 Jouni Jokela1
Jouni Jokela1 Lassi M. P. Heinilä1
Lassi M. P. Heinilä1 Tânia K. Shishido1
Tânia K. Shishido1 Matti Wahlsten1
Matti Wahlsten1 David P. Fewer1
David P. Fewer1 Marli F. Fiore2Hao Wang1Esa Haapaniemi3
Marli F. Fiore2Hao Wang1Esa Haapaniemi3 Perttu Permi3,4
Perttu Permi3,4 Kaarina Sivonen1*
Kaarina Sivonen1*- 1Department of Food and Environmental Sciences, University of Helsinki, Helsinki, Finland
- 2Center for Nuclear Energy in Agriculture, University of São Paulo, Piracicaba, Brazil
- 3Department of Chemistry, University of Jyväskylä, Helsinki, Finland
- 4Department of Biological and Environmental Science, Nanoscience Center, University of Jyväskylä, Helsinki, Finland
Nostoc is a cyanobacterial genus, common in soils and a prolific producer of natural products. This research project aimed to explore and characterize Brazilian cyanobacteria for new bioactive compounds. Here we report the production of hepatotoxins and new protease inhibitors from benthic Nostoc sp. CENA543 isolated from a small, shallow, saline-alkaline lake in the Nhecolândia, Pantanal wetland area in Brazil. Nostoc sp. CENA543 produces exceptionally high amounts of nodularin-R. This is the first free-living Nostoc that produces nodularin at comparable levels as the toxic, bloom-forming, Nodularia spumigena. We also characterized pseudospumigins A–F, which are a novel family of linear tetrapeptides. Pseudospumigins are structurally related to linear tetrapeptide spumigins and aeruginosins both present in N. spumigena but differ in respect to their diagnostic amino acid, which is Ile/Leu/Val in pseudospumigins, Pro/mPro in spumigins, and Choi in aeruginosins. The pseudospumigin gene cluster is more similar to the spumigin biosynthetic gene cluster than the aeruginosin gene cluster. Pseudospumigin A inhibited trypsin (IC50 4.5 μM after 1 h) in a similar manner as spumigin E from N. spumigena but was almost two orders of magnitude less potent. This study identifies another location and environment where the hepatotoxic nodularin has the potential to cause the death of eukaryotic organisms.
Introduction
Nostoc is a cosmopolitan genus of cyanobacteria that is commonly found in soil and in symbiotic associations with plants and fungi. Members of the genus Nostoc are prolific producers of natural products including peptides, polyketides, and alkaloids (Kobayashi and Kajiyama, 1998; Dittmann et al., 2015). Free-living and symbiotic strains of Nostoc are known to produce a range of microcystins, potent cyclic heptapeptide hepatotoxins produced by a number of cyanobacteria (Sivonen et al., 1990; Oksanen et al., 2004; Genuário et al., 2010; Kaasalainen et al., 2012; Figure 1). Nodularin (NOD) is a hepatotoxic protein phosphatase inhibitor and tumor promoter that has two amino acid residues fewer than microcystins (Figure 1). More than 240 structural variants have been reported for microcystins but only 10 structural variants of NOD are known to date (Meriluoto et al., 2017). This disparity can be partly explained by the most variable amino acid in microcystins being absent from NODs. Sivonen et al. (1990) had reported that Nostoc is one of the cyanobacterial genera that produce microcystins and since then microcystin production have been reported to occur in several other Nostoc strains (Oksanen et al., 2004; Genuário et al., 2010; Kaasalainen et al., 2012). NOD was detected only in free living Nodularia until 2012 when NOD was also found in cycad endosymbiotic Nostoc strains in low concentrations (<0.4 μg/g biomass) and also in liverwort symbiotic Nostoc sp. SKS8 (Gehringer et al., 2012; Liaimer et al., 2016). Higher amounts of NOD have been found among the cyanolichens, in which Nostoc is the most common cyanobacterial endosymbiont (Kaasalainen et al., 2012).
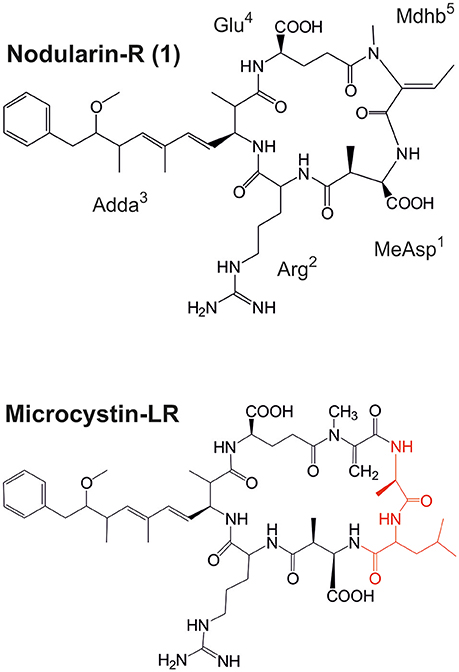
Figure 1. Structures of nodularin-R and microcystin-LR, which is the most common variant of microcystin (amino acids marked red are not present in nodularins). MeAsp, D-erythro-β-methylaspartic; Adda, 3-amino-9-methoxy-10-phenyl-2,6,8-trimethyl-deca-4(E),6(E)-dienoic acid; Mdhb, N-methyl-dehydrobutyric acid.
The cyanobacterium Nodularia spumigena that forms blooms in brackish water can produce NOD at high amounts, >5 mg NOD/g biomass dry weight (DW; Repka et al., 2001). These blooms constitute a health risk for human and domestic animals (Sivonen, 2009; Simola et al., 2012). The latest report of a new NOD producer is from tropical Australia, where a new cyanobacterial species Iningainema pulvinus gen nov., sp. nov. was isolated from a freshwater wetland spring (McGregor and Sendall, 2017). I. pulvinus is the third genus from which NOD production has been reported. The mean level was 0.9 mg NOD g−1 biomass DW, which is over three orders of magnitude higher than the amount of NOD previously reported from Nostoc (Gehringer et al., 2012).
Spumigins and aeruginosins are linear peptide protease inhibitors that comprise three amino acids and a terminal carboxylic acid residue (Fewer et al., 2009, 2013). The presence or absence of the diagnostic amino acids, Pro/mPro in spumigins and the amino acid Choi (2-carboxy-6-hydroxyoctahydroindole) in aeruginosins, are the chemical determining entities that differentiates these two peptide groups (Figure 2). These peptide groups are products of non-ribosomal peptide synthetase biosynthetic pathways that do share a great deal of structural similarities (Fewer et al., 2009, 2013). Spumigins were first described in N. spumigena AV1 (Fujii et al., 1997). Twenty-four spumigin congeners in total have been described. Spumigins A-I and nine other spumigins have been described from N. spumigena strains isolated in the Baltic Sea and also in Turkish freshwater lake (Fujii et al., 1997; Fewer et al., 2009; Mazur-Marzec et al., 2013). Moreover, spumigin J from Anabaena compacta isolated from Lake Esthwaite Water in England (Anas et al., 2012) and four spumigin congeners from Sphaerospermopsis torques-reginae cyanobacterium isolated from a bloom sample collected in the Tapacurá reservoir in Brazil have been described (Sanz et al., 2015). Aeruginosins are more common and roughly 100 variants that show a higher structural variation compared to spumigins have been described (Dittmann et al., 2015). Aeruginosin-865 was the first variant found in the genus Nostoc and was isolated from the strain Lukešová 30/93 growing in forest soil (Kapuścik et al., 2013). Pseudoaeruginosins are a third group of similar linear peptides which contain mPro instead of Choi (Liu et al., 2015; Figure 2).
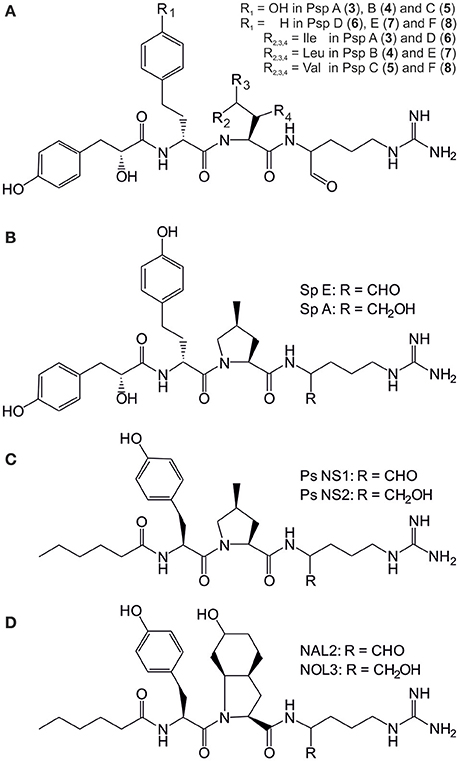
Figure 2. Structures of the pseudospumigins (Psp) A–E (3–8) produced by Nostoc sp. CENA543 (A) and Nodularia spumigena produced spumigins (Sp) E and A (B), pseudoaeruginosins (Ps) NS1 and NS2 (C) and aeruginosins NAL2 and NOL3 (D).
The non-ribosomal peptidome of N. spumigena typically includes nodularins, spumigins, aeruginosins, anabaenopeptins, and sometimes pseudoaeruginosins (Fewer et al., 2013; Mazur-Marzec et al., 2013; Liu et al., 2015). The corresponding biosynthetic gene clusters have been described from N. spumigena CCY9414 (Fewer et al., 2009, 2013; Voß et al., 2013).
In the current study we analyzed non-ribosomal peptides from nine Nostoc and nine other cyanobacterial genera that had been collected from Brazilian saline-alkaline lakes of the Nhecolândia, Pantanal wetland area. Our objective was to discover novel bioactive compounds. Surprisingly Nostoc sp. CENA543 produced NOD in quantities comparable to those produced by toxic N. spumigena strains that had been found in various geographical locations. A new peptide family pseudospumigins (Psp) expressed by Nostoc sp. CENA543 was identified (Table 1). The structure of Psps are identical with that of spumigins but the diagnostic amino acid Pro/mPro that are found in spumigins were replaced by Ile, Leu, or Val amino acids and hence the novel family were named the pseudospumigins. We found that the Psp family are trypsin inhibitors.
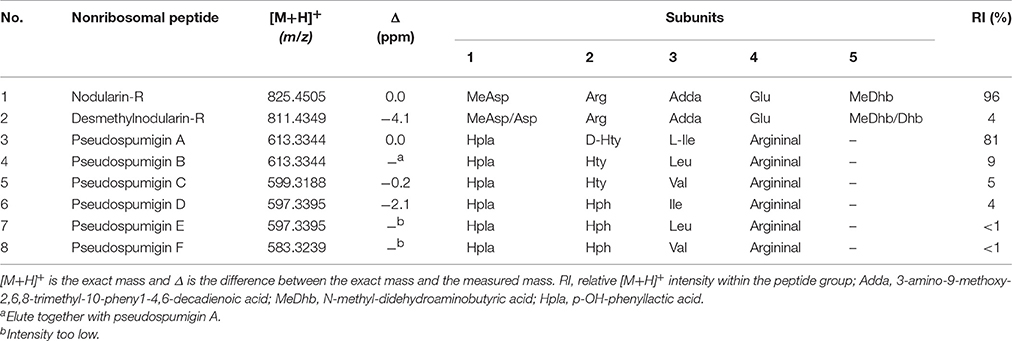
Table 1. Nostoc sp. CENA543 non-ribosomal peptides.
Materials and Methods
Strains and Cultivation
The cyanobacterial strains studied were isolated from water samples that had been collected from several saline-alkaline lakes located at Centenário farm in the southern part of the sub-region Nhecolândia in the north of the municipality of Aquidauana, Mato Grosso do Sul State, Brazil (Andreote et al., 2014; Genuário et al., 2017; Figure S1). The Nostoc sp. strains CENA543 and CENA544 were isolated from water samples collected in September 3, 2010 from the lake “Salina 67 Mil” (19°27′42″S, 56°08′21″W). An additional 7 Nostoc and 9 other cyanobacterial strains were also collected in May 27 or September 3, 2010 from the lakes “Salina 67 Mil,” “Salina Verde” (19°28′13″S, 56°03′22″W), “Salina Preta” (19°26′56″S, 56°07′55″W), “Salina Grande” (19°26′56″S, 56°07′45″W) and “Salina Centenário” (19°26′24″S, 56°05′58″W). The CENA543 strain was purified axenic before peptide analysis and DNA isolation. All strains were cultivated at ambient temperature (20–22°C), in continuous low photon irradiance (5–10 μE m−2 s−1), in low salinity (0.6‰) but at high phosphorus concentration (5,500 μg -P L−1) on Z8 medium (Kotai, 1972) without a nitrogen source for a few weeks. The reference strain N. spumigena AV1 was cultivated as previously described (Fewer et al., 2009). The effect of 0, 1, 2, 3, 4, 5, and 6% salinities adjusted with NaCl to the growth of Nostoc sp. CENA543 was tested on Z8 medium. A 50 ml volume of medium was placed into 250 ml plastic cell cultivation flasks, each of which was inoculated with 1 ml of gently homogenized cell suspension that had been grown for 28 days on Z8 medium without a nitrogen source. Flasks were cultivated for 34 days in the horizontal position with 80 rpm shaking speed at 18°C. Nostoc sp. CENA543 was mass cultivated in 5 L Erlenmeyer flaks containing 2.7 L of Z8 medium with sterilized air bubbling for NMR, amino acid analysis and enzyme inhibition tests. The DNA extraction required to Nostoc sp. CENA543 be grown in 1% salinity for 16 days (addition of 8.75 g NaCl L−1 and 3.75 g MgSO4·7H2O L−1) in Z8 medium without a nitrogen source to reduce the slime formation, which made DNA isolation difficult.
Peptide Recognition by LC-MS
Cells were grown in 40 ml of liquid cultures, collected and freeze dried. The dried biomasses were placed in 2 ml plastic tubes together with 1 ml methanol and glass beads (0.5-mm diameter glass beads, Scientific Industries INC) and were shaken using FastPrep cell disrupter instrument three times for 30 s at a speed of 6.5 m·s−1. Tubes were then centrifuged 10,000 × g for 5 min at room temperature. Supernatants were analyzed first using low resolution LC-ESI-ITMS (Agilent 1100 Series LC/MSD Ion Trap XCT Plus, Agilent Technologies, Palo Alto, CA, USA). A 10 μl sample was injected into a Luna C18 column (2.1 × 100 mm, 5 μm, Phenomenex, Torrance, CA, USA) which was eluted by 30% acetonitrile (solvent B) in 0.1% HCOOH to 70% of B at 40°C for 49 min with a flow rate of 0.15 ml min−1. Mass spectra data were accumulated in ultrascan positive electrospray ionization mode (26,000 m/z s−1) at a scan range of m/z 300–2,200 and by calculating the mean of three spectra.
High resolution UPLC-QTOF analyses were performed using an Acquity I-Class UPLC—Synapt G2-Si HDMS (Waters Corp., Milford, MA, USA) system. Samples that ranged from 0.1 to 1 μl were injected into a Cortecs UPLC® C18+ column (2.1 × 50 mm, 1.6 μm, Waters), which was then eluted at 40°C with a flow rate of 0.3 ml min−1 from 10% acetonitrile (+0.1% HCOOH; solvent B) in 0.1% HCOOH to 70% of B in 5 min, then to 95% of B in 0.01 min, kept there 1.99 min, then back to 10% of B in 0.5 min and finally kept for 2.5 min before the next run. QTOF was calibrated using sodium formate, which gave a calibrated mass range from m/z 91.036 to 472.726 or from 1178.651 to 2121.195 depending of the run. Leucine Enkephalin was used at 10 s intervals as a lock mass reference compound. Mass spectral data were accumulated in positive electrospray ionization Resolution Mode at scan range of m/z 50–200 or 1,500–2,200 depending of the run.
Pseudospumigin A Purification
Pseudospumigin A was purified for NMR, amino acid analysis and the enzyme inhibition test. One gram of freeze dried Nostoc sp. CENA543 cells were lysed in a 25 ml volume of methanol in a 50 ml plastic tube for 30 s using a Silentcrusher M homogenizer (Heidolph Instruments GmbH & Co, Germany) at 20,000 rpm speed. After centrifugation 10,000 × g for 5 min at room temperature, the supernatant was removed and the extraction was repeated once with 20 ml of methanol. The combined methanol solutions were diluted with water to 80% methanol concentration. Solution was added into a methanol (20 ml) preconditioned solid phase C18-E cartridge (5 g/20 ml, Phenomenex, Torrance, CA, USA). The effluent was first evaporated by a vacuum rotatory evaporator and then freeze dried. The residue was dissolved in 15 ml methanol and 450 mg of NaBH4 was added to reduce the peptide aldehydes to chromatographically better behaving alcohols. After gas formation the methanol was vacuum rotatory evaporated. The residue was dissolved in 2 ml of eluent then used in pseudospumigins purification performed by an Agilent LC-ESI-ITMS. The solution was injected in 100–500 μl sample batches with a manual injector into a Luna C8(2) column (10 × 150 mm, 5 μm, Phenomenex, Torrance, CA, USA) eluted isocratically with solvent mixture of 1% aqueous ammonium acetate and acetonitrile (22:78) at speed of 5 ml min−1. Collected fractions were pooled then diluted in 1:2 with water and concentrated with preconditioned (first 6 ml methanol then 6 ml water) solid phase Strata-X cartridge (1 g/6 ml, Phenomenex). The cartridge was first washed with 6 ml water which was then blown out with compressed air and pseudospumigins (Psp A fraction) were eluted from the cartridge with 3 ml of methanol yielding 0.87 mg of freeze dried solid material.
NMR and UV Spectroscopy
NMR spectra for structural elucidation of pseudospumigin A were collected using an Avance III HD 500 MHz NMR spectrometer equipped with 1H, 13C, 15N TXI probe and an Avance III HD 800 MHz NMR spectrometer equipped with 1H, 13C, 15N TCI cryoprobe. All spectra were collected at room temperature. The COSY experiment at 500 MHz were obtained with 8 transients, using 256 and 1,024 complex points in t1 and t2 domains that corresponded to acquisition times of 46.6 and 186.2 ms, respectively. At 800 MHz, the 1H spectrum was collected using 16 transients and 16 k complex points that corresponded to an acquisition time of 1.278 s. The sensitivity enhanced 13C-HSQC (15N-HSQC) spectrum was obtained using 12832 and 1,024 complex points in t1 (t1) and t2 dimensions. This results in acquisition times of 3.53 ms (1.6 ms) and 79.9 ms in 13C (15N) and 1H dimensions, respectively. The signal was accumulated with 32 (160) transients per FID in 13C-HSQC (15N-HSQC) experiment. The data for heteronuclear multiple bond correlation (13C-HMBC) spectrum was collected with 128 and 2,048 complex points in t1 and t2 domains, using 160 transients per FID. This translates to acquisition times of 2.9 and 213 ms in 13C and 1H dimensions, respectively. The long-range 1H-13C transfer delay was optimized according to 6 Hz nJCH couplings. UV (MeOH) λmax (log ε) 225 (4.23), 278 (3.49) nm.
Nodularin-R Quantitation
A 10.14 mg quantity of freeze dried Nostoc sp. CENA543 and a 12.45 mg of freeze dried sample of N. spumigena AV1 cells were extracted with 1 ml methanol as previously explained. A 50-fold methanol diluted extracts together with eight nodularin a (a gift from Z. Grzonka, University of Gdansk, Gdansk, Poland) standard methanol solutions (2-fold dilutions from concentration 2 to 0.0156 μg ml−1) were analyzed by UPLC-QTOF using negative ionization. Nodularin peak areas of m/z 823.432 ± 0.01 ion chromatograms were measured. Nodularin gave a standard curve y = 1010x2 + 2540x − 58.6 (y = peak area, x = nodularin concentration) with R2 = 0.9999 from which the Nostoc sp. CENA543 and N. spumigena AV1 nodularin concentrations were calculated.
Amino Acid Analysis
A 50 μg quantity of purified Psp A fraction in a 200 μl glass tube inside a closed 4 ml vial containing 500 μl of 6 M HCl was hydrolyzed at 110°C for 12 h. The glass tube content was vacuum dried and 50 μl of water, 20 μl of 1 M NaHCO3, and 100 μl of 1% Marfey reagent in acetone (1-fluoro-2,4-dinitrophenyl-5-L-alanine or L-leucine amide) was added. The reaction was stopped after 1 h of incubation at 37°C by adding 20 μl of 1 M HCl and the solution was analyzed by UPLC-QTOF. The reference compounds from which L- and D-Leu, L-, D-, and L-allo-Ile, L- and D-Val and DL-Hph were obtained from Sigma-Aldrich (Switzerland), whereas L- and D-Hty were obtained from ABCR GmbH & Co. (Germany) and were Marfey derivatized accordingly.
IC50 Measurement
Trypsin (porcine pancreas, Sigma-Aldrich) activity was measured at 25°C in a reaction mixture that contained from 0 to 81.6 mM leupeptin or pseudospumigin A and the hydrolysis was followed at every 2 min for 1 h, as described earlier (Liu et al., 2015).
DNA Extraction and Genome Sequencing
Cells were collected in a 50 ml centrifuge tube and centrifuged at 6,000 × g for 5 min. Cells were washed with 2 ml of Tris-EDTA-NaCl-buffer (50 mM Tris-HCl—100 mM EDTA—100 mM NaCl) and transferred to microcentrifuge tubes. The tubes were centrifuged at 7,000 × g for 4 min and the supernatant was discarded. Glass beads were added to the tubes and the cells were frozen at −80°C. Samples were thawed at 64°C and 800 μl of GOS-buffer (100 mM TrisHCl, pH 8, 1.5% SDS, 10 mM EDTA, 1% deoxycholate, 1% Igepal-CA630, 5 mM thiourea, 10 mM dithiothreitol; Kolmonen et al., 2004) was added to the cells. The cells were then disrupted using FastPrep at 5 m.s−1 for 20 s. The samples were kept on ice for 5 min and centrifuged at 15,000 × g for 1 min. The supernatant was pipetted in a 15 ml centrifuge tube. The disruption of the cells and centrifugation steps were repeated with additional 800 μl of GOS-buffer. The supernatants were combined by decanting into the 15 ml centrifuge tube. A volume of 225 μl of 5 M NaCl water solution and 170 μl of 10% CTAB (10% CTAB–0.7 M NaCl) was added to each 800 μl of buffer. The sample was then divided amongst four tubes, each of which was mixed and incubated at 65°C for 20 min. An equal volume of phenol-chloroform (1:1) was added. Samples were mixed and centrifuged at 10,000 × g for 7 min. The water phases that contained the DNA was transferred into fresh tubes and phenol-chloroform treatment was repeated. The isopropanol (5 mL) was added to the tubes, which were mixed and centrifuged at 10,000 × g for 15 min at 4°C. The supernatant was discarded and the pellet was dissolved in 1 ml of 70% ethanol. The samples were combined and centrifuged at 10,000 × g for 5 min. The supernatant was discarded and the samples were dried at 37°C and stored at −20°C.
The DNA pellet was dissolved in 850 μl of TE-buffer and 1.6 μl of RNAse A was added. Each sample was incubated at 37°C for 30 min. A saline solution of 5 M NaCl was added to a final concentration of 0.2 M (32 μl). An equal volume of phenol-chloroform was added and the sample was centrifuged at 20,000 × g for 5 min. The water phase that contained the DNA was transferred into a new tube and an equal volume of chloroform was added and the sample was centrifuged at 16,000 × g for 5 min. The DNA sample was transferred to a new tube and an equal volume of isopropanol was added and centrifuged at 10,000 × g at 4°C for 10 min. The supernatant was discarded and the pellet was washed with 70% ethanol followed by drying at 37°C. The DNA pellet was dissolved in 300 μl of TE-buffer. The purity, concentration and quality of the DNA was measured and evaluated using Nanodrop ND-1000 Spectrophotometer (Nanodrop Technologies, USA), gel electrophoresis and an Agilent TapeStation (Agilent Technologies). High-molecular DNA was subjected to library (Illumina TruSeq® PCR Free 350 bp) construction and sequenced by using an Illumina HiSeq2500 platform with a paired ends 100 cycles run. The gaps of the gene cluster were closed by PCR and Sanger sequencing.
Biosynthetic Gene Cluster Analysis
The genome data (1 Gb) were first checked by the Spades software (version 3.7.1) for read correction and removal of erroneous readings, and then the data were assembled using the Newbler program (version 3.0). The genome sequence obtained for Nostoc sp. CENA543 was analyzed using antiSMASH (Blin et al., 2017) and annotated using Artemis software (Rutherford et al., 2000). The sequence was analyzed for the NRPS/PKS content using PKS/NRPS Analysis (Bachmann and Ravel, 2009) and the substrate prediction of the adenylation domain was done using NRPS predictor 2 (Rausch et al., 2005; Röttig et al., 2011) and manual alignment. The phylogenetic analysis was performed in the Molecular Evolutionary Genetics Analysis (MEGA 6.06; Tamura et al., 2013). A phylogenetic tree was constructed using Neighbor-joining (16S rRNA genes—K2+G, ndaF gene—K2+G, SpuA and SpuB amino acids—Poisson model +G) and Maximum likelihood (16S rRNA genes—K2+G+I) methods.
Results
Growth of Nostoc sp. CENA 543 and Identification of Peptides
We studied the non-ribosomal peptides of the cyanobacterium Nostoc sp. CENA543 isolated from a saline-alkaline lake in the Pantanal wetland area of Brazil (Figure S1). A growth experiment demonstrated that Nostoc sp. CENA543 grew best at low (0.6‰) salt concentration despite, some occasionally high salt concentrations (4.5%) that were encountered in the lake (Figure S1). Growth at 1% salinity was 10 times slower and growth ceased completely at 2% salinity. Therefore, we cultivated this strain in low salinity and light but at high phosphorus concentrations in order to obtain good growth and peptide expression. Nostoc sp. CENA543 grew on culture flask surfaces and showed a benthic growth profile (Figure S1).
A methanol extract of the culture was first analyzed by high performance liquid chromatography electro-spray ion trap mass spectrometry (HPLC-ITMS), which resulted to the identification of hepatotoxic nodularin-R in high quantities and lower quantities of desmethylnodularin-R (Figure 3, Figure S2). Quantitative analysis performed by using ultra performance liquid chromatography guadrupole time of flight (UPLC-QTOF) mass spectrometry determined that Nostoc sp. CENA543 produced 4.3 mg NOD g−1 biomass DW (20–22°C, 5–10 μE m−2 s−1, salinity 0.6‰, 5,500 μg PO4-P L−1), which was more than twice as much NOD production by N. spumigena AV1 (1.9 mg g−1). The chemical structures of the nodularin variants were assigned using the product ion spectra generated by UPLC-QTOF and HPLC-ITMS (Figure S2).
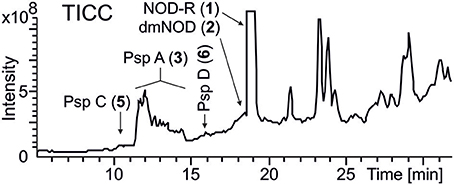
Figure 3. LC-ITMS total ion current chromatogram (TICC) of Nostoc sp. CENA543. Abbreviations as in Figure 1.
HPLC-ITMS analysis of the methanol extract also revealed that there was an early eluting compound group whose chromatographic behavior (broad tailing peaks) and MS fragmentation resembled those of the spumigin and aeruginosin aldehydes (Figure S2; Fewer et al., 2009, 2013). Analysis of the low and high resolution spectra showed that these pseudospumigins (Psps) are closely related to spumigins and aeruginosins that have Pro/mPro or Choi diagnostic amino acids but differ from them by the substitution with Ile/Leu or Val amino acids (Figures 2, 4, 5). Ion assignments of the high-resolution product ion spectrum of protonated Psp A, C, D, and F are presented in Table 2, Table S1, and in Figure S3. The subunit sequence of the Psp family was found to be Hpla1-Hty/Hph2-Ile/Leu/Val3-Argininal4. The aldehyde group of Psp's was seen also in the formation of hydrate (m/z 631.34 [M+H]+ for Psp A) and hemiacetal (m/z 645.36 [M+H]+ for Psp A) structures (Figure 5). Argininol4 containing variants were practically absent. Chiral amino acid analysis of the isolated Psp mixture revealed the amino acids D-Hty, D-Hph, L-Ile, L-Leu, and L-Val, although L-Hph was also detected (Figure S4). This finding together with the 1H-13C HSQC spectrum analysis showed that roughly one tenth of the Aa3 was L-Leu. This finding indicates that Hpla1-Hty2-Leu3-Argininal4 (Psp B) and Hpla1-Hph2-Leu3-Argininal4 (Psp E) variants were also present in the sample. The Hpla1-Hph2-Val3-Argininal4 variant (583 Da, Psp F) was detected only by the UPLC-QTOF analysis. The NMR data that are presented in Supplementary Material (Figure S5, Table S2) were in line with the published data from Sps, aeruginosins, and nostosins and confirmed the Psp structure. The relative amounts of the Psp entities that were calculated from the protonated Psp HPLC-ITMS peaks and NMR data are presented in Table 1.
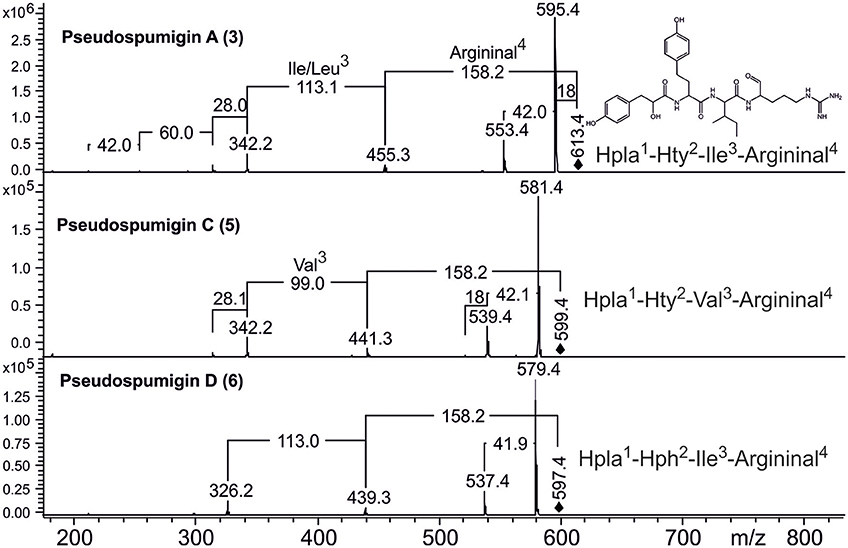
Figure 4. Product ion spectra and structures of pseudospumigins A (3), C (5), and D (6) identified from Nostoc sp. CENA543.
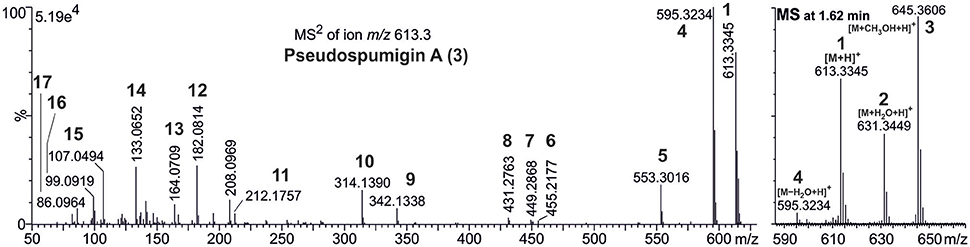
Figure 5. Accurate mass product ion spectrum (MS2) of protonated pseudospumigin (Psp) A (3) m/z 613.3 and mass spectrum (MS) from the chromatographic peak of Psp A identified from Nostoc sp. CENA543. Numbers in bold refer to the ion structures presented in Table 1.
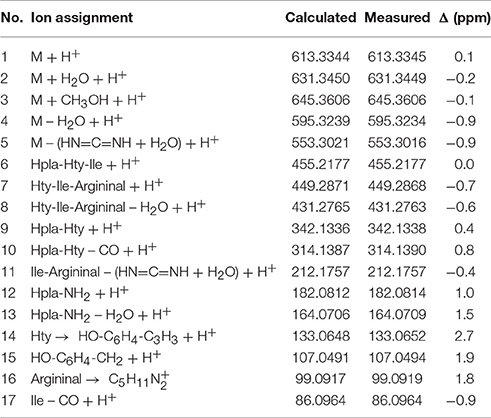
Table 2. Ion assignments, calculated, and measured ion masses and mass difference (Δ) of protonated pseudospumigin A (3) isolated from Nostoc sp. CENA543 analyzed by UPLC-QTOF.
The bioactivity of the isolated and purified Psp mixture from which about 80% is Psp A was tested with trypsin because many structurally analogous Sps, aeruginosins, pseudoaeruginosins, and the more distant nostosins homologs are all trypsin inhibitors. Psp A (mix) inhibited trypsin in a time dependent manner as Sp E15 (Figure 6). When trypsin activity was measured for different Psp A (mix) concentrations at 1 h, the IC50 value of Psp A (mix) was 4.5 μM. At that point the trypsin inhibition activity of Psp was 1/70th the activity of Sp E.
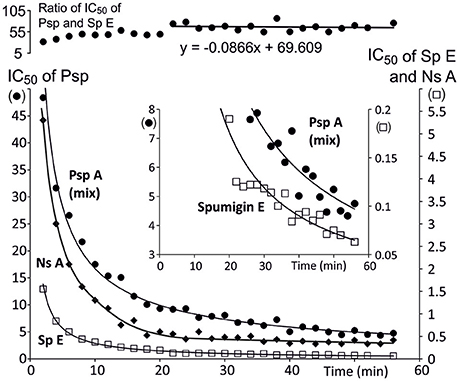
Figure 6. Time dependence of trypsin inhibition efficiency (IC50) of pseudospumigin A [•, Psp A (3) + other Psp variants], nostosin A (♦, Ns A) and spumigin E (□, Sp A, from Fewer et al., 2013). The uppermost curve shows that the ratio of IC50 of Psp mix and Sp E stabilizes and that trypsin inhibition by Psp mix is 1/70th the inhibition of spumigin E. The insert is an enlargement from the small IC50 values of Psp mix and Sp E clearly shows that pseudospumigin A (mix) and spumigin E trypsin interaction were not in equilibrium after 1 h of reaction time.
Both Nostoc strains of CENA543 and CENA544 that had been isolated from lake “Salina 67 Mil” had identical peptide profiles, whereas the third Salina 67 Mil sample, which was a non-Nostoc strain had a different profile. Moreover, all the other 18 Nostoc and non-Nostoc strains that had been collected from the nearby Nhecolândia Pantanal wetland lakes presented different peptidomes (Figures S6, S7).
Nostoc sp. CENA 543 Biosynthetic Gene Clusters
A 7.2 Mb draft genome sequence organized in 67 contigs was obtained from the Nostoc sp. CENA543 strain (data not shown). A complete 48 kb NOD biosynthetic gene cluster that encode hybrid NRPS-PKS enzymes was identified in the draft genome (Figure 7A). The Nostoc sp. CENA543 NOD biosynthetic gene cluster (GenBank accession number MF668122) was found to be similar to the NOD biosynthetic gene clusters of Nostoc sp. 73.1 and microcystin gene cluster of Anabaena sp. 90 (Table S3). The nodularin gene cluster of Nostoc sp. CENA543 comprised two NRPSs (NdaA and NdaB), two hybrid NRPS-PKS (NdaC and NdaF), a PKS (NdaD), an O-methyltransferase (NdaE), a racemase (NdaG), a dehydrogenase (NdaH), and an ABC transporter (NdaI; Table S3).
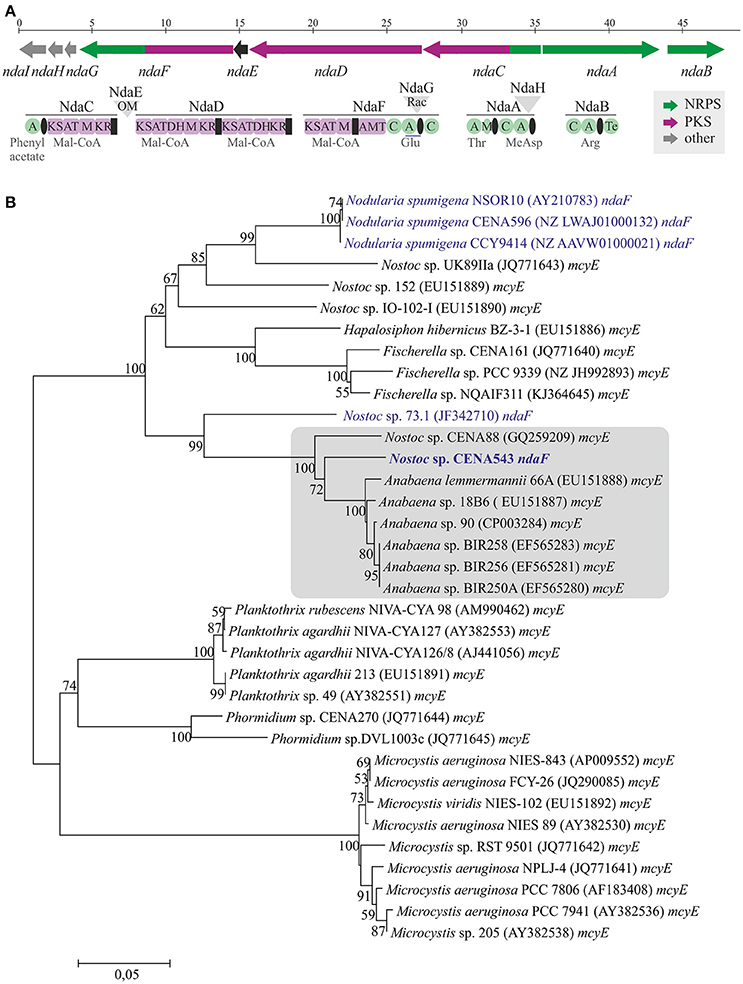
Figure 7. Nodularin biosynthetic gene cluster from Nostoc sp. CENA543 (A) and evolutionary history of ndaF and mcyE genes (B). Neighbor-joining tree based on ndaF and mcyE genes in the adenylation domain and peptidyl carrier protein region. Bootstrap values above 50 percent of 1,000 replicates are given at the nodes. Strains that produce nodularin are highlighted in blue and Nostoc sp. CENA543 is blue and bold.
The complete 19 kb Psp biosynthetic gene cluster was located through BLASTp search using spumigin biosynthetic enzymes as the query (Figure 8A and Table S3, GenBank accession number MF668123). The Psp biosynthetic gene cluster encodes two NRPS enzymes, SpuA, and SpuB in addition to the ABC transporter, SpuC (Figure 8A, Table S3). The amino acids predicted to be incorporated by the adenylation domains in the NRPS of nodularin and pseudospumigin were found to be partly consistent with the chemical structures obtained from these compounds (Table S4).
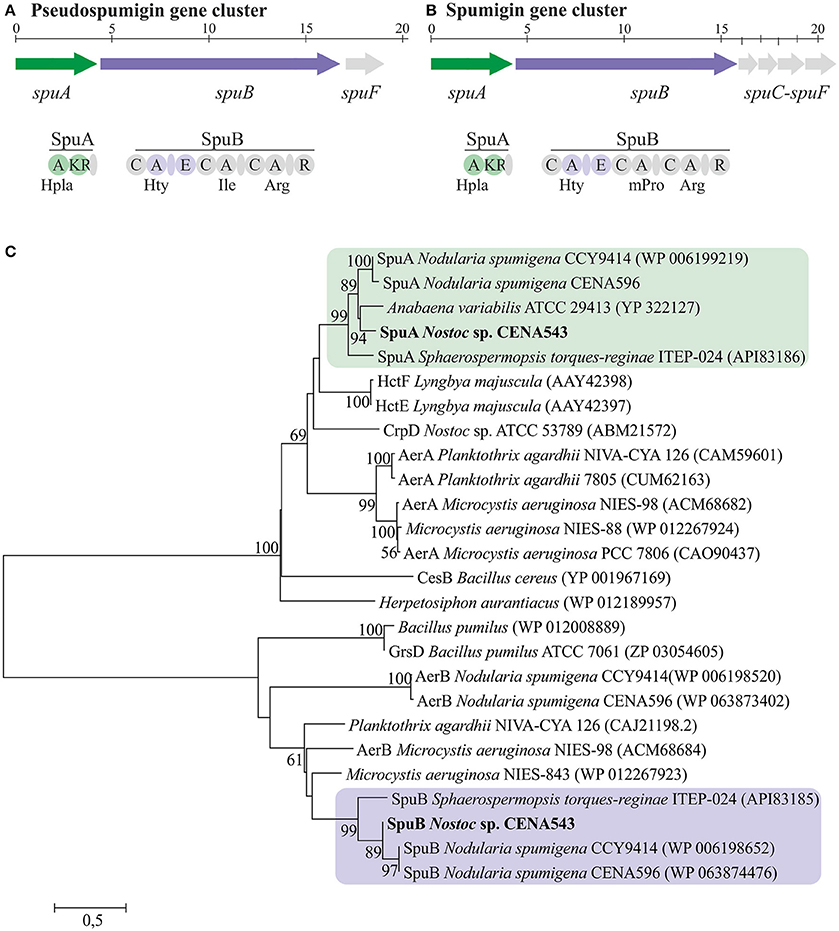
Figure 8. The evolutionary history of SpuA and SpuB of pseudospumigin. Pseudospumigin gene cluster of Nostoc sp. CENA543 (A), Spumigin gene cluster of Nodularia spumigena CCY9414 (B), and phylogenetic (neighbor-joining) tree based on SpuA and SpuB amino acid sequences (C). Bootstrap values above 50 percent of 1,000 replicates are given at the nodes. Nostoc sp. CENA543 is highlighted and region of the proteins used for the analyses are colored in green or purple (A,B).
Evolutionary History of Nostoc sp. CENA543 and Non-ribosomal Biosynthetic Genes
The phylogenetic tree that was constructed using 16S rRNA gene sequences show that Nostoc sp. CENA543 grouped with other Nostoc strains, which are more closely related to Nodularia, Anabaenopsis/Cyanospira, and Halotia strains than with other Nostoc strains (Figure 9). Nodularin producers are widely distributed in this phylogenetic tree according to their taxonomy. A phylogeny based on ndaF and mcyE genes from nodularin and microcystin biosynthetic gene clusters, respectively, indicate the multiple origin of nodularin producers (Figure 7B). Interestingly, the ndaF gene of Nostoc sp. CENA543 was closely related to mcyE genes of the strains that produce microcystins. The evolutionary history of SpuA and SpuB proteins involved in the synthesis of Psps in Nostoc sp. CENA543 shows a high similarity with the spumigin gene cluster (Figure 8C, Table S3).
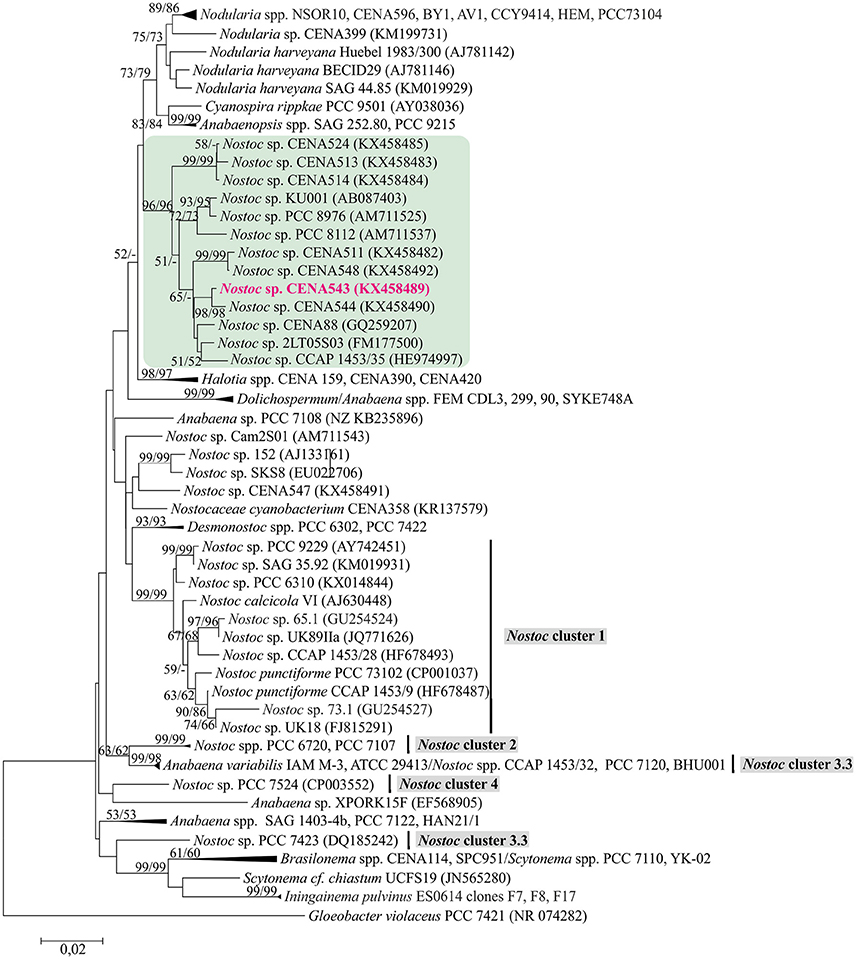
Figure 9. Phylogenetic analysis of 16S rRNA gene including Nostoc sp. CENA543. Neighbor-joining tree based on 16S rRNA gene. Bootstrap values above 50 percent from 1,000 replicates are given at the nodes (Neighbor-joining/Maximum likelihood). Nostoc sp. CENA543 is highlighted in pink.
Discussion
We studied the peptidomes of nine Brazilian Pantanal wetland extremophilic Nostoc and nine other strains of cyanobacterial genera, which have been isolated from saline-alkaline lakes. The results indicated that Nostoc spp. CENA543 and CENA544 have identical peptide profiles that diverged from the profiles of the other analyzed strains and results concerning nodularins and pseudospumigins are reported herein. Nostoc sp. CENA543 isolated from a Brazilian, shallow, saline-alkaline lake water is a free-living cyanobacterium that under laboratory conditions showed benthic growth (Figure S1). Nodularin-R (NOD-R) was the predominant NOD variant detected (Table 1). The production of NOD by the Nostoc sp. CENA543 was shown both at the genetic and the metabolic levels. NOD was originally found in the genus Nodularia, whose representatives originate mostly from brackish waters but are more rarely found in fresh water (Beattie et al., 2000; Akcaalan et al., 2009; Sivonen, 2009). Motuporin and isomotuporins are analogous compounds to nodularin, which contain Val, Ile, or 2-aminobutyric acid instead of Arg in position 2 and Adda or demethoxyAdda in position 3, have been reported in marine sponge Theonella swinhoei (de Silva et al., 1992; Figure 1). It has been commonly suggested that associated cyanobacteria are actually responsible for the production of motuporins (de Silva et al., 1992). Several cyanobacterial genera including Nostoc produce microcystins, which are close structural relatives to NODs (Figure 1). The finding of NOD synthesis by cycad and lichen symbiotic Nostoc strains was the first direct evidence of NOD production by another cyanobacterial genus than Nodularia (Gehringer et al., 2012; Kaasalainen et al., 2012). Recent studies reported that NODs were identified from a symbiotic Nostoc of liverwort Blasia pusilla L, and also from a new cyanobacterium genus and species I. pulvinus gen nov., sp. nov. that was found in a freshwater wetland spring (Liaimer et al., 2016; McGregor and Sendall, 2017). NOD-R has been previously identified from benthic cyanobacterial mats found in Lake Tikitapu, New Zealand, which contained Nostoc and lacked Nodularia strains (Wood et al., 2012). However, none of the isolated strains produced NOD-R, which in the light of what we found in the present study suggests that a non-isolated Nostoc strain might have been the NOD-R producer (Wood et al., 2012).
Quantitative UPLC-QTOF analysis showed that in low light, low salinity and high phosphorus environment without further optimization, laboratory cultivated CENA543 strain produced 4.3 mg NOD g biomass DW. This NOD production level is high and at the same level as that found in planktonic N. spumigena and benthic N. sphaerocarpa strains but more than four orders of magnitude higher compared to laboratory grown cycad endosymbiotic Nostoc strains (Table 3). In symbiosis NOD levels were 0.0025 μg g−1 in cycads and <10–60 μg g−1 in lichens. The NOD levels of lichens were thus four orders of magnitude higher than those expressed in cycads. About 7% of the lichen biomass is photobiont (Ahmadjian, 1993), which gives <0.1–0.9 mg NOD g−1 cyanobiont in lichens. Consequently, the rough estimate is that in lichens the cyanobiont, which most probably is Nostoc, will produce one tenth of the NOD levels of free living Nostoc sp. CENA543. It is, therefore, possible that lichens are much more resistant against NOD than are cycads. The resistance of lichens to hepatotoxins has already been reported earlier for microcystins (Kaasalainen et al., 2012). The mean level of NOD produced by the cyanobacterium I. pulvinus was 0.9 mg g−1 biomass DW, which is at the low level of N. spumigena NOD production but one fifth that of Nostoc sp. CENA543 NOD production (Table 3).
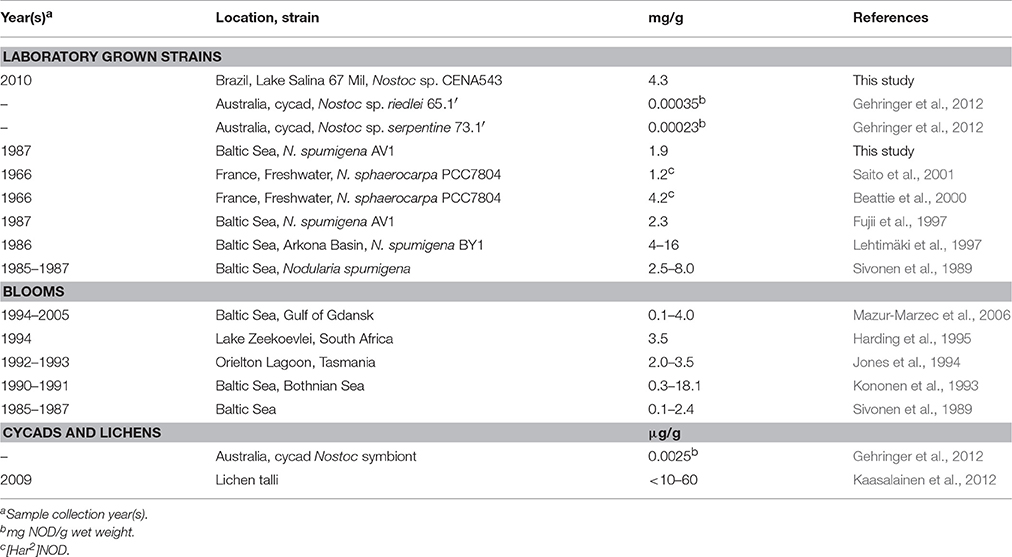
Table 3. Nodularin (NOD) concentrations (mg NOD/g dry weight) in laboratory grown Nostoc and Nodularia strains, Nodularia blooms and in cycad and lichens.
A new peptide family in the Nostoc sp. CENA543 was identified, which was named pseudospumigins (Psp; Figure 2) due to the close genetic and chemical structure resemblance to those of the spumigins. Spumigins have been found in N. spumigena, A. compacta NIES-835 and S. torques-reginae ITEP-024 strains (Fujii et al., 1997; Anas et al., 2012; Sanz et al., 2015). The spumigins structurally belong to the linear four subunit peptides, the diagnostic amino acid is Pro/mPro in position 3, whereas in the pseudospumigins this amino acid is absent and replaced by Ile, Leu, or Val. Furthermore, the genes that encode mPro in spumigin biosynthesis (Fewer et al., 2009) were absent from the pseudospumigin gene cluster. The aeruginosins are also structurally closely related to the pseudospumigins and the spumigins but the diagnostic amino acid is Choi (Figure 2). Many cyanobacterial genera produce aeruginosins but the aeruginosin family has been described in only three Nostoc strains: i.e., the terrestrial Lukešová 30/93 strain and recently found in plant leaves originating CENA352 and 458 strains (Kapuścik et al., 2013; Sanz et al., 2015). Nostosins produced by terrestrial Nostoc sp. FSN are homologous with aforementioned peptide groups but one of the middle amino acids is missing (Liu et al., 2014). In total well over 100 different structures that represent these peptide groups have been described. The Nostoc sp. CENA543 strain is not only excellent NOD producer, which is a typical feature for the Nodularia strains, but also produce pseudospumigins, which are very near structural analogs of spumigins and are almost exclusively known to be produced by N. spumigena strains. The pseudospumigin production level by the Nostoc sp. CENA543 was 0.87 mg purified from one g of biomass DW, which is roughly half that of the spumigin production level found in N. spumigena AV1 (Fewer et al., 2009) for similar purification yields. This result also links the peptidomes of Nostoc sp. CENA543 and N. spumigena strains more closely together. However, aeruginosins/pseudoaeruginosins (Figure 2) produced by the N. spumigena strains (Fewer et al., 2013; Mazur-Marzec et al., 2013; Liu et al., 2015) were not found in the Nostoc sp. CENA543 strain. Aeruginosins are estimated to be produced at significant amounts by N. spumigena AV1, 3 mg g−1 biomass DW (Fewer et al., 2013; Liu et al., 2015).
The Psp mixture (80% Psp A) was a trypsin inhibitor as are the structural analogous Sps, aeruginosins, and nostosins, which all contain a quanidino group in the Arg derivative substructure (Supporting Information in Liu et al., 2014). However, there was a wide variation in the IC50 values (μM) from 0.037 (chlorodysinosin A) to 94 (aeruginosin 126A) of these compounds without there being any clear association with the chemical structures. The inhibition by Psp A (mix) increased until the end of the 1 h measurement period as was the case for spumigin E (Figure 6). Time dependent inhibition is seen as a slow, tight and irreversible binding by inhibitors (Copeland, 2005). At the latter half of the measurement the ratio of IC50 of Psp A and Sp E stabilizes so that the Psp A (mix) was roughly 1/70th the strength of Sp E as a trypsin inhibitor and at the end of the measurement period the IC50 value was 4.5 μM. This large difference in the IC50 values was unexpected as the structures of Psp A and Sp E (Figure 2) of the subunits 1, 2, and 4 are identical. The fourth subunit of both compounds are also equal size and hydrophobically related amino acids, Ile, or Pro. The major difference between Psp A and Sp E is that the cyclic structure of the proline side chain makes Pro structurally rigid, which may in turn, affect the formation/dissociation of the enzyme-inhibitor complex. Psp A and nostosin A have identical subunits argininal and Ile but in nostosin A there is a 2-hydroxy-4-(4-hydroxyphenyl)-butanoic acid, one methylene unit longer Hpla analog and there is no fourth subunit. Nostosin A trypsin inhibition was also time dependent at the beginning of the measurement, but after 40 min IC50 stabilizes at a value of 0.35 μM (Liu et al., 2014; Figure 6). Trypsin inhibition by nostosin A was found to be 10-fold that of Psp A (mix). The commercial trypsin inhibitor, leupeptin, has the structure Ac-Leu-Leu-Argininal that resembles Psp A, Sp E and also nostosin A, and it interacts with trypsin without time dependency (Aoyagi et al., 1969; Kurinov and Harrison, 1996). The reasons for how these compounds show such different interactions with trypsin despite having lot of structural similarity remains to be investigated.
Morphological evaluations have shown that the cyanobacterial strain CENA543 belong to the genera Nostoc (Genuário et al., 2017). In a phylogenetic tree based on 16S rRNA (Figure 9), Nostoc sp. CENA543 and other Nostoc strains that were collected from the same Pantanal wetland area grouped together in a Nostoc-like cluster. Nostoc-like strains are more closely related to the Nodularia clade than to the true Nostoc clade (Herdman et al., 2015), in which the microcystin producing lichen symbiotic Nostoc sp. UK18 and UK89IIa were found (Figure 9). A Nostoc-like cluster is formed by sequences generated from free living cyanobacteria, such as the strain collected from a laundromat discharge pond in Michigan, USA (Nostoc sp. PCC 8112), or from brackish marshland on the Mediterranean coast of France (Nostoc sp. PCC 8676), or from a non-flooded rice field in Thailand (N. elgonense TH3S05), or from Brazilian mangroves (Nostoc sp. CENA175) or from a freshwater reservoir in Brazil (Nostoc sp. CENA88) (Genuário et al., 2017).
The evolutionary history based on ndaF and mcyE gene sequences indicates that different evolutionary events were responsible for the existence of nodularin producers (Figure 7B). Interestingly, ndaF of the Nostoc sp. CENA543 was more closely related to Anabaena spp. and to the microcystin producers of Nostoc sp. than to the nodularin producer Nostoc sp. 73.1 (Figure 7B). Previous studies indicate that the nodularin gene cluster has evolved after a deletion event in the microcystin gene cluster (Moffitt and Neilan, 2004; Rantala et al., 2004; Gehringer et al., 2012). Our analysis is consistent with this hypothesis. The biosynthetic gene cluster of pseudospumigin of the Nostoc sp. CENA543 is very similar to previously described spumigin gene cluster (Fewer et al., 2009; Figure 8). The phylogenetic analysis that used SpuA and SpuB amino acid sequences also indicated the closer relation of the pseudospumigin and spumigin sequences (Figure 8C).
Author Contributions
JJ, TKS, MFF, and KS designed the study. LMPH, TKS, MW, EH, and PP performed the experiments. JJ, TKS, DPF, and HW analyzed and interpreted the data. JJ, TKS, DPF, and KS wrote themanuscript, which was corrected, revised and approved by all authors.
Funding
This work was supported by grants awarded to KS from the Academy of Finland (1273798). The São Paulo Research Foundation (FAPESP, 2013/50425-8) awarded a grant to MFF.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Lyudmila Saari for purifying and maintaining the collected cyanobacteria strains and group of MFF for original isolation of the strains.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.01963/full#supplementary-material
References
Akcaalan, R., Mazur-Marzec, H., Zalewska, A., and Albay, M. (2009). Phenotypic and toxicological characterization of toxic Nodularia spumigena from a freshwater lake in Turkey. Harmful Algae 8, 273–278. doi: 10.1016/j.hal.2008.06.007
Anas, A. R. J., Kisugi, T., Umezawa, T., Matsuda, F., Campitelli, M. R., Quinn, R. J., et al. (2012). Thrombin inhibitors from the freshwater cyanobacterium Anabaena compacta. J. Nat. Prod. 75, 1546–1552. doi: 10.1021/np300282a
Andreote, A. P. D., Vaz, M. G. M. V., Genuario, D. B., Barbiero, L., Rezende-Filho, A. T., and Fiore, M. F. (2014). Non-hetero0cytous cyanobacteria from Brazilian saline-alkaline lakes. J. Phycol. 50, 675–684. doi: 10.1111/jpy.12192
Aoyagi, T., Takeuchi, T., Matsuzak, A., Kawamura, K., Kondo, S., Hamada, M., et al. (1969). Leupeptins, new protease inhibitors from actinomycetes. J. Antibiot. 22, 283–286. doi: 10.7164/antibiotics.22.283
Bachmann, B. O., and Ravel, J. (2009). In silico prediction of microbial secondary metabolic pathways from DNA sequence data. Method Enzymol. 458, 181–217. doi: 10.1016/S0076-6879(09)04808-3
Beattie, K. A., Kaya, K., and Codd, G. A. (2000). The cyanobacterium Nodularia PCC7804, of freshwater origin, produces [L-Har2]nodularin. Phytochemistry 54, 57–61. doi: 10.1016/S0031-9422(00)00045-5
Blin, K., Medema, M. H., Kottmann, R., Lee, S. Y., and Weber, T. (2017). The antiSMASH database, a comprehensive database of microbial secondary metabolitebiosynthetic gene clusters. Nucleic Acids Res. 45, D555–D559. doi: 10.1093/nar/gkw960
Copeland, R. A. (2005). Evaluation of enzyme inhibitors in drug discovery: a guide for medicinal chemists and pharmacologists. Methods Biochem. Anal. 46, 1–265. doi: 10.1002/9781118540398
de Silva, E. D., Williams, D. E., Andersen, R. J., Klix, H., Holmes, C. F. B., and Allen, T. M. (1992). Motuporin, a potent protein phosphatase inhibitor isolated from the Papua New Guinea sponge Theonella swinhoei Gray. Tetrahedron Lett. 33, 1561–1564. doi: 10.1016/S0040-4039(00)91674-5
Dittmann, E., Gugger, M., Sivonen, K., and Fewer, D. P. (2015). Natural product biosynthetic diversity and comparative genomics of the cyanobacteria. Trends Microbiol. 23, 642–652. doi: 10.1016/j.tim.2015.07.008
Fewer, D. P., Jokela, J., Paukku, E., Österholm, J., Wahlsten, M., Permi, P., et al. (2013). New structural variants of aeruginosin produced by the toxic bloom forming cyanobacterium Nodularia spumigena. PLoS ONE 8:e73618. doi: 10.1371/journal.pone.0073618
Fewer, D. P., Jokela, J., Rouhiainen, L., Wahlsten, M., Koskenniemi, K., Stal, L., et al. (2009). The non-ribosomal assembly and frequent occurrence of the protease inhibitors spumigins in the bloom-forming cyanobacterium Nodularia spumigena. Mol. Microbiol. 73, 924–937. doi: 10.1111/j.1365-2958.2009.06816.x
Fujii, K., Sivonen, K., Adachi, K., Noguchi, K., Sano, H., Hirayama, K., et al. (1997). Comparative study of toxic and non-toxic cyanobacterial products: novel peptides from toxic Nodularia spumigena AV1. Tetrahedron Lett. 38, 5525–5528. doi: 10.1016/S0040-4039(97)01192-1
Gehringer, M. M., Adler, L., Roberts, A. A., Moffitt, M. C., Mihali, T. K., Mills, T. J. T., et al. (2012). Nodularin, a cyanobacterial toxin, is synthesized in planta by symbiotic Nostoc sp. ISME J. 6, 1834–1847. doi: 10.1038/ismej.2012.25
Genuário, D. B., Andreote, A. P. D., Vaz, M. G. M. V., and Fiore, M. F. (2017). Heterocyte-forming cyanobacteria from Brazilian saline-alkaline lakes. Mol. Phylogenet. Evol. 109, 105–112. doi: 10.1016/j.ympev.2016.12.032
Genuário, D. B., Silva-Stenico, M. E., Welker, M., Moraes, L. A. B., and Fiore, M. F. (2010). Characterization of a microcystin and detection of microcystin synthetase genes from a Brazilian isolate of Nostoc. Toxicon 55, 846–854. doi: 10.1016/j.toxicon.2009.12.001
Harding, W. R., Rowe, N., Wessels, J. C., Beattie, K. A., and Codd, G. A. (1995). Death of a dog attributed to the cyanobacterial (blue-green algal) hepatotoxin nodularin in South Africa. J. S. Afr. Vet. Ass. 66, 256–259.
Herdman, M., Castenholz, R. W., and Rippka, R. (2015). “Form-Nostoc,” in Bergey's Manual of Systematics of Archaea and Bacteria, ed W. B. Whitman (New York, NY: John Wiley & Sons, Inc.), 1–10.
Jones, G. J., Blackburn, S. I., and Parker, N. S. (1994). A toxic bloom of Nodularia spumigena Mertens in Orielton Lagoon Tasmania. Austr. J. Mar. Freshw. Res. 45, 787–800. doi: 10.1071/MF9940787
Kaasalainen, U., Fewer, D. P., Jokela, J., Wahlsten, M., Sivonen, K., and Rikkinen, J. (2012). Lichens as a nursery for evolution—cyanobacteria produce a high variety of hepatotoxic peptides in symbiosis. Proc. Natl. Acad. Sci. U.S.A. 109, 5886–5891. doi: 10.1073/pnas.1200279109
Kapuścik, A., Hrouzek, P., Kuzma, M., Bártová, S., Novák, P., Jokela, J., et al. (2013). A novel aeruginosin-865 from Nostoc sp. as a potent anti-inflammatory agent. ChemBioChem 14, 2329–2337. doi: 10.1002/cbic.201300246
Kobayashi, A., and Kajiyama, S. I. (1998). Secondary metabolites of cyanobacteria Nostoc sp. Chin. J. Oceanol. Limnol. 16, 109–117. doi: 10.1007/BF02849088
Kolmonen, E., Sivonen, K., Rapala, J., and Haukka, K. (2004). Diversity of cyanobacteria and heterotrophic bacteria in cyanobacterial blooms in Lake Joutikas, Finland. Aquat. Microbiol. Ecol. 36, 201–211. doi: 10.3354/ame036201
Kononen, K., Sivonen, K., and Lehtimäki, J. (1993). “Toxicity of Phytoplankton Blooms in the Gulf of Finland and Gulf of Bothnia, Baltic Sea,” in Toxic Phytoplankton Blooms in the Sea, Proceedings of International Conference on Toxic Marine Phytoplankton, eds T. J. Smayda and Y. Shimizu (Amsterdam: Elsevier Science Publishers), 269–273.
Kotai, J. (1972). Instructions for Preparation of Modified Nutrient Solution Z8 for Algae. Norwegian Institute for Water Research, Oslo.
Kurinov, I. V., and Harrison, R. W. (1996). Two crystal structures of the leupeptin-trypsin complex. Protein Sci. 5, 752–758. doi: 10.1002/pro.5560050420
Lehtimäki, J., Moisander, P., Sivonen, K., and Kononen, K. (1997). Growth, nitrogen fixation, and nodularin production by two Baltic Sea cyanobacteria. Appl. Environ. Microbiol. 63, 1647–1656.
Liaimer, A., Jensen, J. B., and Dittmann, E. (2016). A genetic and chemical perspective on symbiotic recruitment of cyanobacteria of the genus Nostoc into the host plant Blasia pusilla L. Front. Microbiol. 7:1693. doi: 10.3389/fmicb.2016.01693
Liu, L., Budnjo, A., Jokela, J., Haug, B. E., Fossen, T., Fewer, D. P., et al. (2015). Pseudoaeruginosins, nonribosomal peptides in Nodularia spumigena. ACS Chem. Biol. 10, 725–733. doi: 10.1021/cb5004306
Liu, L., Jokela, J., Wahlsten, M., Nowruzi, B., Permi, P., Zhang, Y. Z., et al. (2014). Nostosins, trypsin inhibitors isolated from the terrestrial cyanobacterium Nostoc sp. strain FSN. J. Nat. Prod. 77, 1784–1790. doi: 10.1021/np500106w
Mazur-Marzec, H., Kaczkowska, M. J., Agata Blaszczyk, A., Akcaalan, R., Spoof, L., and Meriluoto, J. (2013). Diversity of peptides produced by Nodularia spumigena from various geographical regions. Mar. Drugs 11, 1–19. doi: 10.3390/md11010001
Mazur-Marzec, H., KreZel, A., Kobos, J., and Plinski, M. (2006). Toxic Nodularia spumigena blooms in the coastal waters of the Gulf of Gdansk: a ten-year survey. Oceanologia 48, 255–273.
McGregor, G. B., and Sendall, B. C. (2017). Iningainema pulvinus gen. nov., sp. nov. (Cyanobacteria, Scytonemataceae) a new nodularin producer from Edgbaston Reserve, north-eastern Australia. Harmful Algae 62, 10–19. doi: 10.1016/j.hal.2016.11.021
Meriluoto, J., Spoof, L., and Codd, G. (eds.). (2017). Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis. Chichester: John Wiley & Sons, Ltd.
Moffitt, M. C., and Neilan, B. A. (2004). Characterization of the nodularin synthetase gene cluster and proposed theory of the evolution of cyanobacterial hepatotoxins. Appl. Environ. Microbiol. 70, 6353–6362. doi: 10.1128/AEM.70.11.6353-6362.2004
Oksanen, I., Jokela, J., Fewer, D. P., Wahlsten, M., Rikkinen, J., and Sivonen, K. (2004). Discovery of rare and highly toxic microcystins from lichen-associated cyanobacterium Nostoc sp. strain IO-102-I. Appl. Environ. Microbiol. 70, 5756–5763. doi: 10.1128/AEM.70.10.5756-5763.2004
Rantala, A., Fewer, D. P., Hisbergues, M., Rouhiainen, L., Vaitomaa, J., Börner, T., et al. (2004). Phylogenetic evidence for the early evolution of microcystin synthesis. Proc. Natl. Acad. Sci. U.S.A. 101, 568–573. doi: 10.1073/pnas.0304489101
Rausch, C., Weber, T., Kohlbacher, O., Wohlleben, W., and Huson, D. H. (2005). Specificity prediction of adenylation domains in nonribosomal peptide synthetases (NRPS) using transductive support vector machines (TSVMs). Nucleic Acids Res. 33, 5799–5808. doi: 10.1093/nar/gki885
Repka, S., Mehtonen, J., Vaitomaa, J., Saari, L., and Sivonen, K. (2001). Effect of nutrients on growth and nodularin production of Nodularia strain GR8b. Microbial. Ecol. 42, 606–613. doi: 10.1007/s00248-001-0026-8
Röttig, M., Medema, M. H., Blin, K., Weber, T., Rausch, C., and Kohlbacher, O. (2011). NRPSpredictor2—a web server for predicting NRPS adenylation domain specificity. Nucleic Acids Res. 39, W362–W367. doi: 10.1093/nar/gkr323
Rutherford, K., Parkhill, J., Crook, J., Horsnell, T., Rice, P., Rajandream, M. A., et al. (2000). Artemis: sequence visualization and annotation. Bioinformatics 16, 944–945. doi: 10.1093/bioinformatics/16.10.944
Saito, K., Konno, A., Ishii, H., Saito, H., Nishida, F., Abe, T., et al. (2001). Nodularin-Har: a new nodularin from Nodularia. J. Nat. Prod. 64, 139–141. doi: 10.1021/np000299z
Sanz, M., Dörr, F. A., and Pinto, E. (2015). First report of spumigin production by the toxic Sphaerospermopsis torques-reginae cyanobacterium. Toxicon 108, 15–18. doi: 10.1016/j.toxicon.2015.09.019
Simola, O., Wiberg, M., Jokela, J., Wahlsten, M., Sivonen, K., and Syrjä, P. (2012). Pathological findings and toxin identification in cyanobacterial (Nodularia spumigena) intoxication in a dog. Vet. Pathol. 49, 755–759. doi: 10.1177/0300985811415703
Sivonen, K. (2009). “Cyanobacterial toxins,” in Encyclopedia of Microbiology, 3rd Edn, ed. M. Schaechter (Oxford: Elsevier), 290–307.
Sivonen, K., Carmichael, W. W., Namikoshi, M., Rinehart, K. L., Dahlem, A. M., and Niemelä, S. I. (1990). Isolation and characterization of hepatotoxic microcystin homologs from the filamentous freshwater cyanobacterium Nostoc sp. strain 152. Appl. Environ. Microbiol. 56, 2650–2657.
Sivonen, K., Kononen, K., Carmichael, W. W., Dahlem, A. M., Rinehart, K. L., Kiviranta, J., et al. (1989). Occurrence of the hepatotoxic cyanobacterium Noduaria spumigena in the Baltic Sea and structure of the toxin. Appl. Environ. Microbiol. 55, 1990–1995.
Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S. (2013). MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729. doi: 10.1093/molbev/mst197
Voß, B., Bolhuis, H., Fewer, D. P., Kopf, M., Möke, F., Haas, F., et al. (2013). Insights into the physiology and ecology of the brackish-water-adapted cyanobacterium Nodularia spumigena CCY9414 based on a genome-transcriptome analysis. PLoS ONE 8:e60224. doi: 10.1371/journal.pone.0060224
Keywords: cyanobacteria, Nostoc, nodularin, spumigin, biosynthesis
Citation: Jokela J, Heinilä LMP, Shishido TK, Wahlsten M, Fewer DP, Fiore MF, Wang H, Haapaniemi E, Permi P and Sivonen K (2017) Production of High Amounts of Hepatotoxin Nodularin and New Protease Inhibitors Pseudospumigins by the Brazilian Benthic Nostoc sp. CENA543. Front. Microbiol. 8:1963. doi: 10.3389/fmicb.2017.01963
Received: 07 June 2017; Accepted: 25 September 2017;
Published: 09 October 2017.
Edited by:
Petra M. Visser, University of Amsterdam, NetherlandsReviewed by:
Antonio Quesada, Universidad Autonoma de Madrid, SpainReyhan Akcaalan, Istanbul University, Turkey
Copyright © 2017 Jokela, Heinilä, Shishido, Wahlsten, Fewer, Fiore, Wang, Haapaniemi, Permi and Sivonen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kaarina Sivonen, kaarina.sivonen@helsinki.fi