Characterisation of Campylobacter spp. Isolated from Poultry in KwaZulu-Natal, South Africa

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Prevalence of Campylobacter spp.
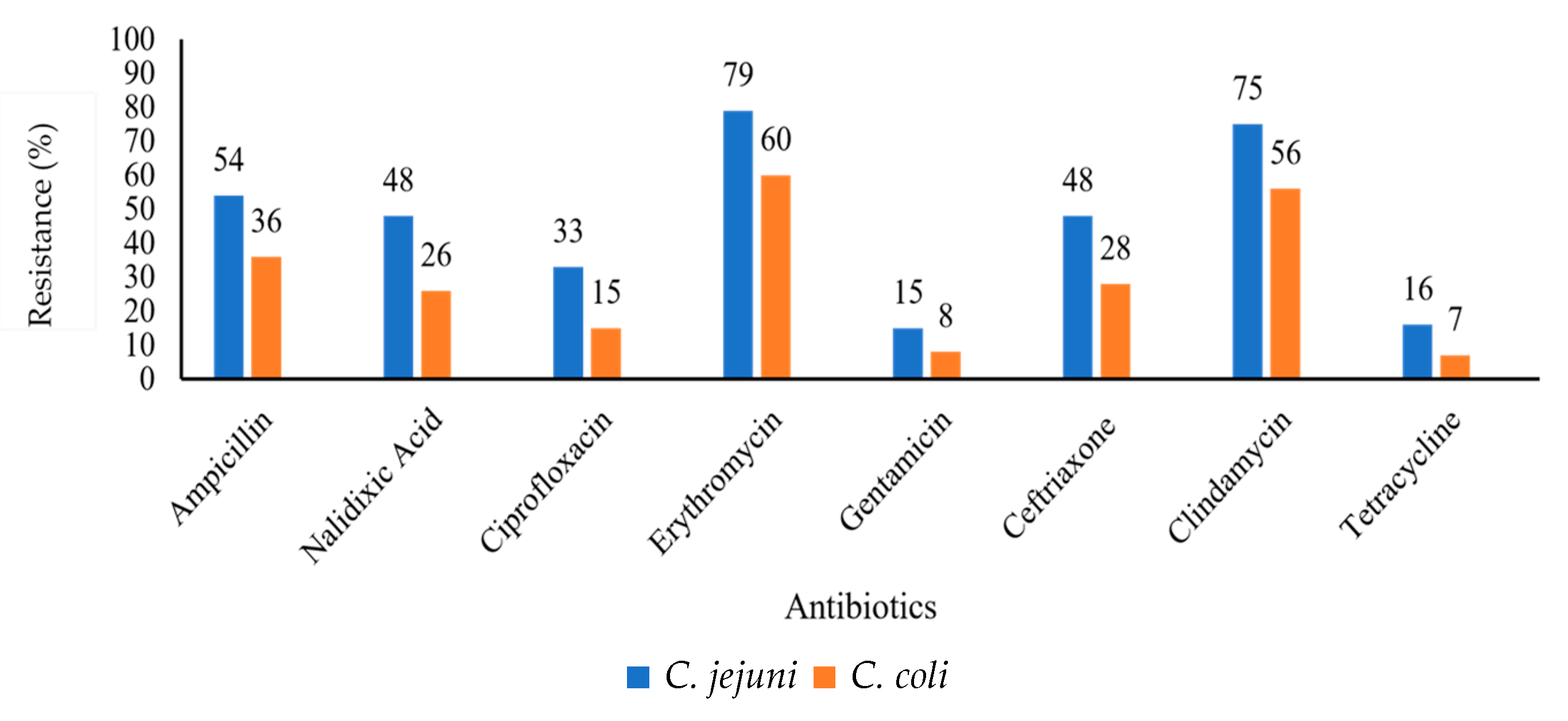
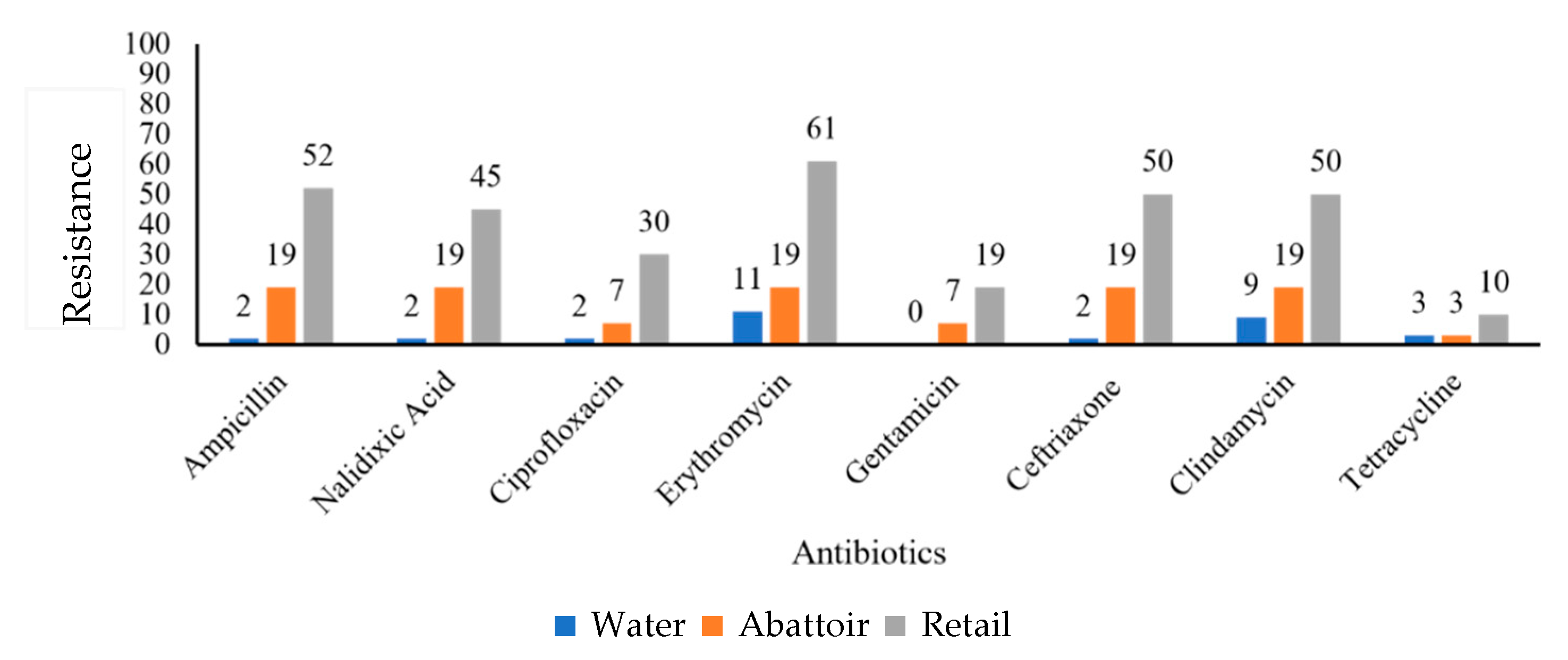
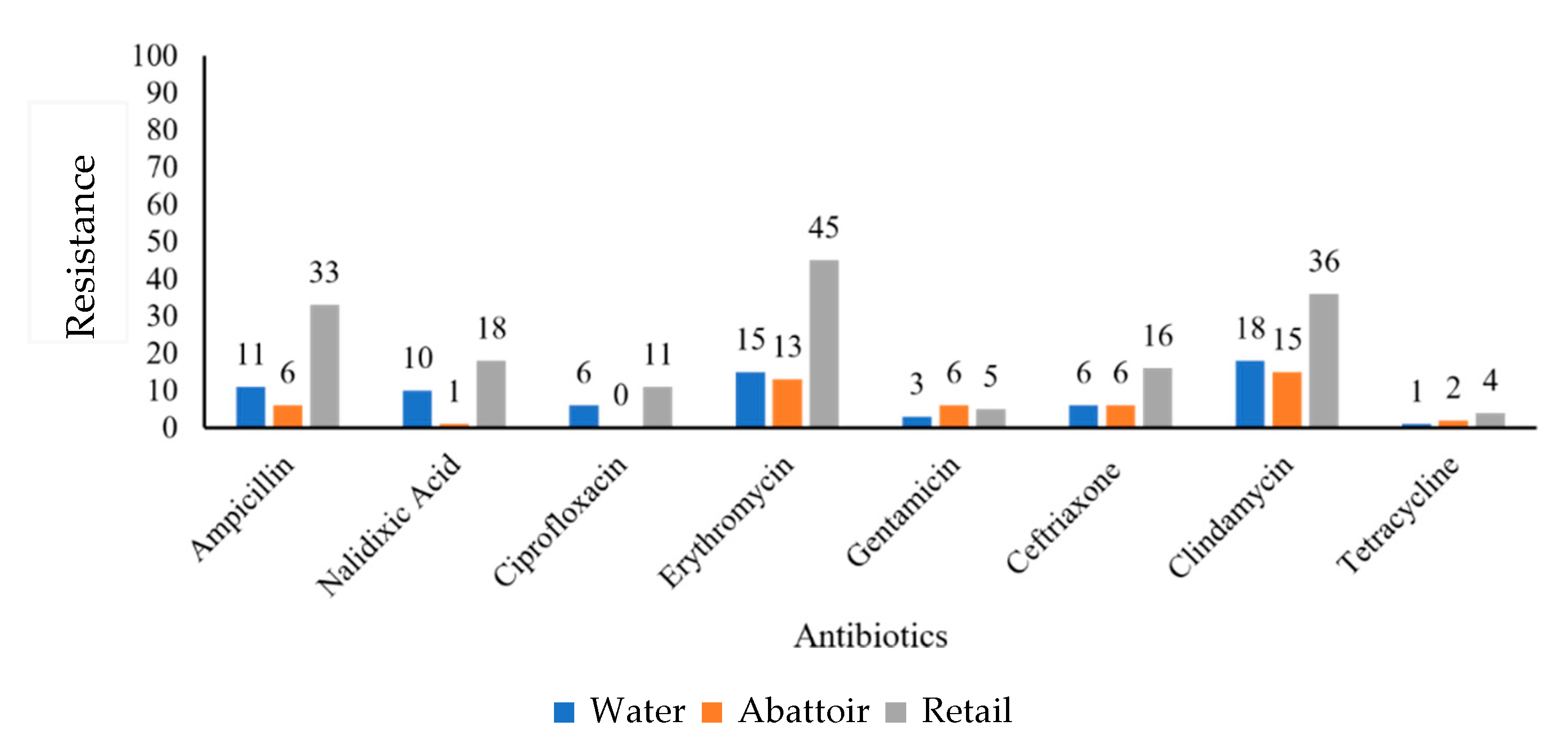
2.2. Antibiotic Susceptibility
2.3. Antibiotic Resistance Genes
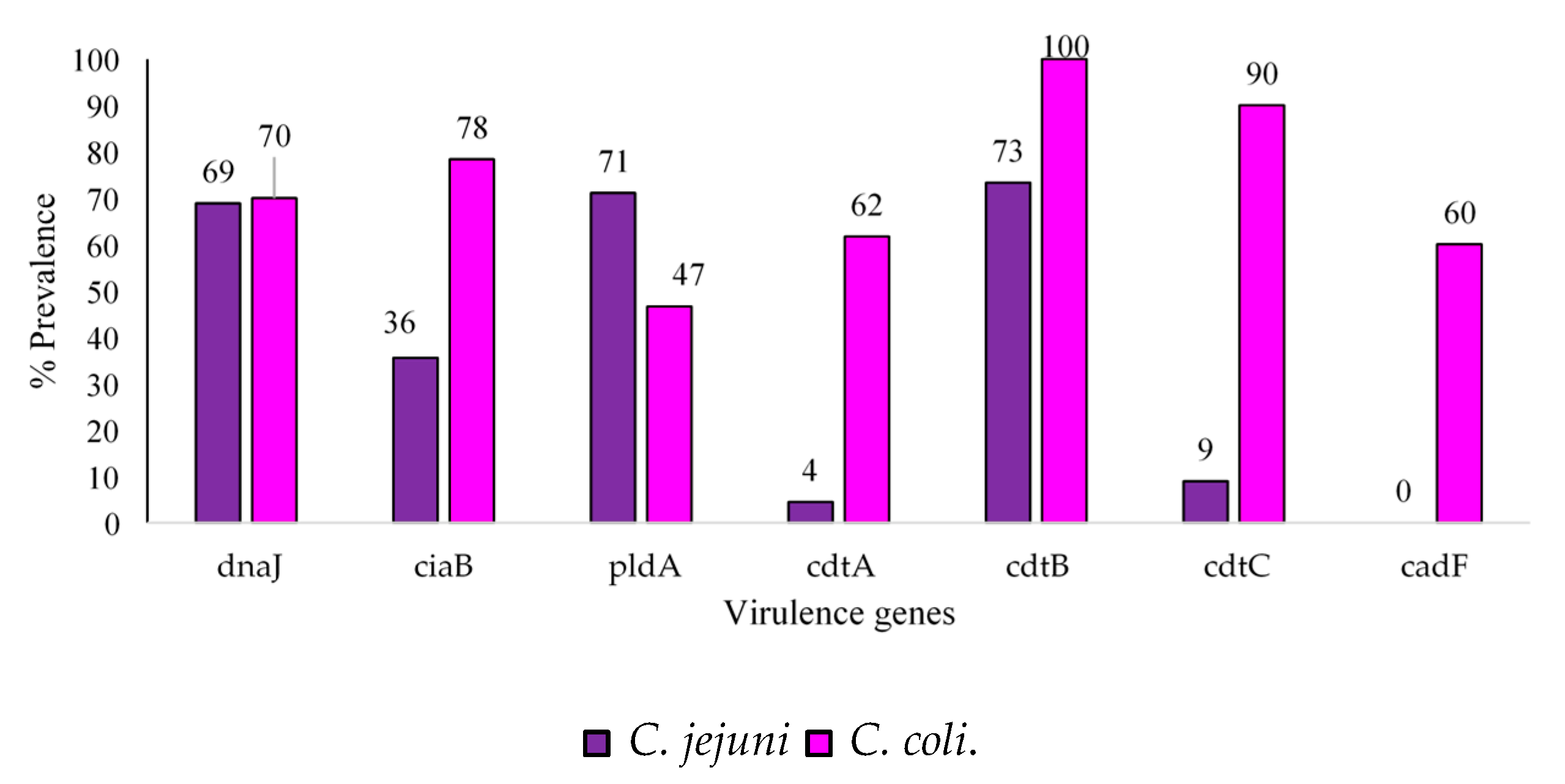
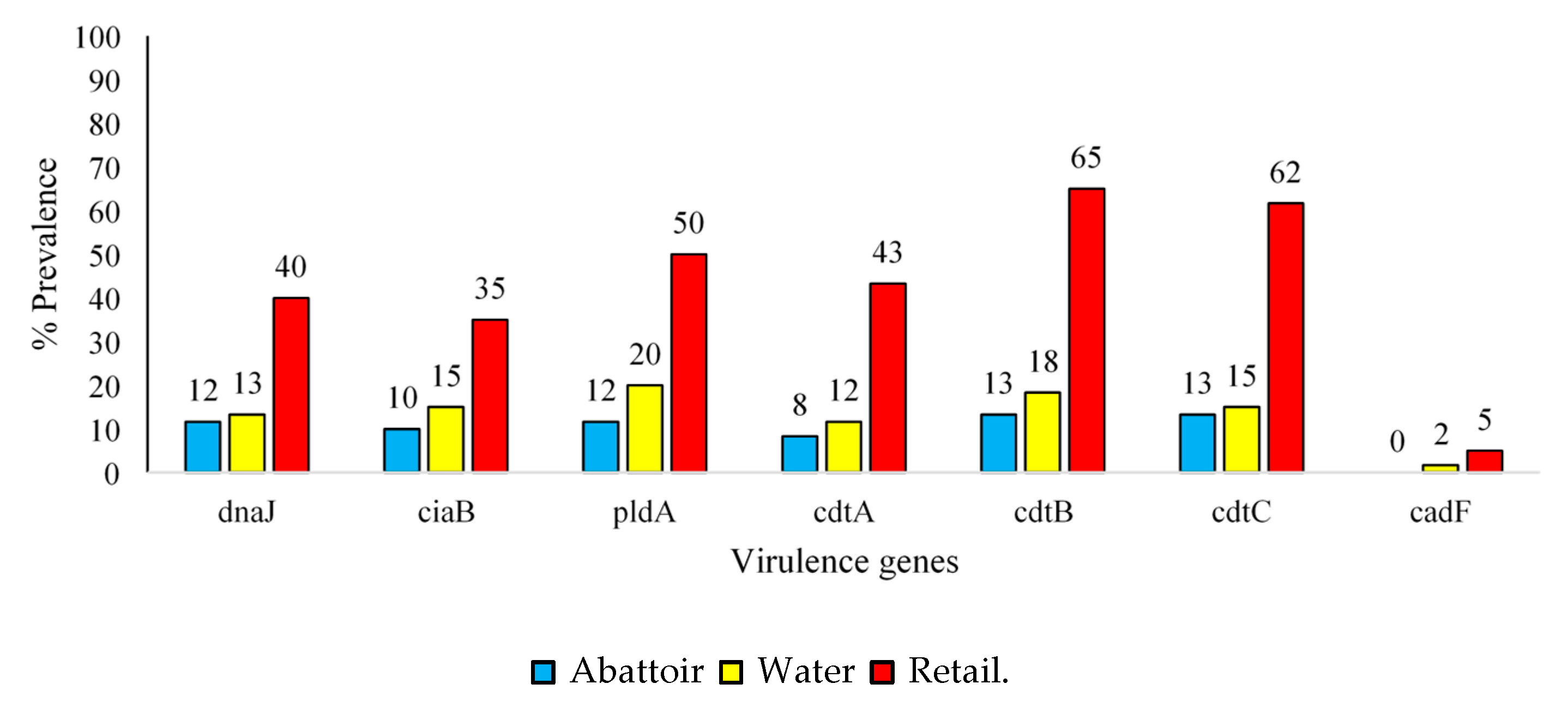
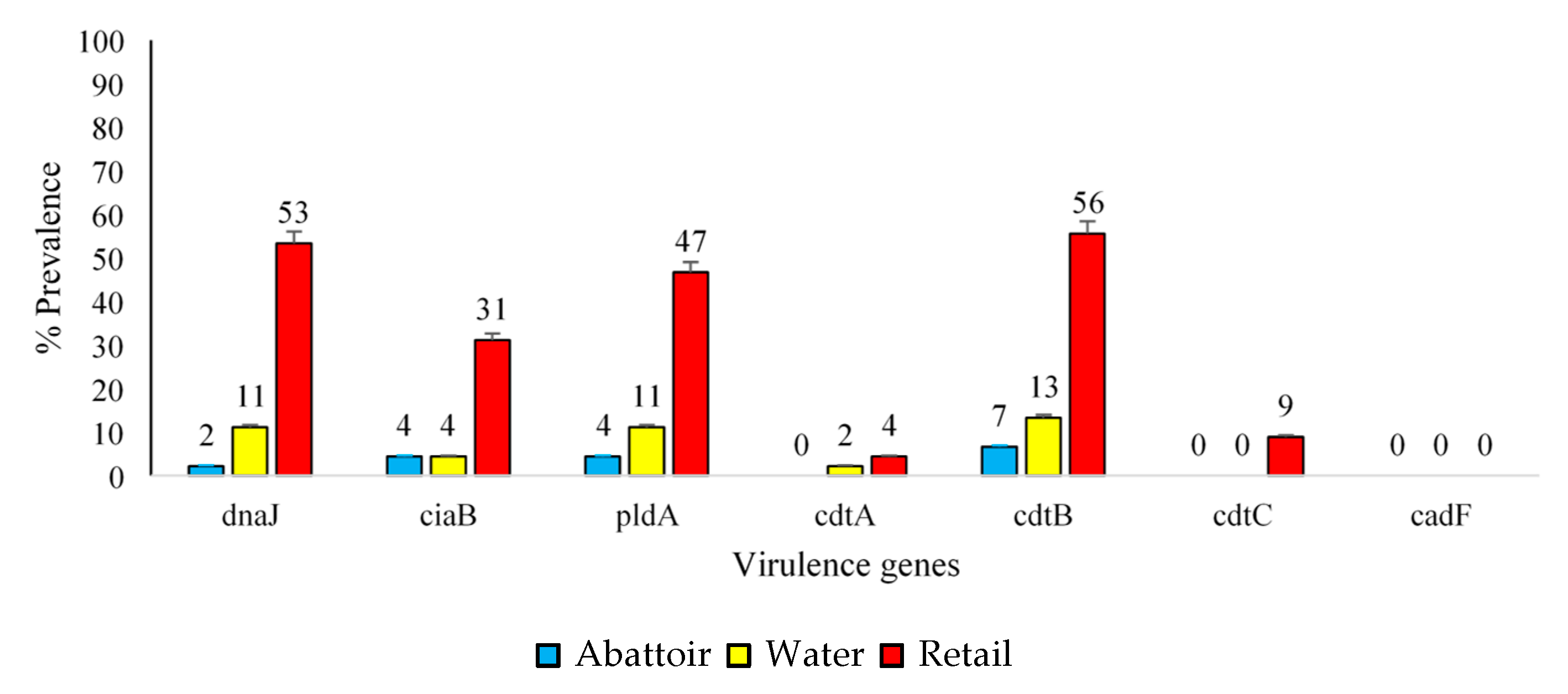
2.4. Virulence Factors
2.5. Genetic Relationship by Pulsed-Field Gel Electrophoresis
3. Discussion
3.1. Prevalence of Campylobacter spp.
3.2. Virulence Factors
3.3. Antibiotic Resistance Profiles and Resistance Determinants
3.4. Genetic Relatedness of Isolates
4. Materials and Methods
4.1. Ethical Approval
4.2. Sampling Procedure
4.3. Isolation and Identification of Campylobacter spp.
4.4. DNA Extraction
4.5. Molecular Confirmation Of Isolates
4.6. Antibiotic Susceptibility Testing of Isolates
4.7. Molecular Detection and DNA Sequence Analysis of Genetic Determinants of Resistance
4.8. Detection of Virulence Genes
4.9. Genetic Relationship by Pulsed-Field Gel Electrophoresis (PFGE)
4.10. Data Analysis and Interpretation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Umaraw, P.; Prajapati, A.; Verma, A.K.; Pathak, V.; Singh, V.P.; Umaraw, P.; Prajapati, A.; Verma, A.K.; Pathak, V. Control of campylobacter in poultry industry from farm to poultry processing unit: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Alanis, A.J. Resistance to antibiotics: Are we in the post-antibiotic era? Arch. Med. Res. 2005, 36, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Butzler, J.-P. Campylobacter, from obscurity to celebrity. Clin. Microbiol. Infect. 2004, 10, 868–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO) Campylobacter. Available online: https://www.who.int/news-room/fact-sheets/detail/campylobacter (accessed on 30 June 2019).
- Reddy, S.; Zishiri, O.T. Detection and prevalence of antimicrobial resistance genes in Campylobacter spp. isolated from chickens and humans. Onderstepoort J. Veter. Res. 2017, 84, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [Green Version]
- Sibanda, N.; McKenna, A.; Richmond, A.; Ricke, S.C.; Callaway, T.; Stratakos, A.C.; Gundogdu, O.; Corcionivoschi, N. A Review of the Effect of Management Practices on Campylobacter Prevalence in Poultry Farms. Front. Microbiol. 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Hermans, D.; Van Deun, K.; Martel, A.; Van Immerseel, F.; Messens, W.; Heyndrickx, M.; Haesebrouck, F.; Pasmans, F. Colonization factors of Campylobacter jejuni in the chicken gut. Veter. Res. 2011, 42, 82. [Google Scholar] [CrossRef] [Green Version]
- Wegener, H.C. Antibiotics in animal feed and their role in resistance development. Curr. Opin. Microbiol. 2003, 6, 439–445. [Google Scholar] [CrossRef]
- Maćkiw, E.; Korsak, D.; Rzewuska, K.; Tomczuk, K.; Rożynek, E. Antibiotic resistance in Campylobacter jejuni and Campylobacter coli isolated from food in Poland. Food Control 2012, 23, 297–301. [Google Scholar] [CrossRef]
- Bolton, D.J. Campylobacter virulence and survival factors. Food Microbiol. 2015, 48, 99–108. [Google Scholar] [CrossRef]
- Semenya, M. Poultry Industry: South African Poultry Association Briefing. In DAFF 2018/19 Annual Performance Plan; Department of Agriculture, Forestry and Fisheries: Pretoria, South Africa, 2018. [Google Scholar]
- Viljoen, W. The South African Poultry Sector—Trade, Consumption, Production and Inputs; Tralac Trade Law Centre: Stellenbosch, South Africa, 2017. [Google Scholar]
- WHO. Integrated Surveillance of Antimicrobial Resistance in Foodborne Bacteria: Application of a One Health Approach: Guidance from the WHO Advisory Group on Integrated Surveillance of Antimicrobial Resistance (AGISAR); WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Amoako, D.G.; Somboro, A.M.; Abia, A.L.K.; Allam, M.; Ismail, A.; Bester, L.; Essack, S.Y. Genomic analysis of methicillin-resistant Staphylococcus aureus isolated from poultry and occupational farm workers in Umgungundlovu District, South Africa. Sci. Total Environ. 2019, 670, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Molechan, C.; Amoako, D.G.; Abia, A.L.K.; Somboro, A.M.; Bester, L.A.; Essack, S.Y. Molecular epidemiology of antibiotic-resistant Enterococcus spp. from the farm-to-fork continuum in intensive poultry production in KwaZulu-Natal, South Africa. Sci. Total Environ. 2019, 692, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Newell, D.G.; Fearnley, C. Sources of Campylobacter Colonization in Broiler Chickens. Appl. Environ. Microbiol. 2003, 69, 4343–4351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, W.L.; Murray, C. Campylobacter jejuni Seasonal Recovery Observations of Retail Market Broilers 1. Poult. Sci. 1996, 76, 314–317. [Google Scholar] [CrossRef] [PubMed]
- Powell, L.; Lawes, J.; Clifton-Hadley, F.; Rodgers, J.; Harris, K.; Evans, S.; Vidal, A. The prevalence of Campylobacter spp. in broiler flocks and on broiler carcases, and the risks associated with highly contaminated carcases. Epidemiol. Infect. 2019, 140, 2233–2246. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, K.; Osek, J. Identification of Virulence Genes in Campylobacter Jejuni and C. Coli Isolates by PCR. Bull. Vet. Inst. Pulawy 2008, 52, 211–216. [Google Scholar]
- Rizal, A.; Kumar, A.; Vidyarthi, A.S. Prevalence of Pathogenic Genes in Campylobacter jejuni Isolated from Poultry and Human. Internet J. Food Saf. 2010, 12, 29–34. [Google Scholar]
- Carvalho, A.F.D.; Martins, D.; Azevedo, S.S.; Piatti, R.M.; Genovez, M.E.; Scarcelli, E. Detection of CDT toxin genes in Campylobacter spp. strains isolated from broiler carcasses and vegetables in São Paulo, Brazil. Braz. J. Microbiol. 2013, 699, 693–699. [Google Scholar] [CrossRef] [Green Version]
- Lapierre, L.; Gatica, M.A.; Riquelme, V.; Vergara, C.; Yañez, J.M.; San Martín, B.; Sáenz, L.; Vidal, M.; Martínez, M.C.; Araya, P.; et al. Characterization of Antimicrobial Susceptibility and Its Association with Virulence Genes Related to Adherence, Invasion, and Cytotoxicity in Campylobacter jejuni and Campylobacter coli Isolates from Animals, Meat, and Humans. Microb. Drug Resist. 2016, 22, 432–444. [Google Scholar] [CrossRef]
- Wieczorek, K. Antimicrobial resistance and virulence markers of campylobacter jejuni and campylobacter coli isolated from retail poultry meat in Poland. Bull. Vet. Inst. Pulawy 2010, 54, 563–569. [Google Scholar]
- Koolman, L.; Whyte, P.; Burgess, C.; Bolton, D. International Journal of Food Microbiology Virulence gene expression, adhesion and invasion of Campylobacter jejuni exposed to oxidative stress (H2O2). Int. J. Food Microbiol. 2016, 220, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Wareing, D.R.A.; Kramer, J.M.; Frost, J.A.; Bolton, F.J. Campylobacter Contamination of Raw Meat and Poultry at Retail Sale: Identification of Multiple Types and Comparison with Isolates from Human Infection. J. Food Prot. 2000, 63, 1654–1659. [Google Scholar]
- Melo, R.T.; Nalevaiko, P.C.; Mendonça, E.P.; Borges, L.W.; Fonseca, B.B.; Beletti, M.E.; Rossi, D.A. Campylobacter jejuni strains isolated from chicken meat harbour several virulence factors and represent a potential risk to humans. Food Control 2013, 33, 227–231. [Google Scholar] [CrossRef] [Green Version]
- Di Sario, G. Investigation of the Effects of Campylobacter iejuni Virulence Factors in Human Cells: Different Pathways Involved. Ph.D. Thesis, Università Degli Studi di Urbino Carlo Bo, Urbino, Italy, 2017. [Google Scholar]
- Baserisalehi, M.; Bahador, N. Anaerobe Chemotactic behavior of Campylobacter spp. in function of different temperatures. Anaerobe 2011, 17, 459–462. [Google Scholar] [CrossRef]
- Shobo, C.O.; Bester, L.A.; Baijnath, S.; Somboro, A.M.; Peer, A.K.; Essack, S.Y. Antibiotic resistance profiles of Campylobacter species in the South Africa private health care sector. J. Infect. Dev. Ctries. 2016, 10, 1214–1221. [Google Scholar] [CrossRef] [Green Version]
- Denis, M.; Rose, V.; Balaine, L.; Salvat, G. Diversity of Pulsed-Field Gel Electrophoresis Profiles of Campylobacter jejuni and Campylobacter coli from Broiler Chickens in France. Poult. Sci. 2007, 87, 1662–1671. [Google Scholar] [CrossRef]
- Bester, L.A.; Essack, S.Y.; Essack, S. Prevalence of antibiotic resistance in Campylobacter isolates from commercial poultry suppliers in KwaZulu-Natal, South Africa. J. Antimicrob. Chemother. 2008, 62, 1298–1300. [Google Scholar] [CrossRef] [Green Version]
- Gibreel, A.; Tracz, D.M.; Nonaka, L.; Ngo, T.M.; Connell, S.R.; Taylor, D.E. Incidence of Antibiotic Resistance in Campylobacter jejuni Isolated in Alberta, Canada, from 1999 to 2002, with Special Reference to tet(O)-Mediated Tetracycline Resistance. Antimicrob. Agents Chemother. 2004, 48, 3442–3450. [Google Scholar] [CrossRef] [Green Version]
- Gardner, S.P.; Kendall, K.J.; Taveirne, M.E.; Olson, J.W. Complete Genome Sequence of Campylobacter jejuni subsp. jejuni ATCC 35925. Genome Announc. 2017, 5, 30–31. [Google Scholar] [CrossRef] [Green Version]
- Linton, D.; Lawson, A.J.; Owen, R.J.; Stanley, J. PCR detection, identification to species level, and fingerprinting of Campylobacter jejuni and Campylobacter coli direct from diarrheic samples. J. Clin. Microbiol. 1997, 35, 2568–2572. [Google Scholar] [CrossRef] [Green Version]
- Amri, A.A.; Senok, A.C.; Ismaeel, A.Y.; Al-mahmeed, A.E.; Botta, G.A. Multiplex PCR for direct identification of Campylobacter spp. in human and chicken stools. J. Med. Microbiol. 2019, 56, 1350–1355. [Google Scholar] [CrossRef] [PubMed]
- Chukwu, M.O.; Abia, A.L.K.; Ubomba-Jaswa, E.; Obi, L.; Dewar, J.B. Characterization and Phylogenetic Analysis of Campylobacter Species Isolated from Paediatric Stool and Water Samples in the Northwest Province, South Africa. Int. J. Environ. Res. Public Health 2019, 16, 2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing: 27th Edition Informational Supplement M100-S27; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Huq, M.; Gonis, G.; Istivan, T. Development and Evaluation of a Multiplex PCR for the Detection of Campylobacter concisus and Other Campylobacter spp. from Gastroenteritis Cases. Open J. Med. Microbiol. 2014, 4, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Epps, S.V.R.; Harvey, R.B.; Hume, M.E.; Phillips, T.D.; Anderson, R.C.; Nisbet, D.J. Foodborne Campylobacter: Infections, Metabolism, Pathogenesis, and Reservoirs. Int. J. Environ. Res. Public Health 2013, 10, 6292–6304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chansiripornchai, N.; Sasipreeyajan, J. PCR detection of four virulence-associated genes of Campylobacter jejuni isolates from Thai broilers and their abilities of adhesion to and invasion of INT-407 cells. J. Veter. Med. Sci. 2009, 71, 839–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribot, E.M.; Fitzgerald, C.; Kubota, K.; Swaminathan, B.; Barrett, T.J. Rapid Pulsed-Field Gel Electrophoresis Protocol for Subtyping of Campylobacter jejuni. J. Clin. Microbiol. 2001, 39, 1889–1894. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resistance Profiles | No. C. jejuni Isolates (n = 45) (%) | No. C. coli Isolates (n = 60) (%) |
|---|---|---|
| AMP-NAL-ERY-GEN-CRO-CLI | 8 (19.04) | 1 (1.67) |
| AMP-NAL-CIP-ERY-CRO-CLI | 11 (26.19) | 8 (13.3) |
| AMP-NAL-CIP-CRO-CLI | 3 (7.14) | 0 |
| AMP-NAL-ERY-GEN-CLI | 0 | 1 (1.67) |
| AMP-NAL-ERY-CRO-CLI | 2 (4.76) | 0 |
| AMP-NAL-ERY-TET-CRO-CLI | 3 (6.7) | 1 |
| AMP-CIP-ERY-CRO | 1 (2.3) | 0 |
| AMP-NAL-ERY-CRO | 1 (2.3) | 0 |
| NAL-CIP-ERY-CRO | 2 (4.76) | 0 |
| AMP-NAL-ERY-CLI | 0 | 3 (5.00) |
| AMP-CIP-ERY-CLI | 0 | 1 (1.67) |
| AMP-ERY-GENT-CLI | 1 (2.3) | 1 (1.67) |
| NAL-ERY-GENT-CLI | 1 (2.3) | 0 |
| AMP-NAL-ERY-GENT | 1 (2.3) | 0 |
| AMP-ERY-CRO-CLI | 0 | 1 (1.67) |
| ERY-CRO-CLI | 0 | 1 (1.67) |
| AMP-NAL-CRO | 1 (2.3) | 3 (5.00) |
| AMP-NAL-ERY | 1 (2.3) | 0 |
| ERY-GENT- CLI | 0 | 1 (1.67) |
| AMP-ERY-CLI | 0 | 2 (3.33) |
| NAL-ERY-CLI | 0 | 1 (1.67) |
| CIP-ERY-GENT | 0 | 1 (1.67) |
| ERY-CLI | 7 (16.67) | 14 (23.33) |
| AMP-CLI | 0 | 2 (3.33) |
| AMP-GENT | 0 | 2 (3.33) |
| AMP-ERY | 0 | 3 (5.00) |
| AMP- CRO | 1 (2.3) | 0 |
| ERY | 3 (7.14) | 4 (26.67) |
| CRO | 0 | 3 |
| CLI | 0 | 4 (26.67) |
| AMP | 0 | 2 (3.33) |
| Target Gene | Primer Sequence (5′–3′) | Product Size (bp) | Annealing Temperature (°C) | References |
|---|---|---|---|---|
| 16S rRNA | F-GGATGACACTTTTCGGAGC R-CATTGTAGCACGTGTGTC | 816 | 58 | [36] |
| Asp | F-GGTATGATTTCTACAAAGCGAG R-ATAAAAGACTATCGTCGCGTG | 500 | 53 | [37] |
| hipO | F-GAAGAGGGTTTGGGTGGT R-AGCTAGCTTCGCATAATAACTTG | 735 | 53 | [37] |
| Target Gene | Primer Sequence (5′–3′) | Product Size (bp) | Annealing Temperature (°C) | References |
|---|---|---|---|---|
| cmeB | F5′GACGTAATGAAGGAGAGCCA R5′CTGATCCACTCCAGCTATG | 1166 | 50 | [40] |
| tet(O) | F-GGCGTTTTGTTTATGTGCG R-ATGGACAACCCGACAGAAGC | 559 | 49 | [5] |
| 23S rRNA at position 2074 | F5′TTAGCTAATGTTGCCCGTACCG R5′AGTAAAGGTCCACGGGGTCTCG | 485 | 59 | [38] |
| 23S rRNA at position 2075 | F5′TTAGCTAATGTTGCCCGTACCG R5′TAGTAAAGGTCCACGGGGTCGC | 485 | 59 | [38] |
| gyrA | F-GAAGAATTTTATATGCTATG R-TCAGTATAAC GCATCGCAGC | 235 | 53 | [30] |
| gyrA | F-ACGCAAGAGAGATGGTT R-TCAGTATAACGCATCGCAGC | 270 | 45 | [30] |
| Target Gene | Primer Sequence (5′–3′) | Product Size (bp) | Annealing Temperature (°C) | References |
|---|---|---|---|---|
| cadF | F-TTGAAGGTAATTTAGATATG R-CTAATACCTAAAGTTGAAAC | 400 | 43 | [41] |
| ciaB | F-TGCGAGATTTTTCGAGAATG R-TGCCCGCCTTAGAACTTACA | 527 | 54 | [41] |
| dnaJ | F-ATTGATTTTGCTGCGGGTAG R-ATCCGCAAAAGCTTCAAAAA | 177 | 50 | [41] |
| pldA | F-AAGAGTGAGGCGAAATTCCA R-GCAAGATGGCAGGATTATCA | 385 | 46 | [41] |
| cdtA | F-CCTTGTGATGCAAGCAATC R-ACACTCCATTTGCTTTCTG | 370 | 49 | [21] |
| cdtB | F-GTTAAAATCCCCTGCTATCAACCA R-GTTGGCACTTGGAATTTGCAAGGC | 495 | 51 | |
| cdtC | F-CGATGAGTTAAAACAAAAAGATA R-TTGGCATTATAGAAAATACAGTT | 182 | 48 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pillay, S.; Amoako, D.G.; Abia, A.L.K.; Somboro, A.M.; Shobo, C.O.; Perrett, K.; Bester, L.A.; Essack, S.Y. Characterisation of Campylobacter spp. Isolated from Poultry in KwaZulu-Natal, South Africa. Antibiotics 2020, 9, 42. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9020042
Pillay S, Amoako DG, Abia ALK, Somboro AM, Shobo CO, Perrett K, Bester LA, Essack SY. Characterisation of Campylobacter spp. Isolated from Poultry in KwaZulu-Natal, South Africa. Antibiotics. 2020; 9(2):42. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9020042
Chicago/Turabian StylePillay, Stephanie, Daniel G. Amoako, Akebe L. K. Abia, Anou M. Somboro, Christiana O. Shobo, Keith Perrett, Linda A. Bester, and Sabiha Y. Essack. 2020. "Characterisation of Campylobacter spp. Isolated from Poultry in KwaZulu-Natal, South Africa" Antibiotics 9, no. 2: 42. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9020042