cues, including pro-inflammatory signals. Excellent reviews about regulatory mechanisms of Cx expression and GJIC have been recently published (13–21).
Mutations in Cx genes or loss of Cx channel functionality have been implicated in the development of different diseases, such as congenital deafness (22), skin disorders (23), cardiac arrhythmias (24), cataracts (25), and cancer (26). The role of Cx channels in the incidence and progression of cancer has been extensively investigated since the year 1966 when Loewensteind and Kanno showed that the electrical coupling found in normal hepatocytes was lost in liver tumors (27). A substantial progress in our understanding of GJ-mediated cell coupling in cancer has occurred since then, and it was recently reviewed (26, 28, 29). In general, cancer cells derived from various tumor types show reduced expression of Cxs and low GJ cell coupling, leading to the concept that Cxs are tumor suppressor genes, principally due to the antiproliferative effect of their overexpression in tumor cells (30). However, recent evidence also indicates that this is partially true and depends on cancer type, disease stages, and Cx isotype (28). Indeed, an analysis of different clinical studies for 15 different cancer types indicates that the expression of Cxs in tumor biopsies could be associated with good or bad prognosis of cancer progression, depending on the Cx isotype and the type of cancer (Table 1).
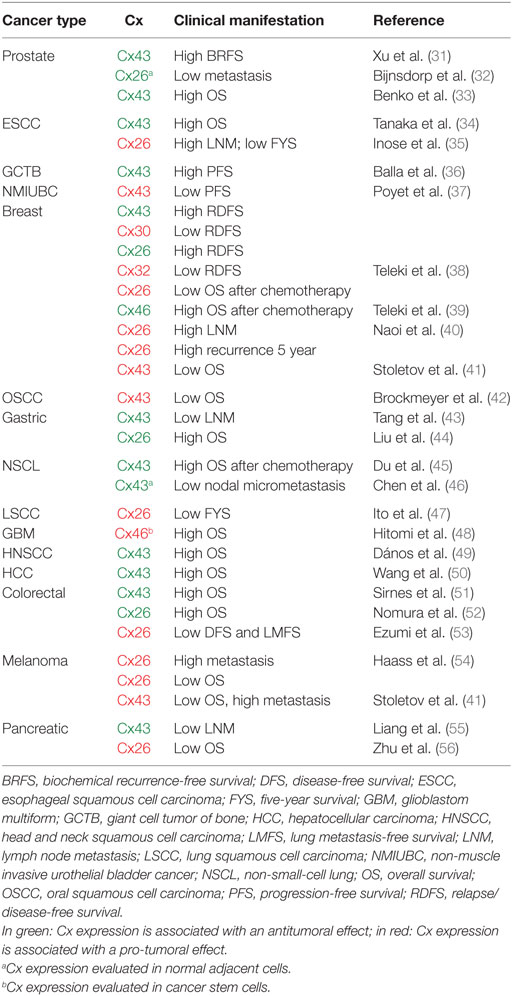
Table 1. Associations between connexin (Cx) expression in tumors and clinical outcome in cancer patients.
Moreover, accumulated evidences strongly suggest that diverse aspects of the functionality of Cxs could differentially impact tumor progression: (i) besides their role as channel forming units, Cxs have channel-independent activities that may affect tumor cell growth (29, 57); (ii) Cx-HCs have differential roles than Cx-GJs in cancer cells (58); and (iii) Cxs can mediate the formation of homotypic GJICs among tumor cells, and/or the formation of heterotypic GJICs between different cell types within the tumor microenvironment, which could differentially impact the tumor cells fate. In this review article, we summarize recent data describing the pro- and antitumor effects of the heterotypic GJICs that tumor cells form (either with immune and non-immune cells), focusing on the role of GJICs in the antitumor immune response. A large number of evidences suggest that Cx expression on tumor cells may promote or halt cancer progression depending on the type of cells engaged at the tumor microenvironment. In general, tumor GJICs with immune cells may promote responses against tumors, while tumor interactions with some stromal cells through GJs may inhibit or enhance tumor cell growth depending on the particular context where those interactions occur.
Heterotypic GJICs among Cancer Cells and Immune Cells: Role of GJ in Antitumoral Immunity
Almost all immune cells and their hematopoietic precursors express Cx proteins, and nowadays its ability to modulate different aspects of immune responses is well recognized (2, 12, 59, 60). Cx channels and GJICs have been implicated in hematopoiesis, hemostasis, phagocytosis, immune cell migration, lymphocyte responses, antigen (Ag) cross-presentation, inflammation, immune tolerance, and cancer immunity. In Table 2, we summarize the current information available on the role of Cxs, Cx-HCs, and GJICs in the immune system activities. Here, we summarize recent data describing immune cell–cancer cell heterotypic GJICs that negatively impact tumor progression (Figure 1, right panel).
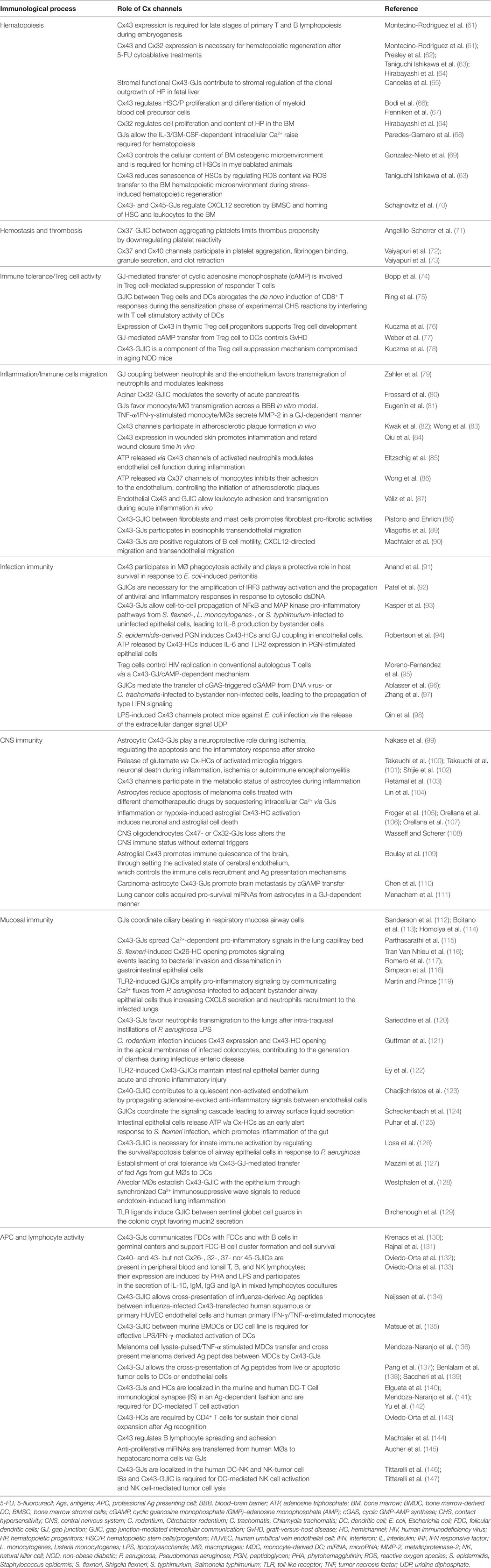
Table 2. Summary of the role of connexin (Cx) channels in immunity.
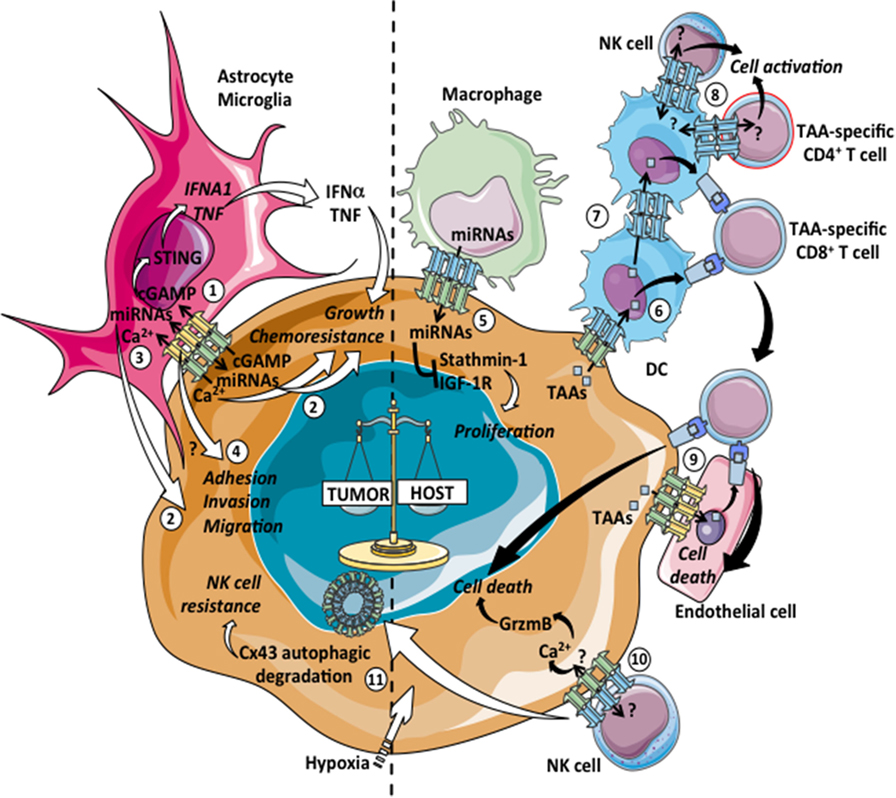
Figure 1. Pro- and antitumoral effects of tumor-immune cell heterotypic GJICs. Gap junction (GJ)-mediated communications among tumor cells and immune cells can lead to pro-tumoral (left: 1–4, 11) or antitumoral (right: 5–10) consequences. 1: carcinoma–astrocyte interactions promote brain metastasis of breast and lung cancers through the passage of the second messenger cyclic guanosine monophosphate–adenosine monophosphate (cGAMP) from tumor cells to astrocytes via Cx43-GJs, the subsequent activation of the STING pathway and the production of IFN-α and TNF that act as paracrine signals promoting growth and chemoresistance of tumor cells. 2: GJ-mediated diffusion of pro-survival microRNAs (miRNAs) between mouse astrocytes and human lung cancer cells provides increased resistance to chemotherapy. Similarly, the transfer of miRNAs from glioma to astrocytes induces glioma invasion. 3: astrocytes reduce apoptosis in melanoma cells treated with different chemotherapeutic drugs by sequestering intracellular Ca2+ via GJICs. 4: GJICs among glioblastoma cells and astrocytes contribute through uncharacterized mechanisms to the adhesion, migration, and invasion of tumor cells to the brain parenchyma. 5: GJ-mediated transfer of miRNAs from macrophages to hepatocellular carcinoma cell lines regulates gene expression of stathmin-1 and insulin-like growth factor-1 receptor and inhibits tumor cell proliferation. 6: Cx43 expression in melanoma cells allows for the transfer of preprocessed tumor associated antigens (TAAs) from melanoma cells to dendritic cells (DCs), improving DC-based tumor vaccination by increasing CD8+ T cell activation and antitumor immunity. 7: Cx43-GJs participate in melanoma antigen transfer and cross-presentation between human DCs, facilitating a more effective DC-mediated CD4+ T cell activation. 8: Cx43-GJs accumulate at the immunological synapse (IS) formed between DCs and melanoma-specific CD4+ T cells and natural killer (NK) cells, contributing to cell activation. 9: Cx43-GJs allow for the passage of TAA peptides from melanoma to autologous endothelial cells, inducing their cross-recognition and elimination by TAA-specific CD8+ T cells. 10: Cx43-GJs accumulate at the lytic IS formed between NK cells and melanoma cells, contributing to Ca2+ influx and granzyme-b (GrzmB)-mediated induction of apoptosis in the target cells. 11: Activation of autophagy in hypoxic melanoma cells causes the selective degradation of GJ-Cx43, impairing NK cell-mediated tumor cell killing.
Initial studies exploring a potential antitumoral role of GJICs among cancer cells and immune cells were encouraged by the seminal work of Jacques Neefjes and collaborators in 2005, where the transfer and cross-presentation of viral Ag peptides via GJs was reported (134). In this study, it was shown that GJ-negative human squamous carcinoma cells (A431 cell line) transferred micro-injected 9-mer linear peptides to surrounding non-micro-injected cells only when the cells were stably transfected with the human Cx43 gene. The closure of GJs by 2-aminoethoxydiphenyl borate prevented this intercellular peptide transfer. Neijssen and coworkers also evaluated the Cx43-GJ-mediated transfer of endogenous and immunologically relevant Ag peptides. They showed that human primary human leukocyte Ag (HLA)-A2+ monocytes stimulated with interferon (IFN)-γ and tumor necrosis factor (TNF)-α (cytokines that induce Cx43 expression) efficiently acquired influenza-derived Ag peptides (FluM57–65) from influenza-infected cells (A431 or endothelial cells) via Cx43-GJs, allowing for the subsequent monocyte-mediated cross-priming of an HLA-A2-restricted FluM57–65-specific T-cell clone (134).
Later, our group described that melanoma Ag peptides could also be transferred and cross-presented between human dendritic cells (DCs) via Cx43-GJs (136). In this work, we reported that melanoma patient’s HLA-A2- monocyte-derived DCs incubated overnight with an allogeneic melanoma cell lysate (MCL), efficiently transfer MelanA/MART127–35 peptides to HLA-A2+ monocyte-derived DCs, leading to the subsequent activation of an HLA-A2-restricted MelanA/MART127–35-specific cytotoxic T lymphocyte (CTL) clone (136). The transfer of MelanA/MART127–35 peptides between DCs was strongly prevented by two different GJ chemical inhibitors (oleamide and 18β-glycyrrhetinic acid) or by a Cx43 inhibitor mimetic peptide, indicating the involvement of Cx43-GJs in the cross-presentation of tumor associated Ags (TAAs) in human DCs. Of note, in a series of clinical trials, these MCL-DCs were used as an antitumor immunotherapy for advanced malignant melanoma patients (148–153). In this series of studies, a positive correlation between the immune response induced by MCL-DC-vaccination, as established by a patient tumor-specific delayed-type IV hypersensitivity reaction and improved long-term survival was reported. In this context, it has been suggested that the efficient clinical effect of adoptively transferred DC vaccines may be improved by their potential to interact with local DCs in vivo and/or other cell types in peripheral tissues and lymph nodes (154). These interactions could include GJ-mediated Ag transfer and cross-presentation from injected DCs to local DCs, which may finally amplify the Ag-specific DC-mediated T cell activation. Indeed, it was reported in a murine model, that the OVA257–264 Ag peptide transfer from OVA-expressing DC vaccines to endogenous professional Ag presenting cells (APCs) was required for efficient OVA257–264-specific CD8+ T cell priming (155). Although in these studies the GJ-mediated transfer of Ag peptides was not evaluated, the group of Rescigno, in a murine model of oral tolerance to fed Ags, elegantly showed that Cx43-GJ-mediated transfer of Ag peptides between gut resident APCs occurs in vivo (127). These observations strongly suggest that the spreading of Ag peptides between cells by GJs could be a general mechanism of Ag cross-presentation.
In the context of tumor immunity, very interesting findings suggest that Cx43-mediated transfer of Ag peptides from melanoma cells to DCs could be the major mechanism of tumor Ag cross-presentation occurring in vivo (139). This mechanism allows DCs infiltrating Salmonella-infected Cx43 positive melanoma tumors to acquire preprocessed Ag peptides from the cancer cells, leading to the activation of tumor Ag-specific CTLs that finally eliminate distal tumors. The authors showed that this antitumor immune response against distal tumors was strongly abrogated when the Salmonella-treated tumor cells were silenced for Cx43 (139). Moreover, DC vaccines loaded ex vivo with Salmonella-infected B16 melanoma cells were more efficient in inducing melanoma growth inhibition in vaccinated mice compared to other types of DC vaccines, but only when Cx43 was not silenced in the Salmonella-infected B16 cells used for loading DC vaccines. These results indicate that transfer of TAA peptides from tumor cells to DCs through Cx43-GJs is far more effective than standard pathways of DC Ag-loading in generating protective DC-based vaccines (139). Interestingly, DCs can also acquire tumor-derived Ag peptides by GJ coupling with apoptotic tumor cells, as suggested by the findings of Pang and coworkers (137). This could be of great relevance in the immune response against tumors, since caspase activation can expose neo epitopes in early apoptotic tumor cells through the direct cleavage of proteins, which results in epitopes from these proteins being favored for cross-presentation, and thus amplifying the repertoire of cross-presented Ags (156). Additionally, in a 3D in vitro cell coculture model, Cx43-GJs have been implicated in the Ag peptide transfer from melanoma to autologous endothelial cells. Once endothelial cells acquire the melanoma Ag peptides, they become susceptible to cross-recognition and elimination by an autologous tumor-specific CTL clone (138). In fact, Cx43-GJs can be detected among melanoma and endothelial cells in metastatic biopsies from patients (157), suggesting that CTL-mediated elimination of endothelial cells may contribute to control tumor progression, which needs further investigation.
In addition to peptides, the GJ-mediated transfer of miRNAs between tumor cells and immune cells have been implicated in tumor immunity. Specifically, Aucher and collaborators (145) reported that miR-142 and miR-223, which are endogenously expressed in human macrophages (MØs) but not in hepatocarcinoma cells (HCCs), were transferred from MØs to HCC cells via GJs and effectively target the expression of stathmin-1 and insulin-like growth factor-1 receptor in the acceptor tumor cells leading to the inhibition of tumor cell proliferation.
Furthermore, recent evidences suggest that Cx43 is a component of the immune synapse, and that Cx43-GJICs are required for Ag-dependent DC-mediated T cell activation (140, 141). In this context, we reported that Cx43 channels (both GJs and HCs) accumulate at the immunological synapse (IS) during DC-mediated Ag-specific CD4+ T cell priming, mediating the bidirectional crosstalk between DCs and T cells. This phenomenon was observed in both murine (DC-OVA/OT-II T cells) and human (MCL-DCs/melanoma-specific autologous CD4+ T cell clone) models. The evidence indicated that Cx43-GJICs between DCs and T cells regulates Ca2+ signals and DC-mediated T cell activation (141), pointing to a role for Cx43 as an important functional component for intercellular signaling in the immune system. Similarly, Cx43 accumulation was detected at the interface of mature human DCs and autologous resting natural killer (NK) cells, mediating bidirectional GJICs between these cells. The blockade of Cx43-GJs strongly inhibits the DC-mediated activation of NK cells, as measured by NK cell expression of CD69 and CD25 and the secretion of IFN-γ (146). The nature of the molecules shuttled via Cx43 channels at the IS between DCs and lymphocytes (both T cells and NK cells) remains uncharacterized, but as lymphocyte activation requires ATP and Ca2+ for biomass synthesis and signal transduction (158, 159), both molecules are reasonable candidates to be mobilized from DCs to lymphocytes by Cx43-GJs.
Moreover, Cx43 channels seem to accumulate at the interface of NK cells and target tumor cells (myelogenous leukemia or melanoma cells) and to mediate intercellular communications that participate in NK cell-mediated tumor cell lysis (146, 147). Cx43-GJICs among NK cells and tumor cells appear to not affect tumor-induced NK cell degranulation but instead do control the NK cell cytotoxicity by contributing to granzyme-b activity and Ca2+ influx into tumor cells (146, 147). Moreover, Cx43 expression in target tumor cells renders these cells more susceptible to NK cell-mediated lysis. Indeed, Cx43 gene knockdown in Cx43 positive tumor cells decreases the level of NK cell-mediated lysis to the same extent as the prevention of GJICs by chemical inhibitors or Cx43 mimetic peptides (146). Additionally, different melanoma cell lines or MCF-7 breast cancer cells with low or negative expression of Cx43, showed diminished susceptibility to NK cell-mediated lysis compared to the Cx43-overexpressing counterparts (147). Additional data, indicating the importance of Cx43 at the NK cell/tumor cell lytic IS, were obtained from the evaluation of the regulation of Cx43 by tumor hypoxia. Hypoxic stress, frequently occurring in the microenvironment of solid tumors, is involved in the tumor escape of immune surveillance, including a suppressed susceptibility of tumor cell lysis by CTLs and NK cells (160). While hypoxic stress increased the total Cx43 protein level in a hypoxia-induced factor 1α-driven manner in melanoma cells, the presence of Cx43 channels at the IS between hypoxic melanoma cells and NK cells was strongly diminished (147). The decline of Cx43 channels at the lytic IS was dependent on increased autophagic flux occurring during hypoxia. Indeed, the presence of Cx43 at the IS could be restored in hypoxic melanoma/NK cell cocultures by inhibiting hypoxia-induced autophagy flux by hydroxychloroquine or the hypoxia-induced autophagosome formation by 3-methyladenine or knock down of the ATG5 gene in hypoxic melanoma cells (147). Importantly, the inhibition of hypoxia-induced autophagy and thus the prevention of the subsequent autophagy-mediated degradation of Cx43 at the lytic IS was very effective in restoring the susceptibility of hypoxic melanoma cells toward NK cell-mediated lysis. These findings were corroborated using the endocytic Cx43Y286A mutant (161), which remained present at the lytic IS during hypoxic stress and restored the susceptibility of melanoma cells to lysis by NK cells in hypoxic conditions, which is inhibited by Cx43-specific inhibitory mimetic peptides (147). These reports highlight an important role for Cx43 channels at the lytic ISs among NK cells and tumor cells, and suggest that the low susceptibility of Cx43-negative tumor cells to NK cell immune surveillance is an additional mechanism that favors the survival of GJ-deficient tumor cells observed principally in primary tumors (162).
Altogether the heterotypic GJICs described so far, support a tumor suppressor role of Cxs, as its expression in tumor cells promotes a plethora of intercellular interactions between tumor cells and immune cells that limit tumor cell survival and growth via the induction of antitumor immune responses. In addition, malignant cells establish bidirectional communications with different stromal cells besides immune cells, such as cancer-associated fibroblast, endothelial cells, mesenchymal stem cells, bone marrow stromal cells (BMSC), and osteocytes. Heterotypic GJICs among cancer cells and their non-immune stromal cell counterparts have also been negatively associated with tumor progression (Figure 2, right panel).
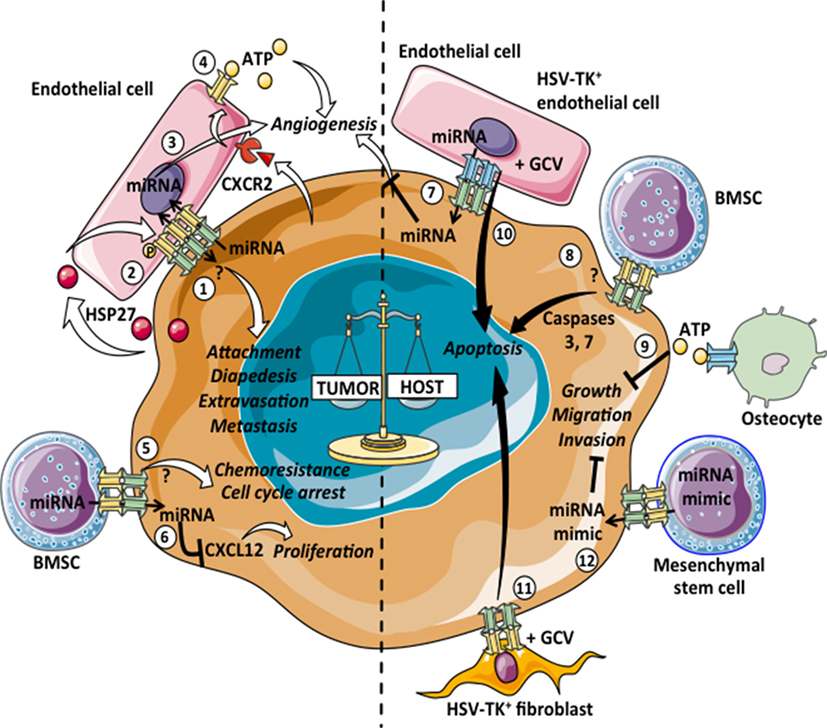
Figure 2. Pro- and antitumoral effects of tumor–stromal non-immune cell heterotypic GJICs. Gap junction (GJ)-mediated communications among tumor cells and normal non-immune cells can lead to pro-tumoral (left: 1–6) or antitumoral (right: 7–12) consequences. 1: intercellular communications mediated by Cx26- and Cx43-GJs among melanoma or breast cancer cells with endothelial cells promote cell attachment, diapedesis, extravasation and metastasis of tumor cells. 2: primary colon cancer cells release heat-shock protein 27 (HSP27), which induces the phosphorylation of Cx43 in endothelial cells and the subsequent formation of heterotypic GJs with tumor cells. This communication thus promotes transendothelial migration of primary colon cancer cells. 3: the GJ-mediated transfer of microRNAs (miRNAs) from glioblastoma cells to endothelial cells promotes angiogenesis. 4: metastatic colon cancer cells induce Cx32-HCs in endothelial cells via CXCR2. The adenosine triphosphate (ATP) released by Cx32-HCs could induce neo-angiogenesis in the metastatic foci. 5: leukemic cells GJ-coupled with bone marrow stromal cells (BMSCs) are arrested in G0 and acquire resistance to chemotherapy-induced apoptosis. 6: breast cancer cells acquired CXCL12-specific miRNAs from BMSCs via GJs, which induce cancer cell dormancy. 7: Cx43-GJ-mediated transfer of antitumoral miRNAs from human microvascular endothelial cells to colon cancer cells inhibits the angiogenesis induced by the cancer cells. 8: BMSCs overexpressing Cx43 form functional GJ with T lymphoblastic leukemia cells and increase the basal level of apoptosis due to the Cx43-dependent activation of caspases 3 and 7. 9: the ATP released through Cx43-HCs by osteocytes inhibits anchorage-independent growth, migration and invasion properties of human and mouse breast cancer cells. 10, 11: when herpes virus thymidine kinase (HSV-TK)+ endothelial cells or fibroblasts are cocultured with different tumor cell lines in the presence of ganciclovir (GCV), the extent of bystander killing correlates with the level of GJ communication between the tumor and HSV-TK+ cells. 12: mesenchymal stem cells deliver synthetic miRNA mimics to glioma and glioma stem cells via GJs, decreasing migration and self-renewal of tumor cells.
Heterotypic GJICs among Cancer Cells and Stromal Cells Associated with Antitumoral Effects
At large, GJICs between tumor cells and tumor stromal cells have been implicated in tumor inhibition. The first evidence suggesting that heterotypic GJICs between non-malignant cells and cancer cells negatively impact tumor cell growth was reported by the Loewenstein and coworkers (163). In this pioneer study, the authors showed that the growth of chemically or virally transformed malignant cells could be inhibited when those cells were chemically coupled with normal non-tumoral cells, specifically with embryo fibroblasts and rat liver cells. More recent data showed that leukemic Jurkat cells cocultured with Cx43-overexpressing BMSCs have a lower proliferation rate and higher methotrexate-induced apoptosis than Jurkat cells alone or cocultured with unmodified Cx43- and GJ-poor BMSCs (164). Similarly, BMSCs overexpressing Cx43, specifically human umbilical cord stem cells (Cx43-hUCSC), can form functional GJICs with the mouse T lymphoblastic leukemia cell line L615. The coculture of these cells increases the basal level of apoptosis in leukemic cells due to the activation of caspases 3 and 7 (165). Additionally, in a minimal residual disease mouse model, the relapse of leukemia was delayed when mice were transplanted with Cx43-hUCSC cells, suggesting a role for Cx43-mediated GJs among BMSCs and leukemic cells in the induction of tumor cell apoptosis in vivo (165). Another study indicates that Cx43-GJ-mediated transfer of the antitumoral miRNA miR-145-5p from human microvascular endothelial cells transfected with miR-145-5p mimics to primary colon cancer cells inhibits the cancer-induced tubulogenesis, suggesting that this heterotypic GJIC downregulate colon cancer cell growth by preventing the formation of new vessels (166). Additionally, a protective role of heterotypic communications mediated by Cx43-HCs has been described in the osteocyte-mediated suppression of breast cancer bone metastasis (167). The opening of Cx43-HCs in osteocytes induced by either bisphosphonate drugs or mechanical stimulation, allows for the release of ATP from osteocytes, which in turn inhibits anchorage-independent growth, migration, and invasion properties of human and mouse breast cancer cells. These inhibitory effects on cancer cells were attenuated when osteocytes were incubated with Cx43(E2), a specific Cx43-HC-blocking antibody. More interestingly, both Cx43 osteocyte-specific knockout mice and osteocyte-specific Δ130–136 transgenic mice with impaired Cx43-GJs and Cx43-HCs showed increased tumor growth and an attenuated inhibitory effect of bisphosphonate drugs, whereas R76W transgenic mice with functional Cx43-HCs but not Cx43-GJs in osteocytes did not show significant differences compared to control mice (167). These results indicate that heterotypic cell communications among normal and tumor cells, both via GJs and Cx-HCs, can mediate antitumor responses (Figure 2, right panel).
Additionally, GJICs have been associated with antitumor effects through the “bystander effect” during suicide gene therapy approaches, whereby the spread of death signals between cells occurs. Using the herpes virus thymidine kinase (HSV-TK) gene to render cancer cells sensitive to the drug ganciclovir (GCV), it was noted that HSV-TK-free neighboring tumor cells also died, and this phenomenon correlates with the level of GJs among tumor cells (168). Heterotypic GJICs have also been implicated in this kind of bystander effect. For example, when HSV-TK+ fibroblasts or HSV-TK+ endothelial cells are cocultured with different tumor cell lines, the extent of GCV-induced bystander killing correlates with the level of GJICs between tumor and HSV-TK+ fibroblasts or endothelial cells (169, 170). Also, heterotypic GJICs have a relevant role in other types of antitumor therapy, as reported by Lee and collaborators (171). They showed that mesenchymal stem cells, derived from different human tissues, efficiently deliver synthetic miRNA mimics to glioma and glioma stem cells in vivo when administered intracranially. In cocultures, it was determined that the transfer of miRNA mimics occurs via GJ- and exosome-dependent processes, affecting the expression of their target genes and decreasing the migration and self-renewal of glioma and glioma stem cells, respectively (171). Recently, functional Cx43 channels were identified in the membrane of exosomes and they can facilitate the release of exosomal content into target cells, including tumor cells, both in vitro and in vivo (172, 173). Indeed, the authors showed that when doxorubicin was incorporated into exosomes and used as a drug delivery vehicle to treat tumor-bearing mice, its antitumor effect was similar to the free drug regardless of the presence of Cx43 in exosomes; however, its cardiotoxicity was significantly lower when administrated in Cx43+ exosomes (173). This evidence strongly suggests that Cx43-GJ-mediated communications among extracellular vesicles and tumor cells could occur in vivo, and it is a very promising area to explore.
Altogether the heterotypic GJICs described so far, support a tumor suppressor role of Cxs, as its expression in tumor cells promotes a plethora of intercellular interactions between tumor cells and immune or non-immune stromal cells that limit tumor cell survival and growth. However, as we mentioned before, some specific heterotypic GJICs among cancer cells and their stroma have been positively associated with tumor progression.
Heterotypic GJICs among Cancer Cells and Stromal Cells Associated with Pro-Tumoral Effects
Several groups have reported that GJ-mediated coupling between tumor cells and endothelial cells contributes to invasion and metastasis (Figure 2, left panel). For example, the B16 melanoma cell subline BL6 establishes efficient cell coupling with endothelial cells through Cx26-Cx43 heterotypic GJs, while the Cx26 negative B16 cell subline F10 does not (174). Interestingly, BL6 cells have a major spontaneous metastatic potential compared to the F10 cells. Transfections with the wild-type Cx26 render F10 cells competent for GJ coupling with endothelial cells, which in turn increases their spontaneous metastatic potential. Conversely, transfections with a dominant negative mutant of Cx26 render BL6 cells deficient in heterotypic GJ coupling and less metastatic (174). Similarly, in human melanoma lesions, melanoma cells in the invasive and perivascular areas as well as the endothelial cells of the small vessels surrounding the melanoma cell nests expressed Cx26, while melanoma cells residing in the basal layer showed lower levels of Cx26, suggesting that heterotypic GJ-mediated cell–cell adhesion and communication contributes to melanoma metastasis in humans (54, 174, 175). A role for Cx43-mediated interactions in melanoma cell diapedesis and in melanoma-endothelial cell attachment, both processes required for metastasis, has also been suggested. Villares and coworkers showed that the expression of the protease-activated receptor-1 contributes, at least partially, to the malignant phenotype of two human metastatic melanoma cell lines via the regulation of Cx43 expression, favoring Cx43-mediated melanoma-endothelial cell attachment (176). More recently, it was shown that Cx-mediated extravasation and heterotypic GJ formation with the brain endothelium could facilitate tumor cell integration into foreign tissues creating a more hospitable niche for metastatic growth (41). Cx26 and Cx43 expression in melanoma and breast cancer cells, respectively, contributes to the in vivo cell extravasation and brain microtumor formation in association with the vasculature. Interestingly, these tumor cells establish functional GJICs with endothelial cells in vitro, and this process seems to be necessary for spheroid formation and colonization in 3D matrices (41), suggesting that Cx43 and Cx26 mediate breast cancer cell and melanoma metastasis to the brain via tumor-endothelial cell GJ-dependent mechanisms. Additionally, it has been shown that the re-expression of Cx43 in mammary carcinoma cell lines lacking endogenous Cx43 enabled the formation of heterotypic GJIC with microvascular endothelial cells and thus increased their diapedesis (177). Moreover, Cx43-GJICs between breast cancer cells and endothelial cells facilitate the metastatic homing of the tumor cells by increasing their arrest in the lung vasculature (178). Interestingly, the co-administration of avastatin (an anti-VEGF antibody used for anti-angiogenic therapy) and oleamide (a GJ chemical inhibitor) or even the administration of oleamide alone, decreases the heterotypic cell communications between MDA-MB-231 breast cancer cells with endothelial cells in vitro, increases their survival rate, and reduces pulmonary and hepatic metastatic foci in mice subdermally injected with MDA-MB-231 cells (179). However, the in vivo administration of oleamide alone does not inhibit metastasis to the lung in mice intravenously injected with MDA-MB-231 cells, suggesting that the inhibition of breast tumor-endothelial cell GJs has an anti-metastatic activity at the extravasation level (179).
Furthermore, it has been shown that primary and metastatic tumor cells can differentially modulate the expression of Cx proteins in endothelial cells. The heat-shock protein 27 released from cells derived from a primary colon tumor induces both the phosphorylation of Cx43 in endothelial cells and the formation of GJs among tumor and endothelial cells, promoting the transendothelial migration of malignant cells (180). In contrast, cells derived from a metastatic colon tumor from the same patient, induce the expression of Cx32-HCs in endothelial cells via CXCR2. The subsequent release of ATP through the Cx32-HCs by endothelial cells then modulates the crosstalk between endothelial and metastatic colon cancer cells, possibly favoring neo-angiogenesis in the metastatic foci (180). Similarly, glioblastom multiform (GBM) cells can also modulate endothelial cell function through heterotypic GJICs. The GJ-mediated transfer of the miRNA miR-5096 from GBM cells to endothelial cells promotes endothelial tubulogenesis by increasing the expression of Cx43 and the concomitant formation of heterotypic GJICs (181). Heterotypic GJs have also been described between leukemic and endothelial cells, allowing cancer cell migration and extravasation (182, 183). Finally, lung carcinoma and gastric cancer cells use heterotypic Cx43-GJICs with lymphatic endothelial cells and peritoneal mesothelial cells to support their migration through the lymphatic endothelium or the peritoneal mesothelium, respectively (184, 185). These evidences suggest that tumor cells engage GJICs with endothelial and epithelial cells to promote their migration, invasion and metastasis via blood vessels, lymphatic endothelium and peritoneal mesothelium. Taking into account the evidences described so far, we can speculate that the expression of Cxs by tumor cells growing in a vascularized microenvironment could be considered as a negative prognosis marker in cancer. However, as we previously discussed, GJICs among tumor cells and endothelial cells could also allow the cross-recognition and elimination of endothelial cells by tumor-Ag-specific CTLs (138). Therefore, we propose that Cx expression by tumor cells in a vascularized microenvironment could have a negative impact in patients with tumors poorly infiltrated by CTLs or a positive effect in those with tumors highly infiltrated by CTL; however, this hypothesis needs to be addressed.
Additionally, pro-tumoral GJICs among BMSCs and malignant cells have also been described. Reports from different groups have shown that GJICs between BMSCs and leukemic or breast cancer cells mediate the cell cycle quiescence of tumor cells. Leukemic cells coupled with BMSCs are arrested in G0, and these coupled leukemic cells are resistant to methotrexate-induced apoptosis, which can be prevented with treatments with the GJ inhibitor carbenoxolone (186). Similarly, Lim and collaborators reported that breast cancer cells acquired CXCL12-specific miRNAs from BMSCs via GJs, which is associated with the maintenance of cancer cell dormancy (187). These studies partly show how metastatic tumor cells could take advantage of GJ coupling with the bone marrow microenvironment for their survival.
The GJs among tumor cells and astrocytes are another example of pro-tumoral heterotypic GJICs occurring in cancer. Astrocytes are the most abundant glial cell population of the central nervous system (CNS), and they participate in the local innate immune response triggered by a variety of insults (188). Indeed, the majority of cancer cells that infiltrate the brain are eliminated by astrocytes (189). However, astrocytes can exert a beneficial effect on cancer cells through GJ-dependent manners (Figure 1, left panel). GJICs between GBM cells and astrocytes contribute somehow to the adhesion, migration and invasion of tumor cells to the brain parenchyma (190, 191). Recent evidence suggests that Cx43 expression in glioma cells and astrocytes influences tumor cell motility in vivo independently of its channel function (192). In a very elegant report by Chen and coworkers (110), it was demonstrated that protocadherin 7, which is expressed in human and mouse breast and lung cancer cells, promotes the assembly of Cx43-GJs between carcinoma cells and astrocytes. These Cx43-GJs allow metastatic cancer cells in the brain to transfer cGAMP to astrocytes, leading to the activation of the STING pathway and the subsequent production of IFN-α and TNF by the cGAMP-receiving astrocytes. These pro-inflammatory cytokines then cause paracrine activation of the STAT1 and NF-κB pathways in brain metastatic cells, supporting tumor growth and chemoresistance. Interestingly, in vivo inhibition of carcinoma-astrocyte GJICs, through the oral delivery of meclofenamate and tonabersat, blocks this paracrine loop, controlling metastatic outgrowth in the brain (110). Moreover, it has been shown that astrocytes can protect tumor cells from chemotherapy through additional GJ-mediated mechanisms. For example, reactive astrocytes reduce apoptosis in melanoma cells treated with different chemotherapeutic drugs by sequestering intracellular calcium via GJICs (104). In addition, astrocytes seem to upregulate the expression of various pro-survival genes in glioma cells trough a GJ-dependent manner, thus reducing the cytotoxic effects of various chemotherapeutic agents in tumor cells (193). Recently, it was shown that lung cancer cells acquire miRNAs from astrocytes in a GJ-dependent manner during in vitro cocultures (111). Several of the transferred miRNAs were implicated in cell survival pathways, and the enforced expression of these miRNAs increases the resistance of lung cancer cells to paclitaxel (111). Similarly, the GJ-mediated transfer of miR-5096 from glioma cells to astrocytes induces in vitro glioma cell invasion (194). Altogether, these data suggest that GJICs occurring between tumor cells and the main immune cells of the CNS, namely astrocytes, allow for the intercellular passage of signals that promote the colonization and survival of tumor cells in the brain.
These exciting and promising new evidences in our understanding of GJICs among tumor cells and their surrounding stromal cells, and particularly immune cells, generates the idea of potentiating the antitumor immune responses induced by current cancer immunotherapies via the modulation of GJIC. The implementation of this concept absolutely deserves further attention.
Concluding Remarks
The essential role of GJICs among tumor cells and neighboring cells of the tumor microenvironment, including immune cells, on tumor cell fate and their relationship with cancer progression is only beginning to be understood. The current literature about cell intrinsic mechanisms of Cxs and homotypic GJICs among tumor cells, in general terms, would support a tumor suppressor role of Cxs in early stages of cancer progression, while in late-stage cancer and metastasis, Cxs could act as oncogenes, promoting the progression of cancer. It is worth noting that this general conclusion depends on the Cx isoforms expressed as well as on the tumor type or subtype. Additionally, the different heterotypic GJICs occurring in the tumor microenvironment should be included in this panorama. For example, while heterotypic GJICs among tumor cells and astrocytes seem to promote tumor progression, heterotypic GJICs between tumor cells and DCs or lymphocytes are involved in tumor cell elimination. Accordingly, re-assessing Cx expression along with a deep characterization of immune cell infiltration in human tumors may, in our opinion, definitively solve the complexity of the mixed findings related to Cxs as a useful diagnostic method (Table 1). Precision-medicine diagnostic tools, such as multilabel immunofluorescence on formalin-fixed paraffin-embedded sections (195) are suitable for evaluation of Cx expression and localization in different cells from the tumor microenvironment. Although much of the data discussed in this review come from in vivo studies, several of the most exciting findings remains to be validated on accurate and specific physiological models. Smart experimental designs using current murine models, such as conditional knockout of Cx43 in T cells (76) or in DCs (127), are required to fully elucidate the physiopathological implication of GJICs on tumor immunity. Additionally, major efforts must be made to determine which intercellular signals are involved in GJ-mediated tumor immunity. In our opinion, special focus should be put on the identification and characterization of immune modulatory miRNAs that can be transferred between tumor and immune cells, and therefore, affect tumor immune attack and/or tumor immune escape.
Nowadays, novel Cx and GJ-based therapeutic approaches have emerged, particularly based on small peptides that specifically block Cx-HCs or enhance GJ plaque formation. These therapeutic approaches have shown promising results in the context of cutaneous wound healing, tissue regeneration, narcolepsy, and reperfusion injury after acute myocardial infarction (196). Efforts to move these therapeutic approaches to cancer should be made in order to promote GJ-mediated antitumor immunity. It seems clear that targeting GJIC alone is not likely to be sufficient and combination with immunotherapy treatments will be necessary.
Author Contributions
MG, MN, FH, FS-O, and AT contributed to the writing of the manuscript; AT contributed to the conception of figures and tables.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding
This work was supported by grants from the Millennium Science Initiative of the Ministry of Economy, Development and Tourism (P09/016-F); the Chilean National Fund of Scientific and Technological Development (FONDECYT 1130320, FONDECYT 11160380 and FONDECYT 3170917).
References
1. Nielsen MS, Axelsen LN, Sorgen PL, Verma V, Delmar M, Holstein-Rathlou NH. Gap junctions. Compr Physiol (2012) 2:1981–2035. doi:10.1002/cphy.c110051
2. Neijssen J, Pang B, Neefjes J. Gap junction-mediated intercellular communication in the immune system. Prog Biophys Mol Biol (2007) 94:207–18. doi:10.1016/j.pbiomolbio.2007.03.008
3. Sáez PJ, Shoji KF, Aguirre A, Sáez JC. Regulation of hemichannels and gap junction channels by cytokines in antigen-presenting cells. Mediators Inflamm (2014) 2014:742734. doi:10.1155/2014/742734
4. Zsiros V, Aradi I, Maccaferri G. Propagation of postsynaptic currents and potentials via gap junctions in GABAergic networks of the rat hippocampus. J Physiol (2007) 578:527–44. doi:10.1113/jphysiol.2006.123463
5. Veeraraghavan R, Poelzing S, Gourdie RG. Intercellular electrical communication in the heart: a new, active role for the intercalated disk. Cell Commun Adhes (2014) 21:161–7. doi:10.3109/15419061.2014.905932
6. Kotini M, Mayor R. Connexins in migration during development and cancer. Dev Biol (2015) 401:143–51. doi:10.1016/j.ydbio.2014.12.023
7. Qin J, Chang M, Wang S, Liu Z, Zhu W, Wang Y, et al. Connexin 32-mediated cell-cell communication is essential for hepatic differentiation from human embryonic stem cells. Sci Rep (2016) 6:37388. doi:10.1038/srep37388
8. Chanson M, Derouette JP, Roth I, Foglia B, Scerri I, Dudez T, et al. Gap junctional communication in tissue inflammation and repair. Biochim Biophys Acta (2005) 1711(2):197–207. doi:10.1016/j.bbamem.2004.10.005
9. Bejarano E, Yuste A, Patel B, Stout RF Jr, Spray DC, Cuervo AM. Connexins modulate autophagosome biogenesis. Nat Cell Biol (2014) 16:401–14. doi:10.1038/ncb2934
10. Krysko DV, Leybaert L, Vandenabeele P, D’Herde K. Gap junctions and the propagation of cell survival and cell death signals. Apoptosis (2005) 10:459–69. doi:10.1007/s10495-005-1875-2
11. Carette D, Gilleron J, Chevallier D, Segretain D, Pointis G. Connexin a check-point component of cell apoptosis in normal and physiopathological conditions. Biochimie (2014) 101:1–9. doi:10.1016/j.biochi.2013.11.015
12. Glass AM, Snyder EG, Taffet SM. Connexins and pannexins in the immune system and lymphatic organs. Cell Mol Life Sci (2015) 72:2899–910. doi:10.1007/s00018-015-1966-3
13. Su V, Lau AF. Connexins: mechanisms regulating protein levels and intercellular communication. FEBS Lett (2014) 588:1212–20. doi:10.1016/j.febslet.2014.01.013
14. Solan JL, Lampe PD. Specific Cx43 phosphorylation events regulate gap junction turnover in vivo. FEBS Lett (2014) 588:1423–9. doi:10.1016/j.febslet.2014.01.049
15. Saéz PJ, Shoji KF, Aguirre A, Sáez JC. Regulation of hemichannels and gap junction channels by cytokines in antigen-presenting cells. Mediators Inflamm (2014) 2014:742734. doi:10.1155/2014/742734
16. Vinken M. Regulation of connexin signaling by the epigenetic machinery. Biochim Biophys Acta (2016) 1859:262–8. doi:10.1016/j.bbagrm.2015.11.002
17. Pogoda K, Kameritsch P, Retamal MA, Vega JL. Regulation of gap junction channels and hemichannels by phosphorylation and redox changes: a revision. BMC Cell Biol (2016) 17(Suppl 1):11. doi:10.1186/s12860-016-0099-3
18. Iyyathurai J, Decuypere JP, Leybaert L, D’hondt C, Bultynck G. Connexins: substrates and regulators of autophagy. BMC Cell Biol (2016) 17(Suppl 1):20. doi:10.1186/s12860-016-0093-9
19. Falk MM, Bell CL, Kells Andrews RM, Murray SA. Molecular mechanisms regulating formation, trafficking and processing of annular gap junctions. BMC Cell Biol (2016) 17(Suppl 1):22. doi:10.1186/s12860-016-0087-7
20. Calderón JF, Retamal MA. Regulation of connexins expression levels by microRNAs, an update. Front Physiol (2016) 7:558. doi:10.3389/fphys.2016.00558
21. Puebla C, Retamal MA, Acuña R, Sáez JC. Regulation of connexin-based channels by fatty acids. Front Physiol (2017) 8:11. doi:10.3389/fphys.2017.00011
22. Wingard JC, Zhao HB. Cellular and deafness mechanisms underlying connexin mutation-induced hearing loss – a common hereditary deafness. Front Cell Neurosci (2015) 9:202. doi:10.3389/fncel.2015.00202
23. Lilly E, Sellitto C, Milstone LM, White TW. Connexin channels in congenital skin disorders. Semin Cell Dev Biol (2016) 50:4–12. doi:10.1016/j.semcdb.2015.11.018
24. Delmar M, Makita N. Cardiac connexins, mutations and arrhythmias. Curr Opin Cardiol (2012) 27:236–41. doi:10.1097/HCO.0b013e328352220e
25. Pichi F, Lembo A, Serafino M, Nucci P. Genetics of congenital cataract. Dev Ophthalmol (2016) 57:1–14. doi:10.1159/000442495
26. Aasen T, Mesnil M, Naus CC, Lampe PD, Laird DW. Gap junctions and cancer: communicating for 50 years. Nat Rev Cancer (2016) 16:775–88. doi:10.1038/nrc.2016.105
27. Loewenstein WR, Kanno Y. Intercellular communication and the control of tissue growth: lack of communication between cancer cells. Nature (1966) 209:1248–9. doi:10.1038/2091248a0
28. Naus CC, Laird DW. Implications and challenges of connexin connections to cancer. Nat Rev Cancer (2010) 10:435–41. doi:10.1038/nrc2841
29. Jiang JX, Penuela S. Connexin and pannexin channels in cancer. BMC Cell Biol (2016) 17(Suppl 1):12. doi:10.1186/s12860-016-0094-8
30. Tittarelli A, Guerrero I, Tempio F, Gleisner MA, Avalos I, Sabanegh S, et al. Overexpression of connexin 43 reduces melanoma proliferative and metastatic capacity. Br J Cancer (2015) 113:259–67. doi:10.1038/bjc.2015.162
31. Xu N, Chen HJ, Chen SH, Xue XY, Chen H, Zheng QS, et al. Reduced connexin 43 expression is associated with tumor malignant behaviors and biochemical recurrence-free survival of prostate cancer. Oncotarget (2016) 7:67476–84. doi:10.18632/oncotarget.11231
32. Bijnsdorp IV, Rozendaal L, van Moorselaar RJ, Geldof AA. A predictive role for noncancerous prostate cells: low connexin-26 expression in radical prostatectomy tissues predicts metastasis. Br J Cancer (2012) 107:1963–8. doi:10.1038/bjc.2012.500
33. Benko G, Spajić B, Demirović A, Stimac G, Kru Sbreve Lin B, Tomas D. Prognostic value of connexin43 expression in patients with clinically localized prostate cancer. Prostate Cancer Prostatic Dis (2011) 14:90–5. doi:10.1038/pcan.2010.51
34. Tanaka T, Kimura M, Ishiguro H, Mizoguchi K, Takeyama H. Connexin 43 expression is associated with poor survival in patients with esophageal squamous cell carcinoma. Mol Clin Oncol (2016) 4:989–93. doi:10.3892/mco.2016.828
35. Inose T, Kato H, Kimura H, Faried A, Tanaka N, Sakai M, et al. Correlation between connexin 26 expression and poor prognosis of esophageal squamous cell carcinoma. Ann Surg Oncol (2009) 16:1704–10. doi:10.1245/s10434-009-0443-3
36. Balla P, Maros ME, Barna G, Antal I, Papp G, Sapi Z, et al. Prognostic impact of reduced connexin43 expression and gap junction coupling of neoplastic stromal cells in giant cell tumor of bone. PLoS One (2015) 10:e0125316. doi:10.1371/journal.pone.0125316
37. Poyet C, Buser L, Roudnicky F, Detmar M, Hermanns T, Mannhard D, et al. Connexin 43 expression predicts poor progression-free survival in patients with non-muscle invasive urothelial bladder cancer. J Clin Pathol (2015) 68:819–24. doi:10.1136/jclinpath-2015-202898
38. Teleki I, Szasz AM, Maros ME, Gyorffy B, Kulka J, Meggyeshazi N, et al. Correlations of differentially expressed gap junction connexins Cx26, Cx30, Cx32, Cx43 and Cx46 with breast cancer progression and prognosis. PLoS One (2014) 9:e112541. doi:10.1371/journal.pone.0112541
39. Teleki I, Krenacs T, Szasz MA, Kulka J, Wichmann B, Leo C, et al. The potential prognostic value of connexin 26 and 46 expression in neoadjuvant-treated breast cancer. BMC Cancer (2013) 13:50. doi:10.1186/1471-2407-13-50
40. Naoi Y, Miyoshi Y, Taguchi T, Kim SJ, Arai T, Tamaki Y, et al. Connexin26 expression is associated with lymphatic vessel invasion and poor prognosis in human breast cancer. Breast Cancer Res Treat (2007) 106:11–7. doi:10.1007/s10549-006-9465-8
41. Stoletov K, Strnadel J, Zardouzian E, Momiyama M, Park FD, Kelber JA, et al. Role of connexins in metastatic breast cancer and melanoma brain colonization. J Cell Sci (2013) 126:904–13. doi:10.1242/jcs.112748
42. Brockmeyer P, Jung K, Perske C, Schliephake H, Hemmerlein B. Membrane connexin 43 acts as an independent prognostic marker in oral squamous cell carcinoma. Int J Oncol (2014) 45:273–81. doi:10.3892/ijo.2014.2394
43. Tang B, Peng ZH, Yu PW, Yu G, Qian F. Expression and significance of Cx43 and E-cadherin in gastric cancer and metastatic lymph nodes. Med Oncol (2011) 28:502–8. doi:10.1007/s12032-010-9492-5
44. Liu X, Furuya T, Li D, Xu J, Cao X, Li Q, et al. Connexin 26 expression correlates with less aggressive phenotype of intestinal type-gastric carcinomas. Int J Mol Med (2010) 25:709–16. doi:10.3892/ijmm_00000395
45. Du G, Yang Y, Zhang Y, Sun T, Liu W, Wang Y, et al. Thrombocytosis and immunohistochemical expression of connexin 43 at diagnosis predict survival in advanced non-small-cell lung cancer treated with cisplatin-based chemotherapy. Cancer Chemother Pharmacol (2013) 71:893–904. doi:10.1007/s00280-013-2080-6
46. Chen JT, Cheng YW, Chou MC, Sen-Lin T, Lai WW, Ho WL, et al. The correlation between aberrant connexin 43 mRNA expression induced by promoter methylation and nodal micrometastasis in non-small cell lung cancer. Clin Cancer Res (2003) 9:4200–4.
47. Ito A, Koma Y, Uchino K, Okada T, Ohbayashi C, Tsubota N, et al. Increased expression of connexin 26 in the invasive component of lung squamous cell carcinoma: significant correlation with poor prognosis. Cancer Lett (2006) 234:239–48. doi:10.1016/j.canlet.2005.03.049
48. Hitomi M, Deleyrolle LP, Mulkearns-Hubert EE, Jarrar A, Li M, Sinyuk M, et al. Differential connexin function enhances self-renewal in glioblastoma. Cell Rep (2015) 11:1031–42. doi:10.1016/j.celrep.2015.04.021
49. Dános K, Brauswetter D, Birtalan E, Pató A, Bencsik G, Krenács T, et al. The potential prognostic value of connexin 43 expression in head and neck squamous cell carcinomas. Appl Immunohistochem Mol Morphol (2016) 24:476–81. doi:10.1097/PAI.0000000000000212
50. Wang ZS, Wu LQ, Yi X, Geng C, Li YJ, Yao RY. Connexin-43 can delay early recurrence and metastasis in patients with hepatitis B-related hepatocellular carcinoma and low serum alpha-fetoprotein after radical hepatectomy. BMC Cancer (2013) 13:306. doi:10.1186/1471-2407-13-306
51. Sirnes S, Bruun J, Kolberg M, Kjenseth A, Lind GE, Svindland A, et al. Connexin43 acts as a colorectal cancer tumor suppressor and predicts disease outcome. Int J Cancer (2012) 131:570–81. doi:10.1002/ijc.26392
52. Nomura S, Maeda K, Noda E, Inoue T, Fukunaga S, Nagahara H, et al. Clinical significance of the expression of connexin26 in colorectal cancer. J Exp Clin Cancer Res (2010) 29:79. doi:10.1186/1756-9966-29-79
53. Ezumi K, Yamamoto H, Murata K, Higashiyama M, Damdinsuren B, Nakamura Y, et al. Aberrant expression of connexin 26 is associated with lung metastasis of colorectal cancer. Clin Cancer Res (2008) 14:677–84. doi:10.1158/1078-0432.CCR-07-1184
54. Haass NK, Ripperger D, Wladykowski E, Dawson P, Gimotty PA, Blome C, et al. Melanoma progression exhibits a significant impact on connexin expression patterns in the epidermal tumor microenvironment. Histochem Cell Biol (2010) 133:113–24. doi:10.1007/s00418-009-0654-5
55. Liang QL, Wang BR, Chen GQ, Li GH, Xu YY. Clinical significance of vascular endothelial growth factor and connexin43 for predicting pancreatic cancer clinicopathologic parameters. Med Oncol (2010) 27:1164–70. doi:10.1007/s12032-009-9354-1
56. Zhu T, Gao YF, Chen YX, Wang ZB, Yin JY, Mao XY, et al. Genome-scale analysis identifies GJB2 and ERO1LB as prognosis markers in patients with pancreatic cancer. Oncotarget (2017) 8(13):21281–9. doi:10.18632/oncotarget.15068
57. Zhang YW, Kaneda M, Morita I. The gap junction-independent tumor-suppressing effect of connexin 43. J Biol Chem (2003) 278:44852–6. doi:10.1074/jbc.M305072200
58. Schalper KA, Carvajal-Hausdorf D, Oyarzo MP. Possible role of hemichannels in cancer. Front Physiol (2014) 5:237. doi:10.3389/fphys.2014.00237
59. Oviedo-Orta E, Howard Evans W. Gap junctions and connexin-mediated communication in the immune system. Biochim Biophys Acta (2004) 1662:102–12. doi:10.1016/j.bbamem.2003.10.021
60. Valdebenito S, Barreto A, Eugenin EA. The role of connexin and pannexin containing channels in the innate and acquired immune response. Biochim Biophys Acta (2017). doi:10.1016/j.bbamem.2017.05.015
61. Montecino-Rodriguez E, Leathers H, Dorshkind K. Expression of connexin 43 (Cx43) is critical for normal hematopoiesis. Blood (2000) 96:917–24.
62. Presley CA, Lee AW, Kastl B, Igbinosa I, Yamada Y, Fishman GI, et al. Bone marrow connexin-43 expression is critical for hematopoietic regeneration after chemotherapy. Cell Commun Adhes (2005) 12:307–17. doi:10.1080/15419060500514200
63. Taniguchi Ishikawa E, Gonzalez-Nieto D, Ghiaur G, Dunn SK, Ficker AM, Murali B, et al. Connexin-43 prevents hematopoietic stem cell senescence through transfer of reactive oxygen species to bone marrow stromal cells. Proc Natl Acad Sci U S A (2012) 109:9071–6. doi:10.1073/pnas.1120358109
64. Hirabayashi Y, Yoon BI, Tsuboi I, Huo Y, Kodama Y, Kanno J, et al. Protective role of connexin 32 in steady-state hematopoiesis, regeneration state, and leukemogenesis. Exp Biol Med (Maywood) (2007) 232:700–12.
65. Cancelas JA, Koevoet WL, de Koning AE, Mayen AE, Rombouts EJ, Ploemacher RE. Connexin-43 gap junctions are involved in multiconnexin-expressing stromal support of hemopoietic progenitors and stem cells. Blood (2000) 96:498–505.
66. Bodi E, Hurtado SP, Carvalho MA, Borojevic R, Carvalho AC. Gap junctions in hematopoietic stroma control proliferation and differentiation of blood cell precursors. An Acad Bras Cienc (2004) 76:743–56. doi:10.1590/S0001-37652004000400009
67. Flenniken AM, Osborne LR, Anderson N, Ciliberti N, Fleming C, Gittens JE, et al. A Gja1 missense mutation in a mouse model of oculodentodigital dysplasia. Development (2005) 132:4375–86. doi:10.1242/dev.02011
68. Paredes-Gamero EJ, Leon CM, Borojevic R, Oshiro ME, Ferreira AT. Changes in intracellular Ca2+ levels induced by cytokines and P2 agonists differentially modulate proliferation or commitment with macrophage differentiation in murine hematopoietic cells. J Biol Chem (2008) 283:31909–19. doi:10.1074/jbc.M801990200
69. Gonzalez-Nieto D, Li L, Kohler A, Ghiaur G, Ishikawa E, Sengupta A, et al. Connexin-43 in the osteogenic BM niche regulates its cellular composition and the bidirectional traffic of hematopoietic stem cells and progenitors. Blood (2012) 119:5144–54. doi:10.1182/blood-2011-07-368506
70. Schajnovitz A, Itkin T, D’Uva G, Kalinkovich A, Golan K, Ludin A, et al. CXCL12 secretion by bone marrow stromal cells is dependent on cell contact and mediated by connexin-43 and connexin-45 gap junctions. Nat Immunol (2011) 12:391–8. doi:10.1038/ni.2017
71. Angelillo-Scherrer A, Fontana P, Burnier L, Roth I, Sugamele R, Brisset A, et al. Connexin 37 limits thrombus propensity by downregulating platelet reactivity. Circulation (2011) 124:930–9. doi:10.1161/CIRCULATIONAHA.110.015479
72. Vaiyapuri S, Jones CI, Sasikumar P, Moraes LA, Munger SJ, Wright JR, et al. Gap junctions and connexin hemichannels underpin hemostasis and thrombosis. Circulation (2012) 125:2479–91. doi:10.1161/CIRCULATIONAHA.112.101246
73. Vaiyapuri S, Moraes LA, Sage T, Ali MS, Lewis KR, Mahaut-Smith MP, et al. Connexin40 regulates platelet function. Nat Commun (2013) 4:2564. doi:10.1038/ncomms3564
74. Bopp T, Becker C, Klein M, Klein-Hessling S, Palmetshofer A, Serfling E, et al. Cyclic adenosine monophosphate is a key component of regulatory T cell-mediated suppression. J Exp Med (2007) 204:1303–10. doi:10.1084/jem.20062129
75. Ring S, Karakhanova S, Johnson T, Enk AH, Mahnke K. Gap junctions between regulatory T cells and dendritic cells prevent sensitization of CD8(+) T cells. J Allergy Clin Immunol (2010) 125:237–46.e1–7. doi:10.1016/j.jaci.2009.10.025
76. Kuczma M, Lee JR, Kraj P. Connexin 43 signaling enhances the generation of Foxp3+ regulatory T cells. J Immunol (2011) 187:248–57. doi:10.4049/jimmunol.1003785
77. Weber M, Lupp C, Stein P, Kreft A, Bopp T, Wehler TC, et al. Mechanisms of cyclic nucleotide phosphodiesterases in modulating T cell responses in murine graft-versus-host disease. PLoS One (2013) 8:e58110. doi:10.1371/journal.pone.0058110
78. Kuczma M, Wang CY, Ignatowicz L, Gourdie R, Kraj P. Altered connexin 43 expression underlies age-dependent decrease of regulatory T cell suppressor function in nonobese diabetic mice. J Immunol (2015) 194:5261–71. doi:10.4049/jimmunol.1400887
79. Zahler S, Hoffmann A, Gloe T, Pohl U. Gap-junctional coupling between neutrophils and endothelial cells: a novel modulator of transendothelial migration. J Leukoc Biol (2003) 73:118–26. doi:10.1189/jlb.0402184
80. Frossard JL, Rubbia-Brandt L, Wallig MA, Benathan M, Ott T, Morel P, et al. Severe acute pancreatitis and reduced acinar cell apoptosis in the exocrine pancreas of mice deficient for the Cx32 gene. Gastroenterology (2003) 124:481–93. doi:10.1053/gast.2003.50052
81. Eugenín EA, Brañes MC, Berman JW, Sáez JC. TNF-alpha plus IFN-gamma induce connexin43 expression and formation of gap junctions between human monocytes/macrophages that enhance physiological responses. J Immunol (2003) 170:1320–8. doi:10.4049/jimmunol.170.3.1320
82. Kwak BR, Veillard N, Pelli G, Mulhaupt F, James RW, Chanson M, et al. Reduced connexin43 expression inhibits atherosclerotic lesion formation in low-density lipoprotein receptor-deficient mice. Circulation (2003) 107:1033–9. doi:10.1161/01.CIR.0000051364.70064.D1
83. Wong CW, Burger F, Pelli G, Mach F, Kwak BR. Dual benefit of reduced Cx43 on atherosclerosis in LDL receptor-deficient mice. Cell Commun Adhes (2003) 10:395–400. doi:10.1080/cac.10.4-6.395.400
84. Qiu C, Coutinho P, Frank S, Franke S, Law LY, Martin P, et al. Targeting connexin43 expression accelerates the rate of wound repair. Curr Biol (2003) 13:1697–703. doi:10.1016/j.cub.2003.09.007
85. Eltzschig HK, Eckle T, Mager A, Küper N, Karcher C, Weissmüller T, et al. ATP release from activated neutrophils occurs via connexin 43 and modulates adenosine-dependent endothelial cell function. Circ Res (2006) 99:1100–8. doi:10.1161/01.RES.0000250174.31269.70
86. Wong CW, Christen T, Roth I, Chadjichristos CE, Derouette JP, Foglia BF, et al. Connexin37 protects against atherosclerosis by regulating monocyte adhesion. Nat Med (2006) 12:950–4. doi:10.1038/nm1441
87. Véliz LP, González FG, Duling BR, Sáez JC, Boric MP. Functional role of gap junctions in cytokine-induced leukocyte adhesion to endothelium in vivo. Am J Physiol Heart Circ Physiol (2008) 295:H1056–66. doi:10.1152/ajpheart.00266.2008
88. Pistorio AL, Ehrlich HP. Modulatory effects of connexin-43 expression on gap junction intercellular communications with mast cells and fibroblasts. J Cell Biochem (2011) 112:1441–9. doi:10.1002/jcb.23061
89. Vliagoftis H, Ebeling C, Ilarraza R, Mahmudi-Azer S, Abel M, Adamko D, et al. Connexin 43 expression on peripheral blood eosinophils: role of gap junctions in transendothelial migration. Biomed Res Int (2014) 2014:803257. doi:10.1155/2014/803257
90. Machtaler S, Choi K, Dang-Lawson M, Falk L, Pournia F, Naus CC, et al. The role of the gap junction protein connexin43 in B lymphocyte motility and migration. FEBS Lett (2014) 588:1249–58. doi:10.1016/j.febslet.2014.01.027
91. Anand RJ, Dai S, Gribar SC, Richardson W, Kohler JW, Hoffman RA, et al. A role for connexin43 in macrophage phagocytosis and host survival after bacterial peritoneal infection. J Immunol (2008) 181:8534–43. doi:10.4049/jimmunol.181.12.8534
92. Patel SJ, King KR, Casali M, Yarmush ML. DNA-triggered innate immune responses are propagated by gap junction communication. Proc Natl Acad Sci U S A (2009) 106:12867–72. doi:10.1073/pnas.0809292106
93. Kasper CA, Sorg I, Schmutz C, Tschon T, Wischnewski H, Kim ML, et al. Cell-cell propagation of NF-κB transcription factor and MAP kinase activation amplifies innate immunity against bacterial infection. Immunity (2010) 33:804–16. doi:10.1016/j.immuni.2010.10.015
94. Robertson J, Lang S, Lambert PA, Martin PE. Peptidoglycan derived from Staphylococcus epidermidis induces Connexin43 hemichannel activity with consequences on the innate immune response in endothelial cells. Biochem J (2010) 432:133–43. doi:10.1042/BJ20091753
95. Moreno-Fernandez ME, Rueda CM, Rusie LK, Chougnet CA. Regulatory T cells control HIV replication in activated T cells through a cAMP-dependent mechanism. Blood (2011) 117:5372–80. doi:10.1182/blood-2010-12-323162
96. Ablasser A, Schmid-Burgk JL, Hemmerling I, Horvath GL, Schmidt T, Latz E, et al. Cell intrinsic immunity spreads to bystander cells via the intercellular transfer of cGAMP. Nature (2013) 503:530–4. doi:10.1038/nature12640
97. Zhang Y, Yeruva L, Marinov A, Prantner D, Wyrick PB, Lupashin V, et al. The DNA sensor, cyclic GMP-AMP synthase, is essential for induction of IFN-β during Chlamydia trachomatis infection. J Immunol (2014) 193:2394–404. doi:10.4049/jimmunol.1302718
98. Qin J, Zhang G, Zhang X, Tan B, Lv Z, Liu M, et al. TLR-activated gap junction channels protect mice against bacterial infection through extracellular UDP release. J Immunol (2016) 196:1790–8. doi:10.4049/jimmunol.1501629
99. Nakase T, Söhl G, Theis M, Willecke K, Naus CC. Increased apoptosis and inflammation after focal brain ischemia in mice lacking connexin43 in astrocytes. Am J Pathol (2004) 164:2067–75. doi:10.1016/S0002-9440(10)63765-0
100. Takeuchi H, Jin S, Wang J, Zhang G, Kawanokuchi J, Kuno R, et al. Tumor necrosis factor-alpha induces neurotoxicity via glutamate release from hemichannels of activated microglia in an autocrine manner. J Biol Chem (2006) 281:21362–8. doi:10.1074/jbc.M600504200
101. Takeuchi H, Jin S, Suzuki H, Doi Y, Liang J, Kawanokuchi J, et al. Blockade of microglial glutamate release protects against ischemic brain injury. Exp Neurol (2008) 214:144–6. doi:10.1016/j.expneurol.2008.08.001
102. Shijie J, Takeuchi H, Yawata I, Harada Y, Sonobe Y, Doi Y, et al. Blockade of glutamate release from microglia attenuates experimental autoimmune encephalomyelitis in mice. Tohoku J Exp Med (2009) 217:87–92. doi:10.1620/tjem.217.87
103. Retamal MA, Froger N, Palacios-Prado N, Ezan P, Sáez PJ, Sáez JC, et al. Cx43 hemichannels and gap junction channels in astrocytes are regulated oppositely by proinflammatory cytokines released from activated microglia. J Neurosci (2007) 27:13781–92. doi:10.1523/JNEUROSCI.2042-07.2007
104. Lin Q, Balasubramanian K, Fan D, Kim SJ, Guo L, Wang H, et al. Reactive astrocytes protect melanoma cells from chemotherapy by sequestering intracellular calcium through gap junction communication channels. Neoplasia (2010) 12:748–54. doi:10.1593/neo.10602
105. Froger N, Orellana JA, Calvo CF, Amigou E, Kozoriz MG, Naus CC, et al. Inhibition of cytokine-induced connexin43 hemichannel activity in astrocytes is neuroprotective. Mol Cell Neurosci (2010) 45:37–46. doi:10.1016/j.mcn.2010.05.007
106. Orellana JA, Hernández DE, Ezan P, Velarde V, Bennett MV, Giaume C, et al. Hypoxia in high glucose followed by reoxygenation in normal glucose reduces the viability of cortical astrocytes through increased permeability of connexin 43 hemichannels. Glia (2010) 58:329–43. doi:10.1002/glia.20926
107. Orellana JA, Froger N, Ezan P, Jiang JX, Bennett MV, Naus CC, et al. ATP and glutamate released via astroglial connexin 43 hemichannels mediate neuronal death through activation of pannexin 1 hemichannels. J Neurochem (2011) 118:826–40. doi:10.1111/j.1471-4159.2011.07210.x
108. Wasseff SK, Scherer SS. Activated immune response in an inherited leukodystrophy disease caused by the loss of oligodendrocyte gap junctions. Neurobiol Dis (2015) 82:86–98. doi:10.1016/j.nbd.2015.05.018
109. Boulay AC, Mazeraud A, Cisternino S, Saubaméa B, Mailly P, Jourdren L, et al. Immune quiescence of the brain is set by astroglial connexin 43. J Neurosci (2015) 35:4427–39. doi:10.1523/JNEUROSCI.2575-14.2015
110. Chen Q, Boire A, Jin X, Valiente M, Er EE, Lopez-Soto A, et al. Carcinoma-astrocyte gap junctions promote brain metastasis by cGAMP transfer. Nature (2016) 533:493–8. doi:10.1038/nature18268
111. Menachem A, Makovski V, Bodner O, Pasmanik-Chor M, Stein R, Shomron N, et al. Intercellular transfer of small RNAs from astrocytes to lung tumor cells induces resistance to chemotherapy. Oncotarget (2016) 7:12489–504. doi:10.18632/oncotarget.7273
112. Sanderson MJ, Chow I, Dirksen ER. Intercellular communication between ciliated cells in culture. Am J Physiol (1988) 254:C63–74.
113. Boitano S, Dirksen ER, Sanderson MJ. Intercellular propagation of calcium waves mediated by inositol trisphosphate. Science (1992) 258:292–5. doi:10.1126/science.1411526
114. Homolya L, Steinberg TH, Boucher RC. Cell to cell communication in response to mechanical stress via bilateral release of ATP and UTP in polarized epithelia. J Cell Biol (2000) 150:1349–60. doi:10.1083/jcb.150.6.1349
115. Parthasarathi K, Ichimura H, Monma E, Lindert J, Quadri S, Issekutz A, et al. Connexin 43 mediates spread of Ca2+-dependent proinflammatory responses in lung capillaries. J Clin Invest (2006) 116:2193–200. doi:10.1172/JCI26605C1
116. Tran Van Nhieu G, Clair C, Bruzzone R, Mesnil M, Sansonetti P, Combettes L. Connexin-dependent inter-cellular communication increases invasion and dissemination of Shigella in epithelial cells. Nat Cell Biol (2003) 5:720–6. doi:10.1038/ncb1021
117. Romero S, Grompone G, Carayol N, Mounier J, Guadagnini S, Prevost MC, et al. ATP-mediated Erk1/2 activation stimulates bacterial capture by filopodia, which precedes Shigella invasion of epithelial cells. Cell Host Microbe (2011) 9:508–19. doi:10.1016/j.chom.2011.05.005
118. Simpson C, Kelsell DP, Marchès O. Connexin 26 facilitates gastrointestinal bacterial infection in vitro. Cell Tissue Res (2013) 351:107–16. doi:10.1007/s00441-012-1502-9
119. Martin FJ, Prince AS. TLR2 regulates gap junction intercellular communication in airway cells. J Immunol (2008) 180:4986–93. doi:10.4049/jimmunol.180.7.4986
120. Sarieddine MZ, Scheckenbach KE, Foglia B, Maass K, Garcia I, Kwak BR, et al. Connexin43 modulates neutrophil recruitment to the lung. J Cell Mol Med (2009) 13:4560–70. doi:10.1111/j.1582-4934.2008.00654.x
121. Guttman JA, Lin AE, Li Y, Bechberger J, Naus CC, Vogl AW, et al. Gap junction hemichannels contribute to the generation of diarrhoea during infectious enteric disease. Gut (2010) 59:218–26. doi:10.1136/gut.2008.170464
122. Ey B, Eyking A, Gerken G, Podolsky DK, Cario E. TLR2 mediates gap junctional intercellular communication through connexin-43 in intestinal epithelial barrier injury. J Biol Chem (2009) 284:22332–43. doi:10.1074/jbc.M901619200
123. Chadjichristos CE, Scheckenbach KE, van Veen TA, Richani Sarieddine MZ, de Wit C, Yang Z, et al. Endothelial-specific deletion of connexin40 promotes atherosclerosis by increasing CD73-dependent leukocyte adhesion. Circulation (2010) 121:123–31. doi:10.1161/CIRCULATIONAHA.109.867176
124. Scheckenbach KE, Losa D, Dudez T, Bacchetta M, O’Grady S, Crespin S, et al. Prostaglandin E2 regulation of cystic fibrosis transmembrane conductance regulator activity and airway surface liquid volume requires gap junctional communication. Am J Respir Cell Mol Biol (2011) 44:74–82. doi:10.1165/rcmb.2009-0361OC
125. Puhar A, Tronchère H, Payrastre B, Nhieu GT, Sansonetti PJ. A Shigella effector dampens inflammation by regulating epithelial release of danger signal ATP through production of the lipid mediator PtdIns5P. Immunity (2013) 39:1121–31. doi:10.1016/j.immuni.2013.11.013
126. Losa D, Köhler T, Bellec J, Dudez T, Crespin S, Bacchetta M, et al. Pseudomonas aeruginosa-induced apoptosis in airway epithelial cells is mediated by gap junctional communication in a JNK-dependent manner. J Immunol (2014) 192:4804–12. doi:10.4049/jimmunol.1301294
127. Mazzini E, Massimiliano L, Penna G, Rescigno M. Oral tolerance can be established via gap junction transfer of fed antigens from CX3CR1+ macrophages to CD103+ dendritic cells. Immunity (2014) 40:248–61. doi:10.1016/j.immuni.2013.12.012
128. Westphalen K, Gusarova GA, Islam MN, Subramanian M, Cohen TS, Prince AS, et al. Sessile alveolar macrophages communicate with alveolar epithelium to modulate immunity. Nature (2014) 506:503–6. doi:10.1038/nature12902
129. Birchenough GM, Nyström EE, Johansson ME, Hansson GC. A sentinel goblet cell guards the colonic crypt by triggering Nlrp6-dependent Muc2 secretion. Science (2016) 352:1535–42. doi:10.1126/science.aaf7419
130. Krenacs T, van Dartel M, Lindhout E, Rosendaal M. Direct cell/cell communication in the lymphoid germinal center: connexin43 gap junctions functionally couple follicular dendritic cells to each other and to B lymphocytes. Eur J Immunol (1997) 27:1489–97. doi:10.1002/eji.1830270627
131. Rajnai H, Teleki I, Kiszner G, Meggyesházi N, Balla P, Vancsik T, et al. Connexin 43 communication channels in follicular dendritic cell development and in follicular lymphomas. J Immunol Res (2015) 2015:528098. doi:10.1155/2015/528098
132. Oviedo-Orta E, Hoy T, Evans WH. Intercellular communication in the immune system: differential expression of connexin40 and 43, and perturbation of gap junction channel functions in peripheral blood and tonsil human lymphocyte subpopulations. Immunology (2000) 99:578–90. doi:10.1046/j.1365-2567.2000.00991.x
133. Oviedo-Orta E, Gasque P, Evans WH. Immunoglobulin and cytokine expression in mixed lymphocyte cultures is reduced by disruption of gap junction intercellular communication. FASEB J (2001) 15:768–74. doi:10.1096/fj.00-0288com
134. Neijssen J, Herberts C, Drijfhout JW, Reits E, Janssen L, Neefjes J. Cross-presentation by intercellular peptide transfer through gap junctions. Nature (2005) 434:83–8. doi:10.1038/nature03290
135. Matsue H, Yao J, Matsue K, Nagasaka A, Sugiyama H, Aoki R, et al. Gap junction-mediated intercellular communication between dendritic cells (DCs) is required for effective activation of DCs. J Immunol (2006) 176(1):181–90. doi:10.4049/jimmunol.176.4.2670-d
136. Mendoza-Naranjo A, Saéz PJ, Johansson CC, Ramírez M, Mandakovic D, Pereda C, et al. Functional gap junctions facilitate melanoma antigen transfer and cross-presentation between human dendritic cells. J Immunol (2007) 178:6949–57. doi:10.4049/jimmunol.178.11.6949
137. Pang B, Neijssen J, Qiao X, Janssen L, Janssen H, Lippuner C, et al. Direct antigen presentation and gap junction mediated cross-presentation during apoptosis. J Immunol (2009) 183:1083–90. doi:10.4049/jimmunol.0900861
138. Benlalam H, Jalil A, Hasmim M, Pang B, Tamouza R, Mitterrand M, et al. Gap junction communication between autologous endothelial and tumor cells induce cross-recognition and elimination by specific CTL. J Immunol (2009) 182:2654–64. doi:10.4049/jimmunol.0800815
139. Saccheri F, Pozzi C, Avogadri F, Barozzi S, Faretta M, Fusi P, et al. Bacteria-induced gap junctions in tumors favor antigen cross-presentation and antitumor immunity. Sci Transl Med (2010) 2:44ra57. doi:10.1126/scitranslmed.3000739
140. Elgueta R, Tobar JA, Shoji KF, De Calisto J, Kalergis AM, Bono MR, et al. Gap junctions at the dendritic cell-T cell interface are key elements for antigen-dependent T cell activation. J Immunol (2009) 183:277–84. doi:10.4049/jimmunol.0801854
141. Mendoza-Naranjo A, Bouma G, Pereda C, Ramírez M, Webb KF, Tittarelli A, et al. Functional gap junctions accumulate at the immunological synapse and contribute to T cell activation. J Immunol (2011) 187:3121–32. doi:10.4049/jimmunol.1100378
142. Yu F, Yan H, Nie W, Zhu J. Connexin43 knockdown in bone marrow-derived dendritic cells by small interfering RNA leads to a diminished T-cell stimulation. Mol Med Rep (2016) 13:895–900. doi:10.3892/mmr.2015.4593
143. Oviedo-Orta E, Perreau M, Evans WH, Potolicchio I. Control of the proliferation of activated CD4+ T cells by connexins. J Leukoc Biol (2010) 88:79–86. doi:10.1189/jlb.0909613
144. Machtaler S, Dang-Lawson M, Choi K, Jang C, Naus CC, Matsuuchi L. The gap junction protein Cx43 regulates B-lymphocyte spreading and adhesion. J Cell Sci (2011) 124:2611–21. doi:10.1242/jcs.089532
145. Aucher A, Rudnicka D, Davis DM. MicroRNAs transfer from human macrophages to hepato-carcinoma cells and inhibit proliferation. J Immunol (2013) 191:6250–60. doi:10.4049/jimmunol.1301728
146. Tittarelli A, Mendoza-Naranjo A, Farías M, Guerrero I, Ihara F, Wennerberg E, et al. Gap junction intercellular communications regulate NK cell activation and modulate NK cytotoxic capacity. J Immunol (2014) 192:1313–9. doi:10.4049/jimmunol.1301297
147. Tittarelli A, Janji B, Van Moer K, Noman MZ, Chouaib S. The selective degradation of synaptic connexin 43 protein by hypoxia-induced autophagy impairs natural killer cell-mediated tumor cell killing. J Biol Chem (2015) 290:23670–9. doi:10.1074/jbc.M115.651547
148. Escobar A, López M, Serrano A, Ramirez M, Pérez C, Aguirre A, et al. Dendritic cell immunizations alone or combined with low doses of interleukin-2 induce specific immune responses in melanoma patients. Clin Exp Immunol (2005) 142:555–68. doi:10.1111/j.1365-2249.2005.02948.x
149. López MN, Pereda C, Segal G, Muñoz L, Aguilera R, González FE, et al. Prolonged survival of dendritic cell-vaccinated melanoma patients correlates with tumor-specific delayed type IV hypersensitivity response and reduction of tumor growth factor beta-expressing T cells. J Clin Oncol (2009) 27:945–52. doi:10.1200/JCO.2008.18.0794
150. Aguilera R, Saffie C, Tittarelli A, González FE, Ramírez M, Reyes D, et al. Heat-shock induction of tumor-derived danger signals mediates rapid monocyte differentiation into clinically effective dendritic cells. Clin Cancer Res (2011) 17:2474–83. doi:10.1158/1078-0432.CCR-10-2384
151. Tittarelli A, González FE, Pereda C, Mora G, Muñoz L, Saffie C, et al. Toll-like receptor 4 gene polymorphism influences dendritic cell in vitro function and clinical outcomes in vaccinated melanoma patients. Cancer Immunol Immunother (2012) 61:2067–77. doi:10.1007/s00262-012-1268-7
152. Durán-Aniotz C, Segal G, Salazar L, Pereda C, Falcón C, Tempio F, et al. The immunological response and post-treatment survival of DC-vaccinated melanoma patients are associated with increased Th1/Th17 and reduced Th3 cytokine responses. Cancer Immunol Immunother (2013) 62:761–72. doi:10.1007/s00262-012-1377-3
153. González FE, Gleisner A, Falcón-Beas F, Osorio F, López MN, Salazar-Onfray F. Tumor cell lysates as immunogenic sources for cancer vaccine design. Hum Vaccin Immunother (2014) 10:3261–9. doi:10.4161/21645515.2014.982996
154. Kleindienst P, Brocker T. Endogenous dendritic cells are required for amplification of T cell responses induced by dendritic cell vaccines in vivo. J Immunol (2003) 170:2817–23. doi:10.4049/jimmunol.170.6.2817
155. Yewdall AW, Drutman SB, Jinwala F, Bahjat KS, Bhardwaj N. CD8+ T cell priming by dendritic cell vaccines requires antigen transfer to endogenous antigen presenting cells. PLoS One (2010) 5:e11144. doi:10.1371/journal.pone.0011144
156. Green DR, Ferguson T, Zitvogel L, Kroemer G. Immunogenic and tolerogenic cell death. Nat Rev Immunol (2009) 9:353–63. doi:10.1038/nri2545
157. Benlalam H, Carré T, Jalil A, Noman Z, Caillou B, Vielh P, et al. Regulation of gap junctions in melanoma and their impact on Melan-A/MART-1-specific CD8+ T lymphocyte emergence. J Mol Med (Berl) (2013) 91:1207–20. doi:10.1007/s00109-013-1058-5
158. Wahl DR, Byersdorfer CA, Ferrara JL, Opipari AW Jr, Glick GD. Distinct metabolic programs in activated T cells: opportunities for selective immunomodulation. Immunol Rev (2012) 249:104–15. doi:10.1111/j.1600-065X.2012.01148.x
159. Joseph N, Reicher B, Barda-Saad M. The calcium feedback loop and T cell activation: how cytoskeleton networks control intracellular calcium flux. Biochim Biophys Acta (2014) 1838:557–68. doi:10.1016/j.bbamem.2013.07.009
160. Janji B, Viry E, Moussay E, Paggetti J, Arakelian T, Mgrditchian T, et al. The multifaceted role of autophagy in tumor evasion from immune surveillance. Oncotarget (2016) 7:17591–607. doi:10.18632/oncotarget.7540
161. Catarino S, Ramalho JS, Marques C, Pereira P, Girão H. Ubiquitin-mediated internalization of connexin43 is independent of the canonical endocytic tyrosine-sorting signal. Biochem J (2011) 437:255–67. doi:10.1042/BJ20102059
162. Naus CC, Laird DW. Implications and challenges of connexin connections to cancer. Nat Rev Cancer (2010) 10:435–41. doi:10.1038/nrc2841
163. Mehta PP, Bertram JS, Loewenstein WR. Growth inhibition of transformed cells correlates with their junctional communication with normal cells. Cell (1986) 44:187–96. doi:10.1016/0092-8674(86)90497-6
164. Zhang X, Liu Y, Si YJ, Chen XH, Li ZJ, Gao L, et al. Effect of Cx43 gene-modified leukemic bone marrow stromal cells on the regulation of Jurkat cell line in vitro. Leuk Res (2012) 36:198–204. doi:10.1016/j.leukres.2011.10.001
165. Yang S, Wen Q, Liu Y, Zhang C, Wang M, Chen G, et al. Increased expression of CX43 on stromal cells promotes leukemia apoptosis. Oncotarget (2015) 6:44323–31. doi:10.18632/oncotarget.6249
166. Thuringer D, Jego G, Berthenet K, Hammann A, Solary E, Garrido C. Gap junction-mediated transfer of miR-145-5p from microvascular endothelial cells to colon cancer cells inhibits angiogenesis. Oncotarget (2016) 7:28160–8. doi:10.18632/oncotarget.8583
167. Zhou JZ, Riquelme MA, Gu S, Kar R, Gao X, Sun L, et al. Osteocytic connexin hemichannels suppress breast cancer growth and bone metastasis. Oncogene (2016) 35:5597–607. doi:10.1038/onc.2016.101
168. Dilber MS, Abedi MR, Christensson B, Björkstrand B, Kidder GM, Naus CC, et al. Gap junctions promote the bystander effect of herpes simplex virus thymidine kinase in vivo. Cancer Res (1997) 57:1523–8.
169. Fick J, Barker FG II, Dazin P, Westphale EM, Beyer EC, Israel MA. The extent of heterocellular communication mediated by gap junctions is predictive of bystander tumor cytotoxicity in vitro. Proc Natl Acad Sci U S A (1995) 92:11071–5. doi:10.1073/pnas.92.24.11071
170. Trepel M, Stoneham CA, Eleftherohorinou H, Mazarakis ND, Pasqualini R, Arap W, et al. A heterotypic bystander effect for tumor cell killing after adeno-associated virus/phage-mediated, vascular-targeted suicide gene transfer. Mol Cancer Ther (2009) 8:2383–91. doi:10.1158/1535-7163.MCT-09-0110
171. Lee HK, Finniss S, Cazacu S, Bucris E, Ziv-Av A, Xiang C, et al. Mesenchymal stem cells deliver synthetic microRNA mimics to glioma cells and glioma stem cells and inhibit their cell migration and self-renewal. Oncotarget (2013) 4:346–61. doi:10.18632/oncotarget.868
172. Soares AR, Martins-Marques T, Ribeiro-Rodrigues T, Ferreira JV, Catarino S, Pinho MJ, et al. Gap junctional protein Cx43 is involved in the communication between extracellular vesicles and mammalian cells. Sci Rep (2015) 5:13243. doi:10.1038/srep13243
173. Martins-Marques T, Pinho MJ, Zuzarte M, Oliveira C, Pereira P, Sluijter JP, et al. Presence of Cx43 in extracellular vesicles reduces the cardiotoxicity of the anti-tumour therapeutic approach with doxorubicin. J Extracell Vesicles (2016) 5:32538. doi:10.3402/jev.v5.32538
174. Ito A, Katoh F, Kataoka TR, Okada M, Tsubota N, Asada H, et al. A role for heterologous gap junctions between melanoma and endothelial cells in metastasis. J Clin Invest (2000) 105:1189–97. doi:10.1172/JCI8257
175. Saito-Katsuragi M, Asada H, Niizeki H, Katoh F, Masuzawa M, Tsutsumi M, et al. Role for connexin 26 in metastasis of human malignant melanoma: communication between melanoma and endothelial cells via connexin 26. Cancer (2007) 110:1162–72. doi:10.1002/cncr.22894
176. Villares GJ, Dobroff AS, Wang H, Zigler M, Melnikova VO, Huang L, et al. Overexpression of protease-activated receptor-1 contributes to melanoma metastasis via regulation of connexin 43. Cancer Res (2009) 69:6730–7. doi:10.1158/0008-5472.CAN-09-0300
177. Pollmann MA, Shao Q, Laird DW, Sandig M. Connexin 43 mediated gap junctional communication enhances breast tumor cell diapedesis in culture. Breast Cancer Res (2005) 7:R522–34. doi:10.1186/bcr1042
178. Elzarrad MK, Haroon A, Willecke K, Dobrowolski R, Gillespie MN, Al-Mehdi AB. Connexin-43 upregulation in micrometastases and tumor vasculature and its role in tumor cell attachment to pulmonary endothelium. BMC Med (2008) 6:20. doi:10.1186/1741-7015-6-20
179. Zibara K, Awada Z, Dib L, El-Saghir J, Al-Ghadban S, Ibrik A, et al. Anti-angiogenesis therapy and gap junction inhibition reduce MDA-MB-231 breast cancer cell invasion and metastasis in vitro and in vivo. Sci Rep (2015) 5:12598. doi:10.1038/srep12598
180. Thuringer D, Berthenet K, Cronier L, Solary E, Garrido C. Primary tumor- and metastasis-derived colon cancer cells differently modulate connexin expression and function in human capillary endothelial cells. Oncotarget (2015) 6:28800–15. doi:10.18632/oncotarget.4894
181. Thuringer D, Boucher J, Jego G, Pernet N, Cronier L, Hammann A, et al. Transfer of functional microRNAs between glioblastoma and microvascular endothelial cells through gap junctions. Oncotarget (2016) 7:73925–34. doi:10.18632/oncotarget.12136
182. Bazarbachi A, Abou Merhi R, Gessain A, Talhouk R, El-Khoury H, Nasr R, et al. Human T-cell lymphotropic virus type I-infected cells extravasate through the endothelial barrier by a local angiogenesis-like mechanism. Cancer Res (2004) 64:2039–46. doi:10.1158/0008-5472.CAN-03-2390
183. Haddad L, El Hajj H, Abou-Merhi R, Kfoury Y, Mahieux R, El-Sabban M, et al. KSHV-transformed primary effusion lymphoma cells induce a VEGF-dependent angiogenesis and establish functional gap junctions with endothelial cells. Leukemia (2008) 22:826–34. doi:10.1038/sj.leu.2405081
184. Karpinich NO, Caron KM. Gap junction coupling is required for tumor cell migration through lymphatic endothelium. Arterioscler Thromb Vasc Biol (2015) 35:1147–55. doi:10.1161/ATVBAHA.114.304752
185. Tang B, Peng ZH, Yu PW, Yu G, Qian F, Zeng DZ, et al. Aberrant expression of Cx43 is associated with the peritoneal metastasis of gastric cancer and Cx43-mediated gap junction enhances gastric cancer cell diapedesis from peritoneal mesothelium. PLoS One (2013) 8:e74527. doi:10.1371/journal.pone.0074527
186. Paraguassú-Braga FH, Borojevic R, Bouzas LF, Barcinski MA, Bonomo A. Bone marrow stroma inhibits proliferation and apoptosis in leukemic cells through gap junction-mediated cell communication. Cell Death Differ (2003) 10:1101–8. doi:10.1038/sj.cdd.4401279
187. Lim PK, Bliss SA, Patel SA, Taborga M, Dave MA, Gregory LA, et al. Gap junction-mediated import of microRNA from bone marrow stromal cells can elicit cell cycle quiescence in breast cancer cells. Cancer Res (2011) 71:1550–60. doi:10.1158/0008-5472.CAN-10-2372
188. Farina C, Aloisi F, Meinl E. Astrocytes are active players in cerebral innate immunity. Trends Immunol (2007) 28:138–45. doi:10.1016/j.it.2007.01.005
189. Valiente M, Obenauf AC, Jin X, Chen Q, Zhang XH, Lee DJ, et al. Serpins promote cancer cell survival and vascular co-option in brain metastasis. Cell (2014) 156:1002–16. doi:10.1016/j.cell.2014.01.040
190. Lin JH, Takano T, Cotrina ML, Arcuino G, Kang J, Liu S, et al. Connexin 43 enhances the adhesivity and mediates the invasion of malignant glioma cells. J Neurosci (2002) 22:4302–11.
191. Oliveira R, Christov C, Guillamo JS, de Boüard S, Palfi S, Venance L, et al. Contribution of gap junctional communication between tumor cells and astroglia to the invasion of the brain parenchyma by human glioblastomas. BMC Cell Biol (2005) 6(1):7. doi:10.1186/1471-2121-6-7
192. Sin WC, Aftab Q, Bechberger JF, Leung JH, Chen H, Naus CC. Astrocytes promote glioma invasion via the gap junction protein connexin43. Oncogene (2016) 35:1504–16. doi:10.1038/onc.2015.210
193. Lin Q, Liu Z, Ling F, Xu G. Astrocytes protect glioma cells from chemotherapy and upregulate survival genes via gap junctional communication. Mol Med Rep (2016) 13:1329–35. doi:10.3892/mmr.2015.4680
194. Hong X, Sin WC, Harris AL, Naus CC. Gap junctions modulate glioma invasion by direct transfer of microRNA. Oncotarget (2015) 6:15566–77. doi:10.18632/oncotarget.3904
195. Pan J, Thoeni C, Muise A, Yeger H, Cutz E. Multilabel immunofluorescence and antigen reprobing on formalin-fixed paraffin-embedded sections: novel applications for precision pathology diagnosis. Mod Pathol (2016) 29:557–69. doi:10.1038/modpathol.2016.52
Keywords: gap junction, connexin, tumor immunity, tumor microenvironment, stromal cells
Citation: Gleisner MA, Navarrete M, Hofmann F, Salazar-Onfray F and Tittarelli A (2017) Mind the Gaps in Tumor Immunity: Impact of Connexin-Mediated Intercellular Connections. Front. Immunol. 8:1067. doi: 10.3389/fimmu.2017.01067
Received: 11 July 2017; Accepted: 16 August 2017;
Published: 01 September 2017
Edited by:
Masoud H. Manjili, Virginia Commonwealth University, United StatesReviewed by:
Fabian Benencia, Ohio University, United StatesMallikarjun Bidarimath, Cornell University College of Veterinary Medicine, United States
Copyright: © 2017 Gleisner, Navarrete, Hofmann, Salazar-Onfray and Tittarelli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Flavio Salazar-Onfray, fsalazar@u.uchile.cl;
Andrés Tittarelli, tittarelli@gmail.com