Saccharomyces Cerevisiae Var. Boulardii: Valuable Probiotic Starter for Craft Beer Production



1
Facultad de Veterinaria y Ciencias Experimentales, Universidad Católica de Valencia San Vicente Mártir, c/Guillem de Castro 94, 46001 Valencia, Spain
2
Departament d’Estadística i Investigació Operativa, Facultat de Matemàtiques, Universitat de València, c/Dr. Moliner, 50, 46100 Burjassot, València, Spain
*
Author to whom correspondence should be addressed.
Appl. Sci. 2019, 9(16), 3250; https://0-doi-org.brum.beds.ac.uk/10.3390/app9163250
Submission received: 10 June 2019
/
Revised: 3 August 2019
/
Accepted: 6 August 2019
/
Published: 8 August 2019
(This article belongs to the Section Applied Biosciences and Bioengineering)
Abstract
:Featured Application
Craft Beer Production.
Abstract
The use of probiotic starters remaining viable in unpasteurized and unfiltered beers could significantly increase health benefits. Here, the probiotic Saccharomyces cerevisiae var. boulardii (Scb) and a commercial Saccharomyces cerevisiae (Sc) strain, which is commonly employed in the brewing industry, are compared as single starters. The healthy value of the produced beers and growth performance in a laboratory bioreactor are analysed by determining antioxidant activity, phenolic content and profile, alcohol, biomass growth modelling by the logistic and Gompertz equations, biovolume estimation from 2D microscopy images, and yeast viability after fermentation. Thus, in this study, the craft beer produced with the probiotic yeast possessed higher antioxidant activity, lower alcohol content, similar sensory attributes, much higher yeast viability and more acidification, which is very desirable to reduce contamination risks at large-scale production. Furthermore, Scb exhibited faster growth in the bioreactor culture and larger cell volumes than Sc, which increases the probiotic volume of the final craft beer.

1. Introduction
The most important brewer’s yeasts are mostly part of the genus Saccharomyces. Thus, ale and lager yeasts belong to Saccharomyces cerevisiae (Sc) and Saccharomyces pastorianus, respectively [1]. S. pastorianus is a hybrid of Sc and Saccharomyces eubayanus. Sc is one of the most important brewing yeast species utilised for ale beer production due to its tolerance to alcohol [2], growth and fermentation rate on an industrial scale [3]. Saccharomyces cerevisiae var. boulardii (Scb) is another popular yeast species that was isolated from fruit in Indochina and has been utilised as a remedy for diarrhea since 1950 [4,5]. Scb has shown a capacity to prevent travellers’ diarrhea usually caused by bacteria such as Escherichia coli and Salmonella sp. [6,7].
Probiotics are microorganisms used as food supplements that in a correct dosage are potentially beneficial for human health, especially for the intestinal microbial balance [8]. Gastrointestinal microbiote imbalances can develop into several disorders, such as diarrhea under antibiotic treatment, ulcers, inflammatory bowel diseases or irritable bowel syndrome [9]. Scb is the only yeast species with probiotic properties able to keep gastrointestinal conditions at a suitable health level. Scb, in particular, has been very effective against Clostridium difficile in the prevention and treatment of antibiotic associated diarrhea [10,11] and may help to eradicate Helicobacter pylori [12]. In fact, the Scb yeast is now commercially available in most countries globally.
Scb shows enhanced ability for pseudohyphal switching upon nitrogen limitation and increased resistance to acidic pH, which are not correlated with increased adherence to epithelial cells or transit through mouse gut [13]. However, a recent genetic comparative study of 5 Scb against 145 Sc yeast strains has revealed the presence of two genes, HXT11 and HXT9, in Sc, which are not present in Scb [14]. The absence of these genes has been related to greater resistance against several antifungals, such as cycloheximide, sulfometuron methyl and 4-nitroquinoline-N-oxide [15]. Other genetic particularities that have been reported for Scb include the absence of ASP3 genes involved in asparagine degradation [16] and the abundant repetition of Flocculatıon (FLO) genes, which could explain the ability of Scb to adhere to the intestinal epithelium [17,18].
The optimal growth temperature of Scb is 37 °C and thus it can reach very high gastrointestinal tract (GT) concentrations in a very short period [19]. In addition, it possesses the capacity to inhibit the growth, adherence and invasion of multiple pathogens such C. difficile [20], E. coli [21], and Candida albicans [22], both in in vitro and in vivo conditions to the epithelial layer of the GT.
In recent years, there has been a worldwide increase in the consumer interest in craft beers, which are unpasteurized, unfiltered and without additional nitrogen or carbon dioxide pressure [23]. Even though there are many controversial studies about the beneficial effects of beer consumption on human health, epidemiological studies have shown that beer has positive effects on cardiovascular protection inhibiting atherosclerosis, decreasing the content of serum low-density lipoprotein cholesterol and triglycerides, by acting as an in vivo free radical scavenger [24,25].
This alcoholic beverage is usually mainly composed of four basic ingredients: water, hops, malt, and brewer’s yeast, as well as non-nutrient components such as phenolic compounds [26]. Brewer’s yeast strains are facultative anaerobes and this ability supports their property as an important industrial microorganism [27]. Phenolic compounds are present in vegetable foods such as malt and hops [28].
The use of Scb as a mixed starter with Sc for craft beer [29] or as a single starter for alcohol-free beer production [30] has been proposed recently. However, these results were preliminary and warrant further investigation in order to compare Scb as a single starter with other yeast strains such as S. cerevisiae Safale S-04, which is commonly employed in the brewing industry and especially selected for this purpose, in terms of health benefits (antioxidant activity, total phenolic content, polyphenol profile, alcohol formation) and probiotic value-added (biomass growth, cell volume and yeast viability after fermentation).
2. Materials and Methods
2.1. Materials
Methanol (HPLC grade), formic acid, Folin–Ciocalteu’s phenol reagent and sodium carbonate was acquired from Panreac (Barcelona, Spain). The 2,2-Diphenyl-1-picrylhydrazyl (DPPH) reagent, methanol (99.8%) and some polyphenol standard patterns (phloroglucinol, protocatechuic acid, epicatechin, quercetin, myricetin, ferulic acid, p-coumaric acid, rutin, gentisic, caffeic and catechin) were purchased from Sigma-Aldrich (Steinheim, Switzerland). Chlorogenic acid and vanillic acid polyphenol patterns were provided by HWI (Ruelzheim, Germany) and Fluka Analytical (Steinheim, Switzerland) respectively.
S. cerevisiae Safale S-04, hereafter referred to as SF-04, was provided by the Fermentis Division of S. I. Lesaffre (Lille, France). Saccharomyces cerevisiae var. boulardii CECT 1474 was purchased from the Spanish Type Culture Collection (CECT, Valencia, Spain). Sc and Scb were suspended in distilled water to be grown in Sabouraud Glucose Agar with Chloramphenicol (SGAC, Sigma-Aldrich, Steinheim, Switzerland) solid medium plates at their optimal culture temperatures, 27 and 37 °C, respectively, for isolation. After that, Sc and Scb were grown in yeast peptone dextrose (YPD) broth (Sigma-Aldrich, Steinheim, Switzerland) with an orbital shaker-incubator for 48 h also at 27 and 37 °C, respectively. Finally, 1-mL aliquots of pure cultures were subsequently frozen at −80 °C.
2.2. Wort Preparation and Beer Fermentation Procedure
The wort was prepared with a commercial Connoisseur Indian Pale Ale (IPA) bitter Kit (Muntons, UK) containing 100% hopped malt extract. This beer style was chosen because it is very popular in the craft-beer market [31]. For each yeast strain, 1.56 kg of this extract was mixed with 3.5 L of boiling drinking water. Then, pure cold drinking water was added to that mixture to reach a total volume of 20 L under vigorous mixing in a sterilized plastic fermenter. Subsequently, 5 mL of pure yeast culture at a concentration of 8 × 106 cell/mL was inoculated into this mixture. Finally, the fermenter was covered with a lid to maintain a sterile internal environment and was fermented in anaerobic conditions at ambient temperature (24 ± 1 °C) for 9 days. This temperature was maintained within the fermentation temperature range recommended by the SF-04 strain producer (Fermentis). After fermentation the beer was bottled and stored at 4 ± 0.5 °C.
The main analytical characteristics of the initial wort (pH, °Plato, density, and free amino nitrogen (FAN)) were measured with 3 10-mL aliquots in order to ensure its reproducibility. The FAN of the wort was determined according to the Sörensen analytical method [32]. Sugar consumption (in °Plato) and pH were measured as a function of time during the fermentation process. These fermentations were performed 3 times in order to ensure reproducibility.
2.3. Polyphenols and Antioxidant Activity
The total phenolic content (TPC) measurement is usually combined with the stable free radical 2,2-Diphenyl-1-picrylhydrazyl (DPPH) technique in order to estimate also the total antioxidant activity [33]. Thus, an UV/VIS Nanocolor UV0245 spectrophotometer (Macherey-Nagel, Düren, Germany) was used at different wavelengths (765 and 515 nm, respectively) to determine the TPC and antioxidant activity of the craft beers after 9 days fermentation. For each experiment, aliquots of 1 mL were taken from the fermented beers to remove CO2 gas by sonication for 10 min and then passed through a 0.2-µm filter.
2.3.1. Total Phenolic Content
TPC of the produced craft beers was determined by the Folin–Ciocalteu method [34]. A volume of 100 µL of the filtered and sonicated beer aliquots were introduced into a test tube with 8.4 mL of water and 0.5 mL of Folin–Ciocalteu reagent. The mixture was left to react for 5 min in the dark. Then, 1 mL of sodium carbonate (20%, v/v) was added to this mixture, stirred, and left in the dark at 27 ± 1 °C for 60 min. The TPC values were then determined by measuring the absorbance at 765 nm and applying the linear regression equation (y = 0.0009x + 0.0093), expressed in equivalents of gallic acid (GA) per L of beer (mg GA/L). These measurements were performed 6 times for each beer to ensure accurate results as both mean and standard deviation.
2.3.2. DPPH Scavenging Activity
The antioxidant activity was determined by the DPPH method [35]. A volume of 100 µL of the filtered and sonicated beer aliquots was placed in a test tube with 5 mL of mixture containing 0.005 g of DPPH in 200 mL of methanol. This mixture was well-stirred and left in the dark at 27 ± 1 °C for 20 min. The absorbance of this mixture (Asample) and that of a control sample (A0) with only methanol was then measured at 515 nm. Finally, the antioxidant activity of each craft beer could be determined by applying the radical scavenging activity (RSA) Equation (1).
These measurements were performed 6 times with each beer in order to guarantee the reproducibility of the results.
2.3.3. Polyphenol Profiles
The polyphenol profiles of the beers produced with the two yeast strains (Sc and Scb) were determined by chromatographic analysis after 9 days fermentation. Beer aliquots of 1 mL were centrifuged, passed through 0.20-μm filters and then sonicated to remove carbon dioxide prior to analysis.
High-performance liquid chromatography coupled with a L-2455 diode array detector (HPLC/DAD) was performed with a HPLC Hitachi 890-0442 (Tokyo, Japan) apparatus equipped with L-2300 column oven, L-2200U autosampler and L-2130 pump, connected to the EZChromEliteTM software. The separation was performed with a LiChrospher®100 RP-18 (Darmstadt, Germany) reverse phase column (250 mm × 4.6 mm, 5 µm) at 27 °C. A gradient system with two mobile phases was used: water with 0.1% formic acid (eluent A) and methanol (eluent B). The flow rate was 1.0 mL/min with an injection volume of 60 µL beer. The elution conditions applied consisted of 0–4 min of isocratic 10%B, 4–15 min linear gradient from 10% to 50%B and 15–30 min of isocratic 50%B. HPLC/DAD chromatographs for each craft beer were taken at 270, 324 and 373 nm.
Thirteen commercial polyphenol patterns usually found in beer [36] were used to identify and quantify the polyphenol profile in each beer sample. These polyphenols patterns were previously analysed to determine their retention times at the corresponding wavelengths (see Table 1) and the calibration lines employed to transform peak areas into concentrations.
The stock solutions with the polyphenol standards were prepared and diluted with methanol to the appropriate concentration in the range of 0.5–250 mg/L. Thus, 6 concentrations of each polyphenol pattern were prepared and injected in triplicate to construct the calibration lines (with R2 > 0.999) by plotting peak area versus concentration for each polyphenol.
These HPLC/DAD measurements were carried out in triplicate to be reported as mean and standard deviations.
2.4. Bioreactor Culture
The bioreactor cultures were treated in an autoclave (121° for 15 min) and performed in a 2-L Biostat A bench-top bioreactor (Sartorius, Frankfurt, Germany) containing 1 L of Sabouraud Glucose with Chloramphenicol liquid medium. This medium was prepared with 40 g/L of d-(+)-Glucose (≥99.5%, Sigma-Aldrich, Steinheim, Switzerland), 17.4 g/L of nutrient broth containing 5 g/L of peptone (Thermo Fisher Scientific (Oxoid), Berkshire, UK), 0.05 g/L chloramphenicol (≥98%, Sigma-Aldrich, Steinheim, Switzerland) and 25 g/L of Miller’s LB Broth with 10 g/L of tryptone (Sigma-Aldrich, Steinheim, Switzerland).
Bioreactor cultures were conducted in anaerobic conditions with a constant stirring speed of 100 rpm to ensure homogeneous mixing and low foam formation. Acidification was studied by continuous pH monitoring during 85 h of incubation time. Inoculation was performed with 5 mL of the yeast culture with an average concentration of 106 cells/mL. The experiments were carried out at 24 ± 1 °C in order to study cell growth at the same temperature to that used for the craft beer fermentations.
Yeast growth was analysed in the bioreactor culture by turbidimetry [37]. Thus, continuous spectrophotometric measurements with 10-mL aliquots were performed at 630 nm every 1 h for 50 h. These bioreactor culture determinations were carried out in triplicate in order to ensure reproducibility.
2.5. Growth Analysis and Modelling
Absorbance measurements at 630 nm (turbidimetry) as a function of time (t) were modelled by the Gompertz equation [38] and logistic function [39] (Equations (2) and (3), respectively) in order to study cell growth trends. In these models, there are three parameters: the maximum attainable value (K), the growth rate (r) and the x-axis displacement (a).
Experimental growth curves were also modelled by a particular Gompertz and logistic model (Equations (4) and (5), respectively) capable of treating data differently depending on the type of yeast employed. These mathematical models have only one parameter (r) and include an indicator variable (I) with a value of 0 for Sc and 1 for Scb. These equations also include three new parameters (s1, s2 and s3) to estimate the possible growth level of each yeast species by modifying K, r and a parameters as a function of time (t).
2.6. Biovolume Estimation
Equations for the estimation of cell volume (biovolume) for a wide range of microorganisms have become increasingly precise and powerful, especially since the introduction of allometric models [42]. Although laser and digital confocal microscopy can acquire 3D images, these techniques are time-consuming and expensive. In this regard, digital two-dimensional images acquired with an optical microscope have been recently proposed as a very promising alternative low-cost technique for biovolume mathematical estimation [43,44]. Thus, approximately 100 microscopy images of individual randomly selected cells (colonies were not considered) were taken for each yeast strain with a BA410 optical microscope (Motic, Kowloon, Hong Kong) provided with a Motic 580 5.0 MP camera and Motic Image Plus 3.0 software. These microscopy images were analysed with the aid of the R EBImage package [45]. Thus, the cell volumes were estimated from the 2D cell images by applying Equation (6) [43]. In this equation, A represents the estimated area of the cell cross-section in pixels according to the EBImage functionalities, d is the major axis of the ellipse that better fits their section and E is the exact area of the ellipse. In addition, the term is a correction factor to obtain a more realistic approximation of the real cell volume.
This method produces an average error below 1% even when some complex-shaped cells are analysed [42]. Therefore, since the analysed shapes of this study are quite simple (elliptical), the error must be almost negligible.
2.7. Yeast Viability
Cell viability of the yeasts present in the craft beers was evaluated by the colony counting method [48] after 45 days (9 days fermentation and 36 days stored at 4 °C) to determine the colony forming units (CFUs) per mL of viable cells in each craft beer. Thus, serial dilutions of the fermented craft beers were prepared in YPD. Then, 100 μL of each cell dilution was spread onto SGAC plates and grown at 37 °C for 72 h. This experiment was performed 3 times in order to ensure reproducibility.
2.8. Specific Gravity and Alcohol Concentration
The specific gravities of the wort (ρwort) and the fermented beers (ρbeer) were determined with an EasyDens meter (Anton-Paar GmbH, Austria, Graz). It is well-known that an accurate estimate of alcohol content can be determined from these specific gravity values [49]. However, ρbeer is an apparent value and its real value (ρ’beer) must be estimated with the Equation (7).
Thus, the alcohol percentage by weight (ABWt), as grams of alcohol per 100 g of beer, can be obtained by applying Equation (8) with ρwort and ρ’beer expressed in degrees Plato:
The alcohol percentage by volume of the produced beers (ABV) can be deduced with a correction to account for the specific gravity of the beers (ρbeer in g/cm3) in the following Equation (9).
2.9. Sensory Evaluation
The analysis for the content of main volatile compounds has recently shown that the use of Scb in combination with Sc as mixed starter does not negatively affect beer aroma [29]. Nonetheless, a deep sensory evaluation of attributes of the beers produced with Scb and Sc as single starters after 9 days of fermentation was performed by a panel of 10 trained experts. The descriptive assessment of attributes (appearance, aroma, flavor and bitterness) was carried out on a scale from 1 (extremely dislike attribute) to 9 (extremely like attribute).
2.10. Software
The R programming language (3.4.1 version, R Development Core Team, Vienna, Austria) [40] was used to perform the modelling and statistical analysis of the experimental results. Thus, the ggplot2 (plot creation), imputeTS (time series missing values imputation), nls (non-linear least squares), and EBImage (image analysis) R packages were selected for this study.
The statistical analysis (ANOVA and Tukey’s post hoc test) of the fermentation measurements (pH and °Plato), polyphenol concentrations and sensory evaluations were performed with SPSS22 at a significance level of p < 0.05.
3. Results
3.1. Physicochemical Parameters During Fermentation
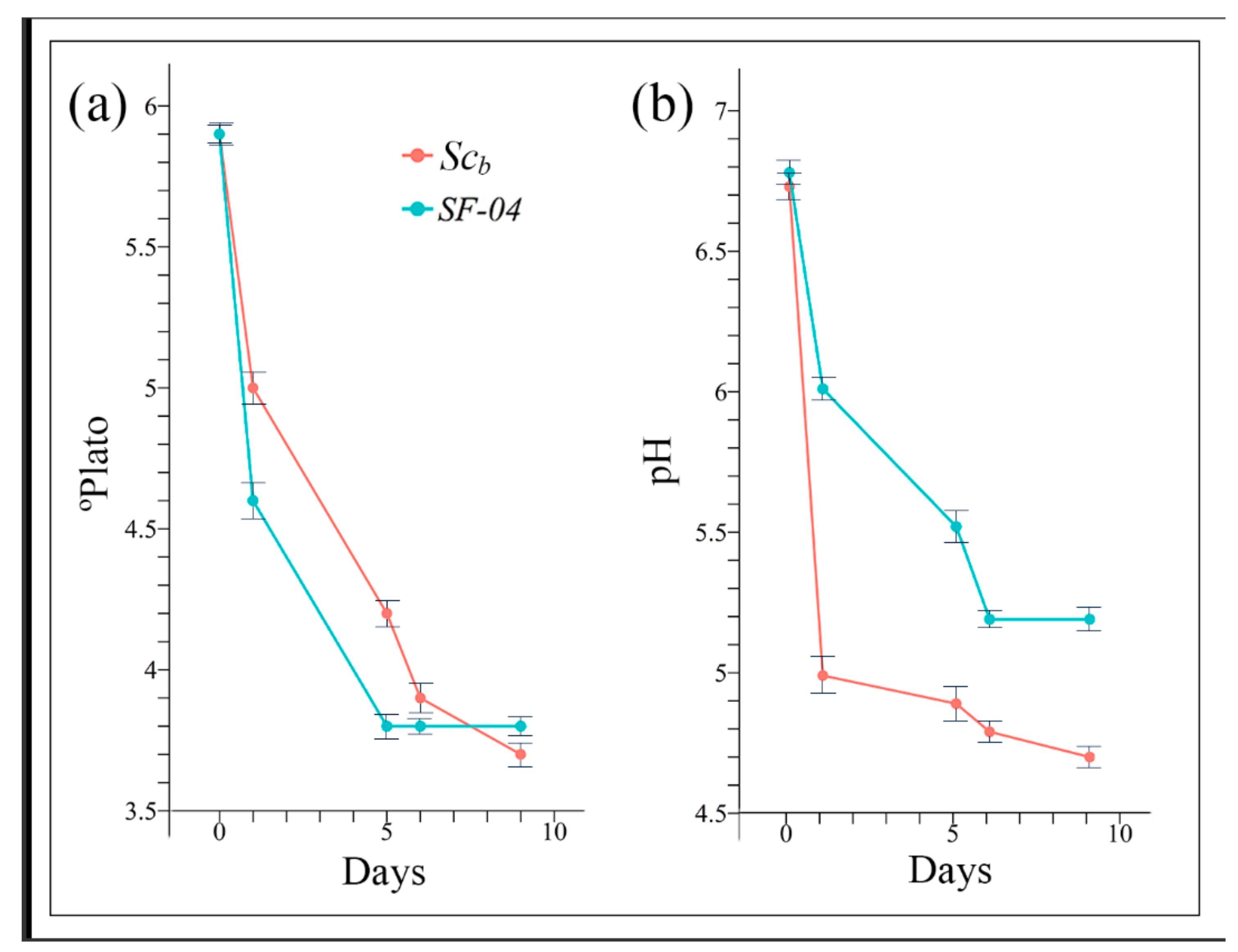
The main analytical characteristics of the initial unfermented wort were: pH 6.7 ± 0.2, 5.9 ± 0.6 °Plato and 150.8 ± 8.2 mg/L of FAN. It is of note that the pH of the wort is higher than usual due to the type of tap water used for its preparation. Thus, this water was collected from Valencia, Spain, and contains a high CaCO3 content. Since the goal of this study was focused on producing a craft beer with healthy added value, a low initial Plato value ensured a lower alcohol formation. Figure 1 shows the °Plato and pH measurements during the nine days of wort fermentation for the two yeast strains.
Figure 1a shows that the Plato degree trend followed a similar pattern independently from the type of yeast strain utilised. However, the pH achieved with Scb in the fermented beer was statistically significantly (p < 0.05) lower than with Sc during the whole fermentation process (see Figure 1b). This decrease of pH as a function of time was expected with both beers due to organic acid formation during fermentation [50]. Furthermore, FAN, which possess buffering capacity [51], is consumed by yeast during fermentation [52]. The stronger acidification achieved with this probiotic yeast during fermentation considerably reduces the contamination risk [53] and thus renders a more promising possibility for the production of this craft beer on a large scale. Furthermore, in a brewery, it has been reported that acidification may improve foam stability with a more well-rounded taste and increased shelf life in comparison to the non-acidified beer [54].
3.2. Antioxidant Activity and Total Phenolic Content After Fermentation
Phenolic compounds mostly come from both the malt and hops in beer [55] and will decrease from wort due to its degradation depending on the type of fermentation conditions [56]. However, in this study, the beers were produced with similar amounts and types of 100% hopped malt extract, employing the same experimental conditions. Thus, the TPC measurements determined by the Folin–Ciocalteu method showed similar results for both Scb and Sc species (see Table 2), within the experimental uncertainty expected.
However, the DPPH technique (RSA) revealed a significant increase of antioxidant activity for the craft beer produced with the probiotic Scb yeast, in good agreement with previous results of co-fermentation of Scb with Sc [29]. This increase of antioxidant activity of the beer fermented with Scb could be attributed to the presence of metabolites secreted by the Scb species such as proteins [57,58,59,60,61] with high antioxidant activity [62].
Table 2 shows that the alcohol content by weight (ABWt) and by volume (ABV), determined with ρwort and ρbeer in Equations (7)–(9), is lower in the beer produced with Scb than in the commercial beer.
3.3. HPLC/DAD Analysis After Fermentation
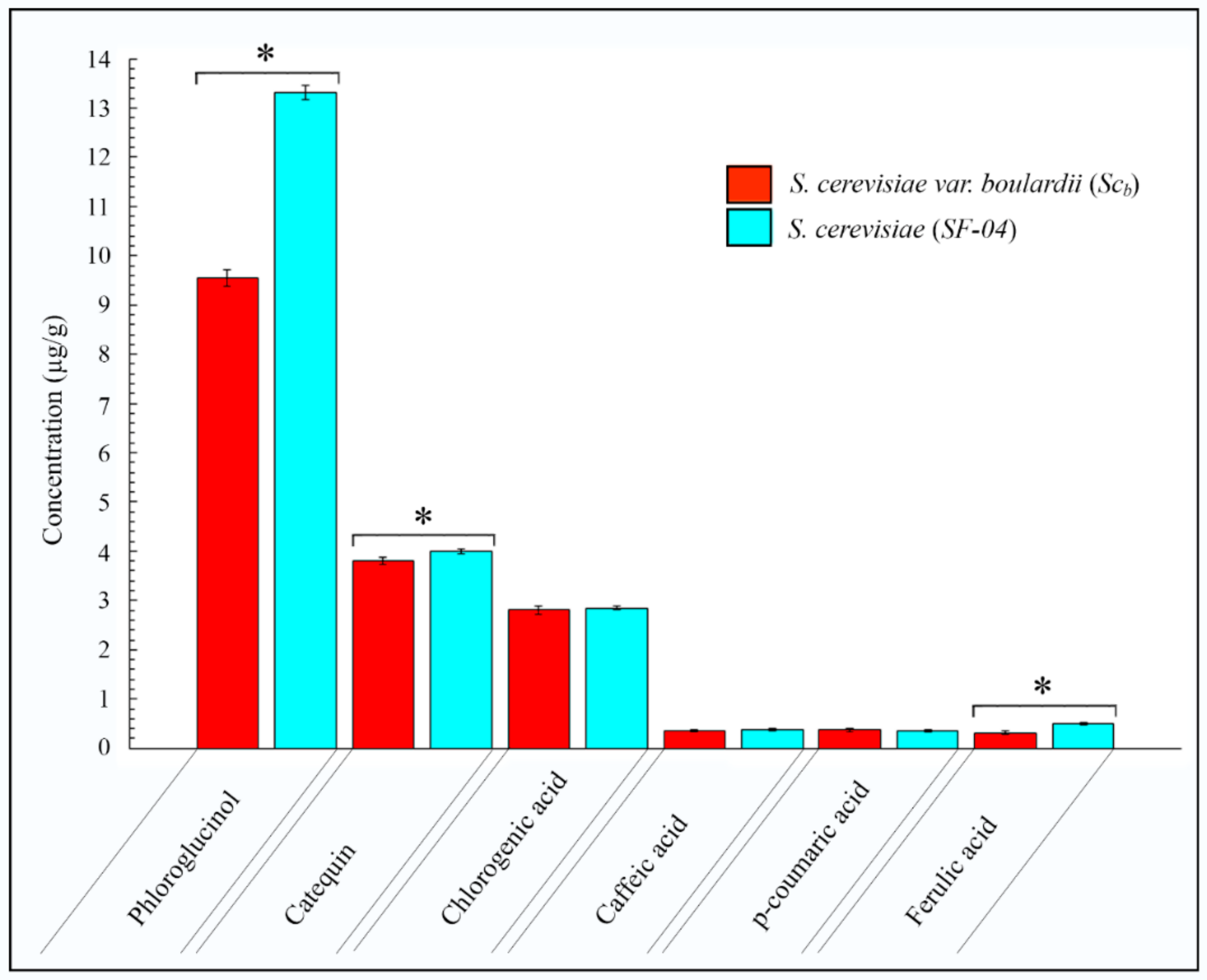
Even though the Folin–Ciocalteu (F–C) assay [34] is a standardized method to determine the phenolic content of food products and dietary supplements [63], an accurate HPLC/DAD analysis of the polyphenol types and contents was performed to confirm these results. Thus, the results of this analysis showed that, from the 13 polyphenol patterns, 6 peaks exhibited significant concentrations (see Figure 2).
These results confirm that both beers exhibit a similar polyphenol profile with a few polyphenols (phloroglucinol, catequin and ferulic acid) showing slightly higher concentrations in the beer produced with Sc (see Figure 2). Therefore, the higher antioxidant activity found in the beer produced with Scb could be attributed to antioxidant compounds other than polyphenols, secreted by the probiotic yeast. Thus, for example, the Scb yeast was found to produce (1→3)-β-d-glucan, which exhibits antioxidant and immunomodulation properties very suitable for anti-infective and antitumor therapy [64]. In addition, literature reports also suggest that Scb produces certain antioxidant metabolites responsible for its anti-inflammatory activity [58,60].
3.4. Yeast Growth Analysis in Bioreactor Culture
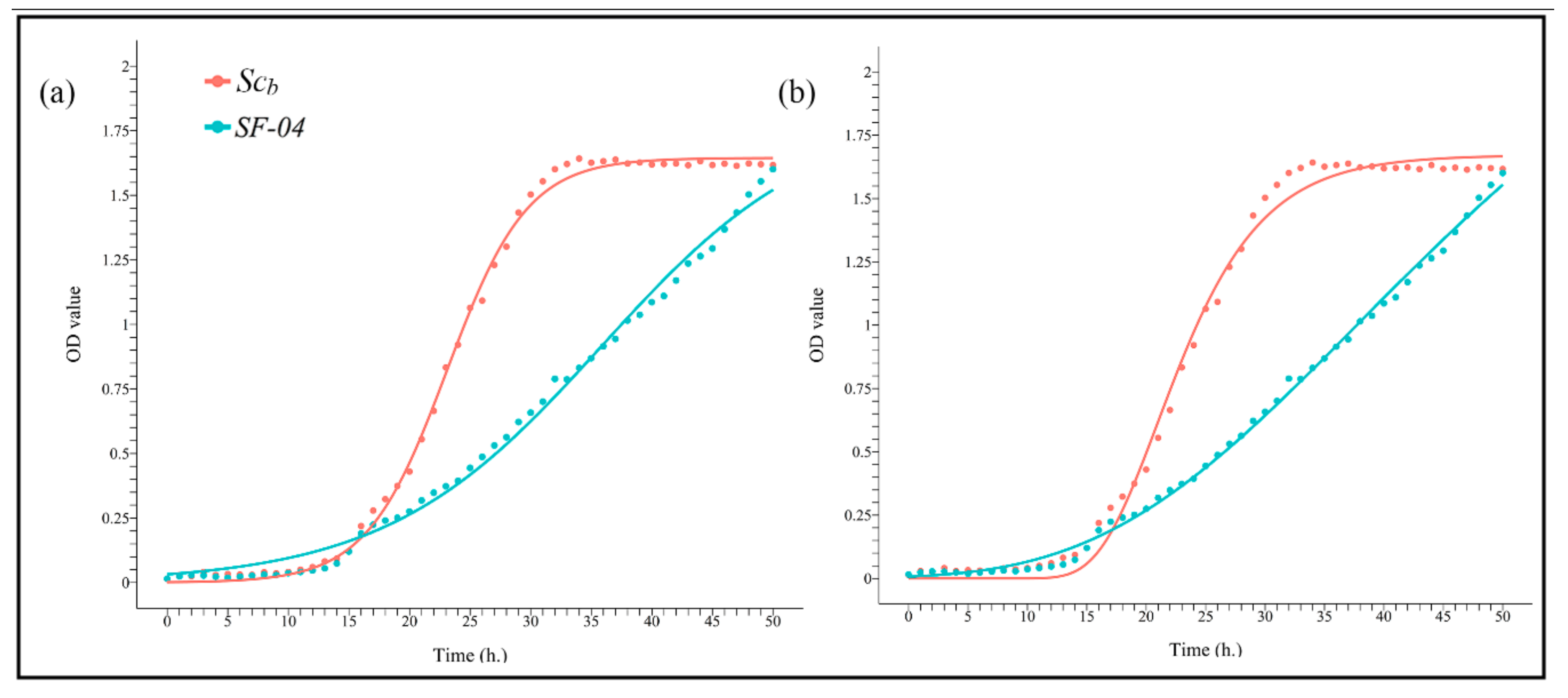
The use of the logistic function [39,65] and the Gompertz model [66,67] to represent the cell growth of several types of microorganisms as a function of time has been proved to be successful in many studies during the last decade. Since craft beers are unpasteurized and unfiltered, it is important to study cell growth at the same fermentation temperature (24 ± 1 °C) and anaerobic conditions in monitored bench-top bioreactors. Figure 3 shows the experimental turbidimetric results obtained during 50 h incubation and the mathematical growth modelling performed by logistic and Gompertz equations.
The estimated values of K, a and r obtained from the modelling of the turbidimetric measurements as a function of time are shown in Table 3 for the logistic and Gompertz models, respectively.
Both mathematical models clearly show superior growth rates for Scb (see r values in Table 3). In addition, Equations (4) and (5) confirmed the increased growth rates achieved by Scb in comparison to Sc. Indeed, these equations estimated significant s2 parameter values (p-value < 2 × 10−16) for Scb and Sc respectively, corresponding to different growth rates for both yeast species. A similar level of significance was found for the s3 parameter, which estimates the likely different growth levels for both yeast species by modifying the a parameter. Thus, this result confirms a different growth curve temporization because the a parameter is related to the displacement along the temporal x-axis. However, the s1 parameter was not significant for both Gompertz and logistic models, suggesting a similar saturation value for both species during a growth time cycle of 50 h.
A previous study also showed higher growth rates for Scb (Floratil®) in comparison with Sc (W303) at 30 and 37 °C [68]. Therefore, Scb grows faster than Sc at the same fermentation temperature of this study (24 ± 1 °C) and at the body temperature (37 °C).
3.5. Physicochemical Parameters During Bioreactor Culture
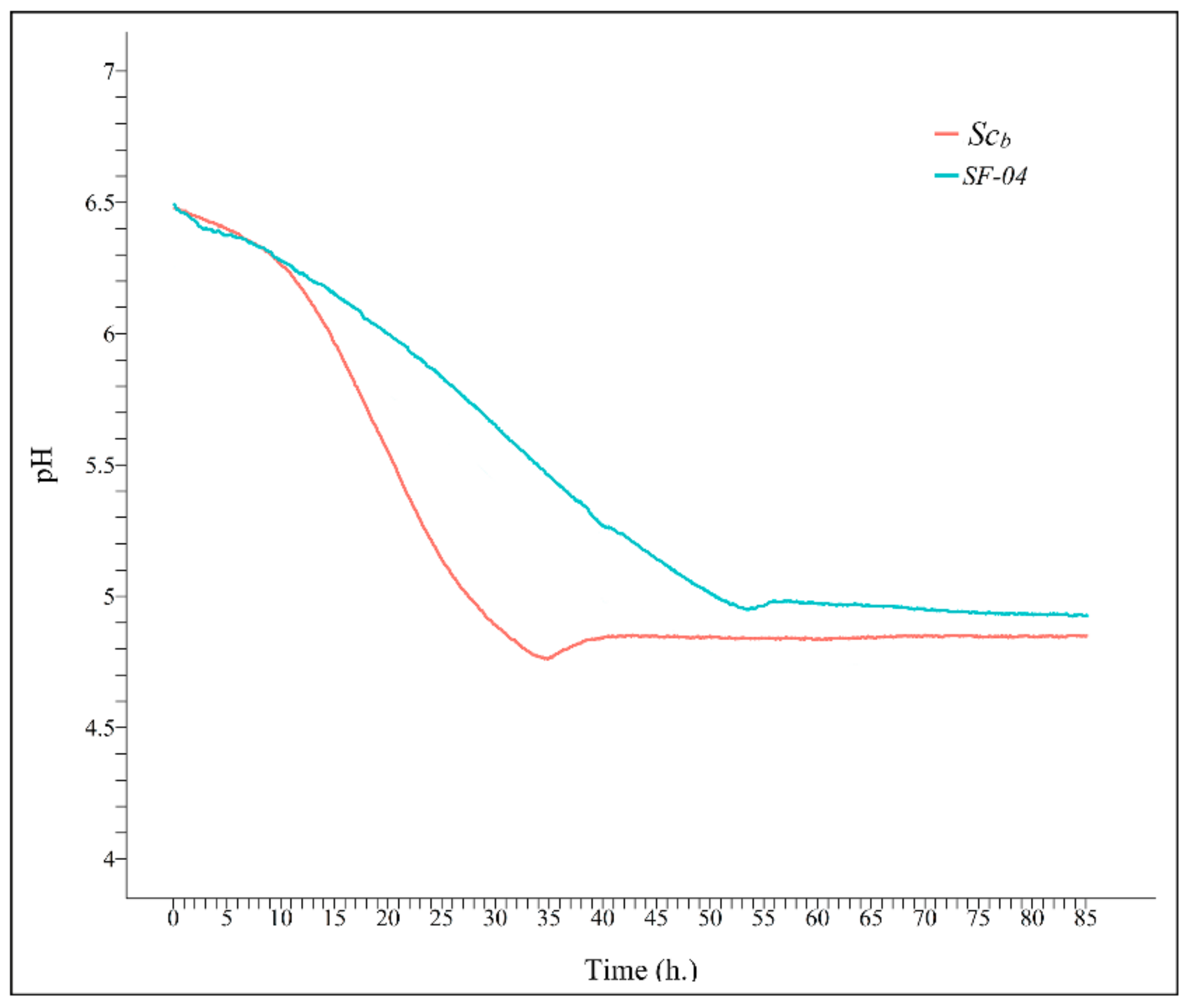
Figure 4 shows acidification as a function of time produced by Scb and Sc during bioreactor culture at 24 ± 1 °C.
These pH results are in accordance with the lower pH values achieved with Scb during fermentation (Figure 1b). Moreover, the specific gravity (in °Plato) for the Sc and Scb yeasts were also very similar at the beginning (9.2 ± 0.2 and 9.2 ± 0.1 °Plato, respectively) and at the end (4.0 ± 0.1 and 3.7 ± 0.2 °Plato, respectively) of fermentation during bioreactor culture (Figure 1a).
3.6. Yeast Biovolumes
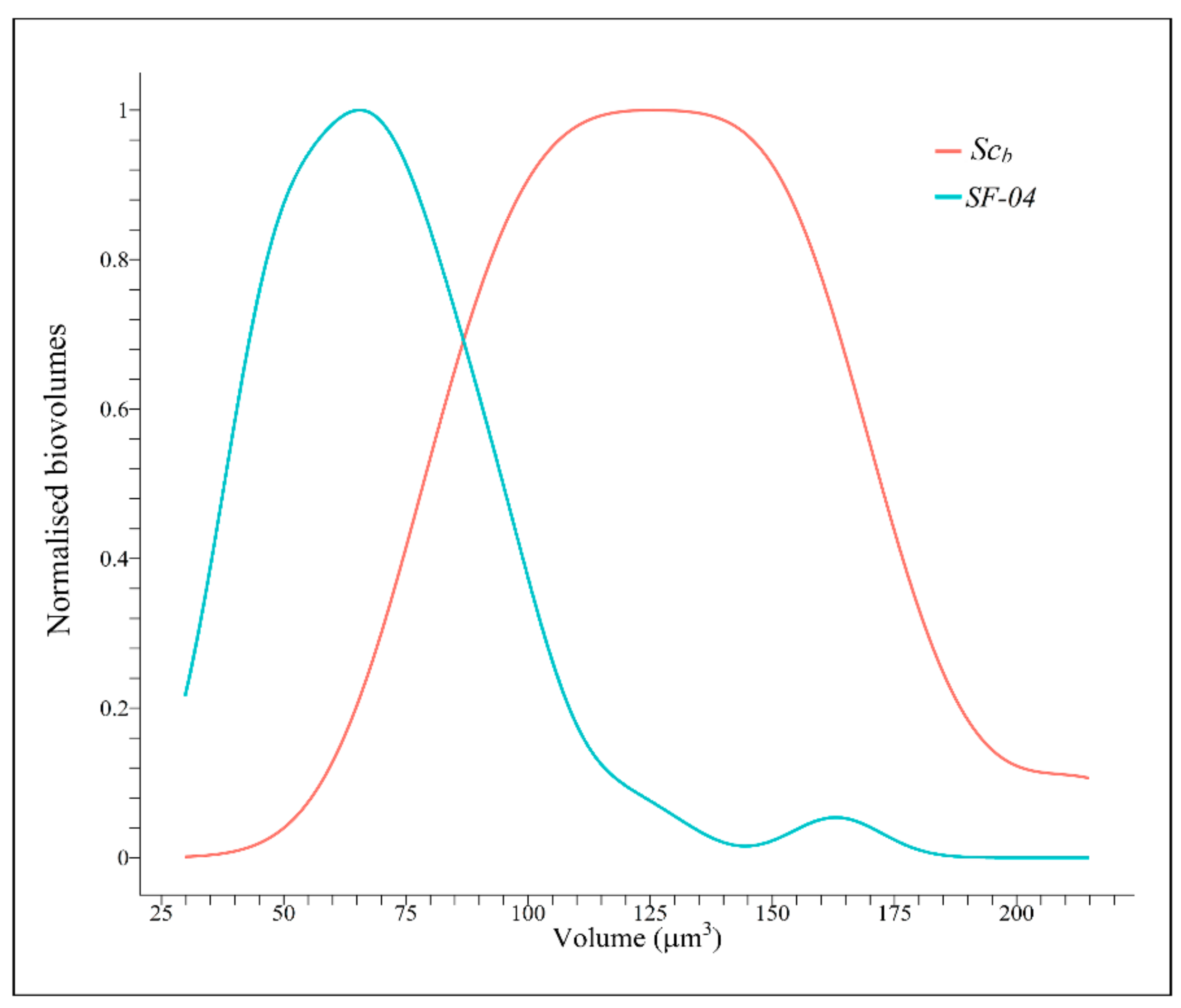
Figure 5 shows the biovolume value distribution results (in µm3) estimated from the 2D optical microscopy images for both yeast strains. The biovolumes were normalised and scaled to [0,1] to be comparable. Thus, the Scb cells exhibited higher average values than the Sc cells. The biovolumes of the Scb and Sc were 129.27 ± 32.90 and 70.72 ± 24.58 µm3, respectively. This result of biovolume of Sc is in good accordance with those cell volumes shown for this type of yeast [69,70]. However, as far as we know, the biovolumes of Scb have never been reported previously in the literature. The higher biovolume of Scb was confirmed with the use of a unilateral t-test, which yielded a p-value below 4.578 × 10−10 and thus allowed rejection of the hypothesis that both species possess equal mean biovolumes. In addition, the Kolmogorov–Smirnov test showed a p-value below 7.985 × 10−11, which led to rejection of the hypothesis that the two samples are drawn from the same continuous distribution.
3.7. Yeast Viability in the Craft Beers
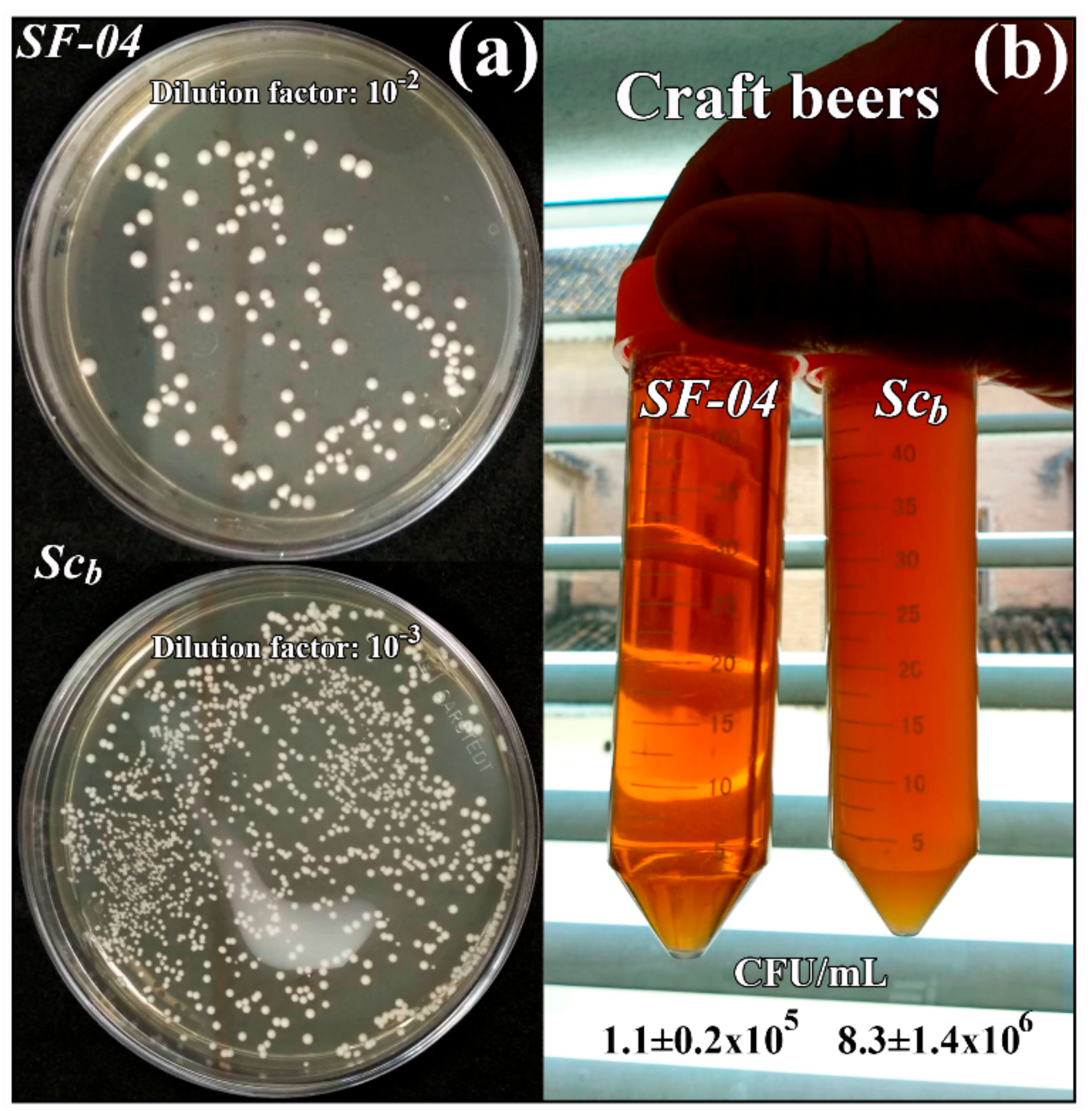
Since craft beers are largely unpasteurized and unfiltered, and their probiotic value depends on the amount of viable probiotic cells present in the beer after fermentation, yeast viability was measured in the craft beers by the colony counting method. Thus, Figure 6 shows the yeast viability results obtained for each craft beer after 45 days.
These results clearly show higher viability of Scb than Sc (see CFU/mL and the transparency of the Falcon tubes in Figure 6b) after fermentation, which is ideal for producing a craft beer containing viable probiotic cells.
Table 4 shows that both yeasts produce craft beers with similar sensory attributes. Thus, these results are in good agreement with previous studies demonstrating that the use of Scb does not negatively affect beer aroma [29] and produces craft beer with acceptable sensory attributes [30].
Therefore, all the results obtained in this and previous studies render Scb as a very promising yeast for craft beer production with increased health benefits as a result of large-scale production. However, the results of the present study have been determined at a small scale and thus cannot be directly extended to large-scale production.
4. Conclusions
In this study, probiotic Saccharomyces cerevisiae var. boulardii as a single yeast starter has been shown to produce craft beer with higher antioxidant activity, lower alcohol content, similar sensory attributes and higher yeast viability after 45 days than that produced by a commercial Saccharomyces cerevisiae strain commonly used in the brewery industry. Furthermore, it produced higher acidification in bioreactor culture and also during fermentation, which is desirable to avoid contamination risks during large-scale production. Since polyphenols come mostly from hops and malt in beer, both craft beers resulted in a similar phenolic content and polyphenol profile. Scb showed faster growth than Sc in the batch bioreactor culture at 24 °C and the microscopy analysis revealed superior cell volumes for the Scb yeast, which increases the probiotic volume in the final craft beer. However, these results were obtained at the 20-L scale and thus more research should be performed before considering this promising beverage for large-scale industrial production.
Author Contributions
J.M.-C. and Á.S.-A. conceived the idea of this study. J.M.-C. performed all the experiments. Á.B.-R. performed the mathematical analysis and contributed to the manuscript text and figures of this part. Á.S.-A. got financial support, designed the experimental planning, supervised, analysed and discussed the data results, and prepared the draft and final manuscript text and figures.
Funding
This research was funded by the Universidad Católica de Valencia San Vicente Mártir through the PRUCV2018-231-001 and PRUCV 2019-231-003 grants (awarded to Á.S.-A).
Acknowledgments
The authors would like to acknowledge the Universidad Católica de Valencia San Vicente Mártir for the financial support, the Biotechnologist Laura Salinero Martín for her help with the bioreactor batch cultures, and both Carolina Padrón and Mariola Belda for their help with the HPLC/DAD measurements.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. The Yeasts: A Taxonomic Study; Elsevier: Amsterdam, The Netherlands, 2011; ISBN 9780080931272. [Google Scholar]
- Ding, J.; Huang, X.; Zhang, L.; Zhao, N.; Yang, D.; Zhang, K. Tolerance and stress response to ethanol in the yeast Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2009, 85, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Lodolo, E.J.; Kock, J.L.F.; Axcell, B.C.; Brooks, M. The yeast Saccharomyces cerevisiae—The main character in beer brewing. In FEMS Yeast Research; Wiley/Blackwell: Hoboken, NJ, USA, 2008; Volume 8, pp. 1018–1036. [Google Scholar]
- McCullough, M.J.; Clemons, K.V.; Mccusker, J.H.; Stevens, D.A. Species identification and virulence attributes of Saccharomyces boulardii. J. Clin. Microbiol. 1998, 36, 2613–2617. [Google Scholar] [PubMed]
- Hancox, L.R.; Le Bon, M.; Richards, P.J.; Guillou, D.; Dodd, C.E.R.; Mellits, K.H. Effect of a single dose of Saccharomyces cerevisiae var. boulardii on the occurrence of porcine neonatal diarrhoea. Animal 2015, 9, 1756–1759. [Google Scholar] [CrossRef] [PubMed]
- Leggat, P.A.; Goldsmid, J.M. Others the returned traveller with diarrhoea. Aust. Fam. Physician 2007, 36, 322. [Google Scholar]
- McFarland, L.V. Meta-analysis of probiotics for the prevention of traveler’s diarrhea. Travel Med. Infect. Dis. 2007, 5, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Moslehi-Jenabian, S.; Pedersen, L.L.; Jespersen, L. Beneficial effects of probiotic and food borne yeasts on human health. Nutrients 2010, 2, 449–473. [Google Scholar] [CrossRef] [PubMed]
- Czerucka, D.; Piche, T.; Rampal, P. Review article: Yeast as probiotics—Saccharomyces boulardii. Aliment. Pharmacol. Ther. 2007, 26, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Kabbani, T.A.; Pallav, K.; Dowd, S.E.; Villafuerte-Galvez, J.; Vanga, R.R.; Castillo, N.E.; Hansen, J.; Dennis, M.; Leffler, D.A.; Kelly, C.P. Prospective randomized controlled study on the effects of Saccharomyces boulardii CNCM I-745 and amoxicillin-clavulanate or the combination on the gut microbiota of healthy volunteers. Gut Microbes 2017, 8, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Surawicz, C.M. Treatment of recurrent Clostridium difficile-associated disease. Nat. Clin. Pract. Gastroenterol. Hepatol. 2004, 1, 32–38. [Google Scholar] [CrossRef]
- Greenhill, C. Probiotic helps to eradicate H. pylori. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 362. [Google Scholar] [CrossRef]
- Edwards-Ingram, L.; Gitsham, P.; Burton, N.; Warhurst, G.; Clarke, I.; Hoyle, D.; Oliver, S.G.; Stateva, L. Genotypic and physiological characterization of Saccharomyces boulardii, the probiotic strain of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2007, 73, 2458–2467. [Google Scholar] [CrossRef] [PubMed]
- Khatri, I.; Tomar, R.; Ganesan, K.; Prasad, G.S.; Subramanian, S. Complete genome sequence and comparative genomics of the probiotic yeast Saccharomyces boulardii. Sci. Rep. 2017, 7, 371. [Google Scholar] [CrossRef] [PubMed]
- Nourani, A.; Wesolowski-Louvel, M.; Delaveau, T.; Jacq, C.; Delahodde, A. Multiple-drug-resistance phenomenon in the yeast Saccharomyces cerevisiae: Involvement of two hexose transporters. Mol. Cell. Biol. 1997, 17, 5453–5460. [Google Scholar] [CrossRef] [PubMed]
- League, G.P.; Slot, J.C.; Rokas, A. The ASP3 locus in Saccharomyces cerevisiae originated by horizontal gene transfer from Wickerhamomyces. FEMS Yeast Res. 2012, 12, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, K.J.; Klis, F.M. Flocculation, adhesion and biofilm formation in yeasts. Mol. Microbiol. 2006, 60, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Stratford, M. Genetic aspects of yeast flocculation: In particular, the role of FLO genes in the flocculation of Saccharomyces cerevisiae. Colloids Sur. B Biointerfaces 1994, 2, 151–158. [Google Scholar] [CrossRef]
- Karen, M.; Yuksel, O.; Akyürek, N.; Ofluoǧlu, E.; Çaǧlar, K.; Şahin, T.T.; Paşaoǧlu, H.; Memiş, L.; Akyürek, N.; Bostanci, H. Probiotic Agent Saccharomyces boulardii Reduces the Incidenceof Lung Injury in Acute Necrotizing Pancreatitis Induced Rats. J. Surg. Res. 2010, 160, 139–144. [Google Scholar] [CrossRef]
- Surawicz, C.M.; McFarland, L.V.; Greenberg, R.N.; Rubin, M.; Fekety, R.; Mulligan, M.E.; Garcia, R.J.; Brandmarker, S.; Bowen, K.; Borjal, D.; et al. The Search for a Better Treatment for Recurrent Clostridium difficile Disease: Use of High-Dose Vancomycin Combined with Saccharomyces boulardii. Clin. Infect. Dis. 2000, 31, 1012–1017. [Google Scholar] [CrossRef]
- Czerucka, D.; Dahan, S.; Mograbi, B.; Rossi, B.; Rampal, P. Saccharomyces boulardii preserves the barrier function and modulates the signal transduction pathway induced in enteropathogenic Escherichia coli-infected T84 cells. Infect. Immun. 2000, 68, 5998–6004. [Google Scholar] [CrossRef]
- Berg, R.; Bernasconi, P.; Fowler, D.; Gautreaux, M. Inhibition of Candida albicans Translocation from the Gastrointestinal Tract of Mice by Oral Administration of Saccharomyces boulardii. J. Infect. Dis. 1993, 168, 1314–1318. [Google Scholar] [CrossRef]
- Canonico, L.; Comitini, F.; Ciani, M. Dominance and influence of selected Saccharomyces cerevisiae strains on the analytical profile of craft beer refermentation. J. Inst. Brew. 2014, 120, 262–267. [Google Scholar] [CrossRef]
- Granato, D.; Branco, G.F.; de Faria, A.J.; Cruz, A.G. Characterization of Brazilian lager and brown ale beers based on color, phenolic compounds, and antioxidant activity using chemometrics. J. Sci. Food Agric. 2011, 91, 563–571. [Google Scholar] [CrossRef] [PubMed]
- De Gaetano, G.; Cerletti, C.; Alkerwi, A.; Iacoviello, L.; Badimon, L.; Costanzo, S.; Pounis, G.; Trevisan, M.; Panico, S.; Stranges, S.; et al. Effects of moderate beer consumption on health and disease: A consensus document. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 443–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esslinger, H.M. Handbook of Brewing: Processes, Technology, Markets; Wiley-VCH: Hoboken, NJ, USA, 2009; ISBN 3527623493. [Google Scholar]
- Stewart, G. Saccharomyces species in the Production of Beer. Beverages 2016, 2, 34. [Google Scholar] [CrossRef]
- Callemien, D.; Collin, S. Structure, organoleptic properties, quantification methods, and stability of phenolic compounds in beer-A review. Food Rev. Int. 2010, 26, 1–84. [Google Scholar] [CrossRef]
- Capece, A.; Romaniello, R.; Pietrafesa, A.; Siesto, G.; Pietrafesa, R.; Zambuto, M.; Romano, P. Use of Saccharomyces cerevisiae var. boulardii in co-fermentations with S. cerevisiae for the production of craft beers with potential healthy value-added. Int. J. Food Microbiol. 2018, 284, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Senkarcinova, B.; Graça Dias, I.A.; Nespor, J.; Branyik, T. Probiotic alcohol-free beer made with Saccharomyces cerevisiae var. boulardii. LWT 2019, 100, 362–367. [Google Scholar] [CrossRef]
- Sanekata, A.; Tanigawa, A.; Takoi, K.; Nakayama, Y.; Tsuchiya, Y. Identification and Characterization of Geranic Acid as a Unique Flavor Compound of Hops (Humulus lupulus L.) Variety Sorachi Ace. J. Agric. Food Chem. 2018, 66, 12285–12295. [Google Scholar] [CrossRef]
- Bishop, L.R. The Nitrogen Compounds of Wort And Beer. J. Inst. Brew. 1943, 49, 173–178. [Google Scholar] [CrossRef]
- Hseu, Y.-C.; Chang, W.-H.; Chen, C.-S.; Liao, J.-W.; Huang, C.-J.; Lu, F.-J.; Chia, Y.-C.; Hsu, H.-K.; Wu, J.-J.; Yang, H.-L. Antioxidant activities of Toona Sinensis leaves extracts using different antioxidant models. Food Chem. Toxicol. 2008, 46, 105–114. [Google Scholar] [CrossRef]
- Julkunen-Tiitto, R. Phenolic Constituents in the Leaves of Northern Willows: Methods for the Analysis of Certain Phenolics. J. Agric. Food Chem. 1985, 33, 213–217. [Google Scholar] [CrossRef]
- Ditrych, M.; Kordialik-Bogacka, E.; Czyżowska, A. Antiradical and Reducing Potential of Commercial Beers. Czech. J. Food Sci. 2015, 33. [Google Scholar] [CrossRef]
- Quifer-Rada, P.; Vallverdú-Queralt, A.; Martínez-Huélamo, M.; Chiva-Blanch, G.; Jáuregui, O.; Estruch, R.; Lamuela-Raventós, R. A comprehensive characterisation of beer polyphenols by high resolution mass spectrometry (LC-ESI-LTQ-Orbitrap-MS). Food Chem. 2015, 169, 336–343. [Google Scholar] [CrossRef]
- Bovee, T.F.H.; Helsdingen, R.J.R.; Hamers, A.R.M.; van Duursen, M.B.M.; Nielen, M.W.F.; Hoogenboom, R.L.A.P. A new highly specific and robust yeast androgen bioassay for the detection of agonists and antagonists. Anal. Bioanal. Chem. 2007, 389, 1549–1558. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, R.L.; Whiting, R.C.; Damert, W.C. When is simple good enough: A comparison of the Gompertz, Baranyi, and three-phase linear models for fitting bacterial growth curves. Food Microbiol. 1997, 14, 313–326. [Google Scholar] [CrossRef]
- Kacena, M.A.; Merrell, G.A.; Manfredi, B.; Smith, E.E.; Klaus, D.M.; Todd, P. Bacterial growth in space flight: Logistic growth curve parameters for Escherichia coli and Bacillus subtilis. Appl. Microbiol. Biotechnol. 1999, 51, 229–234. [Google Scholar] [CrossRef]
- R Core Team. R Language Definition; R Foundation for Statistical Computing: Vienna, Austria, 2000. [Google Scholar]
- Nonlinear Regression Analysis and Its Applications; Bates, D.M.; Watts, D.G. (Eds.) Wiley Series in Probability and Statistics; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1988. [Google Scholar]
- Saccà, A. Methods for the estimation of the biovolume of microorganisms: A critical review. Limnol. Oceanogr. Methods 2017, 15, 337–348. [Google Scholar] [CrossRef]
- Saccà, A. A simple yet accurate method for the estimation of the biovolume of planktonic microorganisms. PLoS ONE 2016, 11, e0151955. [Google Scholar] [CrossRef]
- Lenz, M.; Roumans, N.J.T.; Vink, R.G.; Van Baak, M.A.; Mariman, E.C.M.; Arts, I.C.W.; De Kok, T.M.; Ertaylan, G. Estimating real cell size distribution from cross-section microscopy imaging. Bioinformatics 2016, 32, 396–404. [Google Scholar] [CrossRef]
- Pau, G.; Fuchs, F.; Sklyar, O.; Boutros, M.; Huber, W. EBImage--an R package for image processing with applications to cellular phenotypes. Bioinformatics 2010, 26, 979–981. [Google Scholar] [CrossRef]
- Jekel, J.F.; Katz, D.L.; Elmore, J.G.; Wild, D. Epidemiology, Biostatistics and Preventive Medicine; Elsevier Health Sciences: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Massey, F.J. The Kolmogorov-Smirnov Test for Goodness of Fit. J. Am. Stat. Assoc. 1951, 46, 68–78. [Google Scholar] [CrossRef]
- Martí, M.; Frígols, B.; Serrano-Aroca, Á. Antimicrobial Characterization of Advanced Materials for Bioengineering Applications. J. Vis. Exp. 2018, e57710. [Google Scholar] [CrossRef] [Green Version]
- Spedding, G. Alcohol and Its Measurement. In Brewing Materials and Processes: A Practical Approach to Beer Excellence; Academic Press: Cambridge, MA, USA, 2016; pp. 123–149. [Google Scholar]
- Erkan, H.; Çelik, S.; Bilgi, B.; Köksel, H. A new approach for the utilization of barley in food products: Barley tarhana. Food Chem. 2006, 97, 12–18. [Google Scholar] [CrossRef]
- Agu, R.C.; Palmer, G.H. A reassessment of sorghum for lager-beer brewing. Bioresour. Technol. 1998, 66, 253–261. [Google Scholar] [CrossRef]
- Verbelen, P.J.; De Schutter, D.P.; Delvaux, F.; Verstrepen, K.J.; Delvaux, F.R. Immobilized yeast cell systems for continuous fermentation applications. Biotechnol. Lett. 2006, 28, 1515–1525. [Google Scholar] [CrossRef]
- Strezov, V.; Evans, T.J. Biomass Processing Technologies; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Lowe, D.P.; Ulmer, H.M.; Barta, R.C.; Goode, D.L.; Arendt, E.K. Biological acidification of a mash containing 20% barley using Lactobacillus amylovorus FST 1.1: Its effects on Wort and beer quality. J. Am. Soc. Brew. Chem. 2005, 63, 96–106. [Google Scholar] [CrossRef]
- Araki, S.; Kimura, T.; Shimizu, C.; Furusho, S.; Takashio, M.; Shinotsuka, K. Estimation of Antioxidative Activity and its Relationship to Beer Flavor Stability. J. Am. Soc. Brew. Chem. 1999, 57, 34–37. [Google Scholar] [CrossRef]
- Montanari, L.; Perretti, G.; Natella, F.; Guidi, A.; Fantozzi, P. Organic and phenolic acids in beer. LWT-Food Sci. Technol. 1999, 32, 535–539. [Google Scholar] [CrossRef]
- Pothoulakis, C.; Kelly, C.P.; Joshi, M.A.; Gao, N.; O’Keane, C.J.; Castagliuolo, I.; Lamont, J.T. Saccharomyces boulardii inhibits Clostridium difficile toxin A binding and enterotoxicity in rat ileum. Gastroenterology 1993, 104, 1108–1115. [Google Scholar] [CrossRef]
- Castagliuolo, I.; Riegler, M.F.; Valenick, L.; LaMont, J.T.; Pothoulakis, C. Saccharomyces boulardii protease inhibits the effects of Clostridium difficile toxins A and B in human colonic mucosa. Infect. Immun. 1999, 67, 302–307. [Google Scholar] [CrossRef]
- Czerucka, D.; Roux, I.; Rampal, P. Saccharomyces boulardii inhibits secretagogue-mediated adenosine 3′, 5′-cyclic monophosphate induction in intestinal cells. Gastroenterology 1994, 106, 65–72. [Google Scholar] [CrossRef]
- Castagliuolo, I.; Thomas Lamont, J.; Nikulasson, S.T.; Pothoulakis, C. Saccharomyces boulardii protease inhibits Clostridium difficile toxin A effects in the rat ileum. Infect. Immun. 1996, 64, 5225–5232. [Google Scholar]
- Chen, X.; Kokkotou, E.G.; Mustafa, N.; Bhaskar, K.R.; Sougioultzis, S.; O’Brien, M.; Pothoulakis, C.; Kelly, C.P. Saccharomyces boulardii inhibits ERK1/2 mitogen-activated protein kinase activation both in vitro and in vivo and protects against Clostridium difficile toxin A-induced enteritis. J. Biol. Chem. 2006, 281, 24449–24454. [Google Scholar] [CrossRef]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant activity of proteins and peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Kogan, G.; Pajtinka, M.; Babincova, M.; Miadokova, E.; Rauko, P.; Slamenova, D.; Korolenko, T.A. Yeast cell wall polysaccharides as antioxidants and antimutagens: Can they fight cancer. In Neoplasma; In Tech: Rijeka, Croatia, 2008; Volume 55, pp. 387–393. [Google Scholar]
- Pearl, R.; Slobodkin, L. The Growth of Populations. Q. Rev. Biol. 1976, 51, 6–24. [Google Scholar] [CrossRef]
- Chowdhury, B.R.; Chakraborty, R.; Chaudhuri, U.R. Validity of modified Gompertz and Logistic models in predicting cell growth of Pediococcus acidilactici H during the production of bacteriocin pediocin AcH. J. Food Eng. 2007, 80, 1171–1175. [Google Scholar] [CrossRef]
- Chirivella-Martorell, J.; Briz-Redón, Á.; Serrano-Aroca, Á. Modelling of Biomass Concentration, Multi-Wavelength Absorption and Discrimination Method for Seven Important Marine Microalgae Species. Energies 2018, 11, 1089. [Google Scholar] [CrossRef]
- Fietto, J.L.; Araújo, R.S.; Valadão, F.N.; Fietto, L.G.; Brandão, R.L.; Neves, M.J.; Gomes, F.C.; Nicoli, J.R.; Castro, I.M. Molecular and physiological comparisons between Saccharomyces cerevisiae and Saccharomyces boulardii. Can. J. Microbiol. 2004, 50, 615–621. [Google Scholar] [CrossRef]
- Sun, J.; Yang, J.; Gao, Y.; Xu, D.; Li, D. Reference channel-based microfluidic resistance sensing for single yeast cell volume growth measurement. Microfluid. Nanofluidics 2017, 21, 33. [Google Scholar] [CrossRef]
- Wang, J.; Mao, J.; Yang, G.; Zheng, F.; Niu, C.; Li, Y.; Liu, C.; Li, Q. The FKS family genes cause changes in cell wall morphology resulted in regulation of anti-autolytic ability in Saccharomyces cerevisiae. Bioresour. Technol. 2018, 249, 49–56. [Google Scholar] [CrossRef]
Figure 1.
Plato degrees (a) and pH (b) evolution during beer fermentation using the Saccharomyces cerevisiae var. boulardii (Scb) and Saccharomyces cerevisiae (SF-04) strains as single starters. Results shown as mean ± standard deviation.
Figure 1.
Plato degrees (a) and pH (b) evolution during beer fermentation using the Saccharomyces cerevisiae var. boulardii (Scb) and Saccharomyces cerevisiae (SF-04) strains as single starters. Results shown as mean ± standard deviation.

Figure 2.
Hıgh performance lıquıd chromatograpy results for the beers fermented with the Saccharomyces cerevisiae var. boulardii (SCb) and Saccharomyces cerevisiae (SF-04) yeast strains. The polyphenols were detected at three different wavelengths (270 nm, 324 nm and 373 nm). Results shown as mean ± standard deviation. * Statistically significant difference of polyphenol concentrations (p < 0.05).
Figure 2.
Hıgh performance lıquıd chromatograpy results for the beers fermented with the Saccharomyces cerevisiae var. boulardii (SCb) and Saccharomyces cerevisiae (SF-04) yeast strains. The polyphenols were detected at three different wavelengths (270 nm, 324 nm and 373 nm). Results shown as mean ± standard deviation. * Statistically significant difference of polyphenol concentrations (p < 0.05).

Figure 3.
Experimental values (points) and mathematical modelling (continuous lines) of absorbance measurements at 630 nm as a function of time for the Saccharomyces cerevisiae (SF-04) and Saccharomyces cerevisiae var. boulardıi (SCb) strains cultured in batch bioreactor in anaerobic conditions at 24 ± 1 °C. Mathematical modelling was performed with the logistic (a) and Gompertz (b) equations.
Figure 3.
Experimental values (points) and mathematical modelling (continuous lines) of absorbance measurements at 630 nm as a function of time for the Saccharomyces cerevisiae (SF-04) and Saccharomyces cerevisiae var. boulardıi (SCb) strains cultured in batch bioreactor in anaerobic conditions at 24 ± 1 °C. Mathematical modelling was performed with the logistic (a) and Gompertz (b) equations.

Figure 4.
Monitored pH values as a function of time for the Saccharomyces cerevisiae (SF-04) and Saccharomyces cerevisiae var. boulardii (SCb) strains in bioreactor batch culture at 24 ± 1 °C.
Figure 4.
Monitored pH values as a function of time for the Saccharomyces cerevisiae (SF-04) and Saccharomyces cerevisiae var. boulardii (SCb) strains in bioreactor batch culture at 24 ± 1 °C.

Figure 5.
Normalised cell volume (biovolume) distribution values estimated for the Saccharomyces cerevisiae var. boulardii (SCb) and Saccharomyces cerevisiae (SF-04) strains.
Figure 5.
Normalised cell volume (biovolume) distribution values estimated for the Saccharomyces cerevisiae var. boulardii (SCb) and Saccharomyces cerevisiae (SF-04) strains.

Figure 6.
Yeast viability (a) of the Saccharomyces cerevisiae var. boulardii (Scb) and Saccharomyces cerevisiae (SF-04) strains in the craft beers (b) determined by the colony counting method after 45 days. The number of colonies, which are the number of viable microorganisms (expressed in CFU/mL) in the craft beers and representative plate images of the viable yeast colonies with a dilution factor 10−2 and 10−3 for SF-04 and SCb, respectively, are shown.
Figure 6.
Yeast viability (a) of the Saccharomyces cerevisiae var. boulardii (Scb) and Saccharomyces cerevisiae (SF-04) strains in the craft beers (b) determined by the colony counting method after 45 days. The number of colonies, which are the number of viable microorganisms (expressed in CFU/mL) in the craft beers and representative plate images of the viable yeast colonies with a dilution factor 10−2 and 10−3 for SF-04 and SCb, respectively, are shown.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Retention times and wavelengths for the polyphenols standard patterns.
| Polyphenol | Retention Time (min) | Wavelength (nm) |
|---|---|---|
| Phloroglucinol | 4.01 | 270 |
| Catequin | 12.66 | |
| Vanillic acid | 14.29 | |
| Epicatechin | 15.14 | |
| Protocatechuic acid | 18.49 | |
| Rutin | 19.20 | |
| Gentisic acid | 11.86 | 324 |
| Chlorogenic acid | 13.81 | |
| Caffeic acid | 14.63 | |
| p-coumaric acid | 16.84 | |
| Ferulic acid | 17.41 | |
| Myricetin | 20.48 | 373 |
| Quercetin | 24.18 |
Table 2.
Total phenolic content (TPC) determined by the Folin–Ciocalteu method and expressed in mg of gallic acid (GA) per L, antioxidant activity (RSA) obtained by the stable free radical 2,2-Diphenyl-1-picrylhydrazyl (DPPH) technique and expressed in % by Equation (1), wort density (ρwort), beer density (ρbeer), alcohol percentage by weight (ABWt) and by volume (ABV) of the craft beer fermented with Saccharomyces cerevisiae (Sc) and Saccharomyces cerevisiae var. boulardii (SF-04) yeast strains. Results of TPC and RSA shown as mean ± standard deviation. * Statistically significant differences (p < 0.05).
Table 2.
Total phenolic content (TPC) determined by the Folin–Ciocalteu method and expressed in mg of gallic acid (GA) per L, antioxidant activity (RSA) obtained by the stable free radical 2,2-Diphenyl-1-picrylhydrazyl (DPPH) technique and expressed in % by Equation (1), wort density (ρwort), beer density (ρbeer), alcohol percentage by weight (ABWt) and by volume (ABV) of the craft beer fermented with Saccharomyces cerevisiae (Sc) and Saccharomyces cerevisiae var. boulardii (SF-04) yeast strains. Results of TPC and RSA shown as mean ± standard deviation. * Statistically significant differences (p < 0.05).
| Yeast Strain | TPC (mg GA/L) | RSA (%) | ρwort (kg/m3) | ρbeer (kg/m3) | ABWt (%) | ABV (%) |
|---|---|---|---|---|---|---|
| SF-04 | 0.1597 ± 0.0373 | 11.51 ± 0.36 | 1028 | 1008 | 1.87 | 2.39 |
| Scb | 0.1545 ± 0.0192 | 16.80 ± 0.31 * | 1028 | 1014 | 1.29 | 1.65 |
Table 3.
Parameter estimates of k, a, r and their errors for the logistic and Gompertz models applied to the absorbance measurements at 630 nm for the Saccharomyces cerevisiae (SF-04) and Saccharomyces cerevisiae var. boulardii (SCb) strains cultured in batch bioreactors at 24 ± 1 °C for 50 h. All these results are statistically significant (p < 0.05).
Table 3.
Parameter estimates of k, a, r and their errors for the logistic and Gompertz models applied to the absorbance measurements at 630 nm for the Saccharomyces cerevisiae (SF-04) and Saccharomyces cerevisiae var. boulardii (SCb) strains cultured in batch bioreactors at 24 ± 1 °C for 50 h. All these results are statistically significant (p < 0.05).
| Yeast Strain | K | Error | a | Error | r | Error | Model |
|---|---|---|---|---|---|---|---|
| SF-04 | 1.832 | 0.066 | 35.858 | 0.799 | 0.113 | 0.005 | Logistic |
| Scb | 1.644 | 0.008 | 23.092 | 0.093 | 0.302 | 0.007 | |
| SF-04 | 2.719 | 0.143 | 5.960 | 0.179 | 0.047 | 0.002 | Gompertz |
| Scb | 1.671 | 0.016 | 69.873 | 14.079 | 0.203 | 0.009 |
Table 4.
Sensory evaluation results for the beers produced with the Saccharomyces cerevisiae (SF-04) and Saccharomyces cerevisiae var. boulardii (Scb) strains. Scale from 1 (extremely dislike attribute) to 9 (extremely like attribute). Results expressed as mean ± standard deviation. No significant differences (p < 0.05) between row and column values were found.
Table 4.
Sensory evaluation results for the beers produced with the Saccharomyces cerevisiae (SF-04) and Saccharomyces cerevisiae var. boulardii (Scb) strains. Scale from 1 (extremely dislike attribute) to 9 (extremely like attribute). Results expressed as mean ± standard deviation. No significant differences (p < 0.05) between row and column values were found.
| Yeast Strain | Appearance | Aroma | Flavor | Bitterness |
|---|---|---|---|---|
| SF-04 | 6.25 ± 1.03 | 6.12 ± 0.99 | 5.87 ± 1.13 | 4.75 ± 1.04 |
| Scb | 6.50 ± 1.41 | 5.62 ± 1.51 | 5.37 ± 1.30 | 5.25 ± 1.58 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mulero-Cerezo, J.; Briz-Redón, Á.; Serrano-Aroca, Á. Saccharomyces Cerevisiae Var. Boulardii: Valuable Probiotic Starter for Craft Beer Production. Appl. Sci. 2019, 9, 3250. https://0-doi-org.brum.beds.ac.uk/10.3390/app9163250
AMA Style
Mulero-Cerezo J, Briz-Redón Á, Serrano-Aroca Á. Saccharomyces Cerevisiae Var. Boulardii: Valuable Probiotic Starter for Craft Beer Production. Applied Sciences. 2019; 9(16):3250. https://0-doi-org.brum.beds.ac.uk/10.3390/app9163250
Chicago/Turabian StyleMulero-Cerezo, Joaquín, Álvaro Briz-Redón, and Ángel Serrano-Aroca. 2019. "Saccharomyces Cerevisiae Var. Boulardii: Valuable Probiotic Starter for Craft Beer Production" Applied Sciences 9, no. 16: 3250. https://0-doi-org.brum.beds.ac.uk/10.3390/app9163250
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.