
Coconut (Cocos nucifera) Ethanolic Leaf Extract Reduces Amyloid-β (1-42) Aggregation and Paralysis Prevalence in Transgenic Caenorhabditis elegans Independently of Free Radical Scavenging and Acetylcholinesterase Inhibition

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Leaf Harvesting and Crude Ethanolic Extraction of Cocos nucifera Leaves
2.2. Caenorhabditis elegans Strains
- N2—wild type
- C. elegans var. Bristol
- CL4176—expresses Aβ1–42
- dvIs27 [myo-3p::A-β(1-42)::let-851 3′UTR]
- CL2006—expresses Aβ1–42; temperature-sensitive
- dvIs2 [pCL12(unc-54/human Aβ peptide 1-42 minigene) + pRF4]
2.3. Antioxidant and Acetylcholinesterase Inhibitory Activities of Crude CLE Extracts
2.4. Nematode Toxicity Assay
2.5. Amyloid-β Aggregation and Paralysis Tests
2.6. Statistical Analyses
3. Results
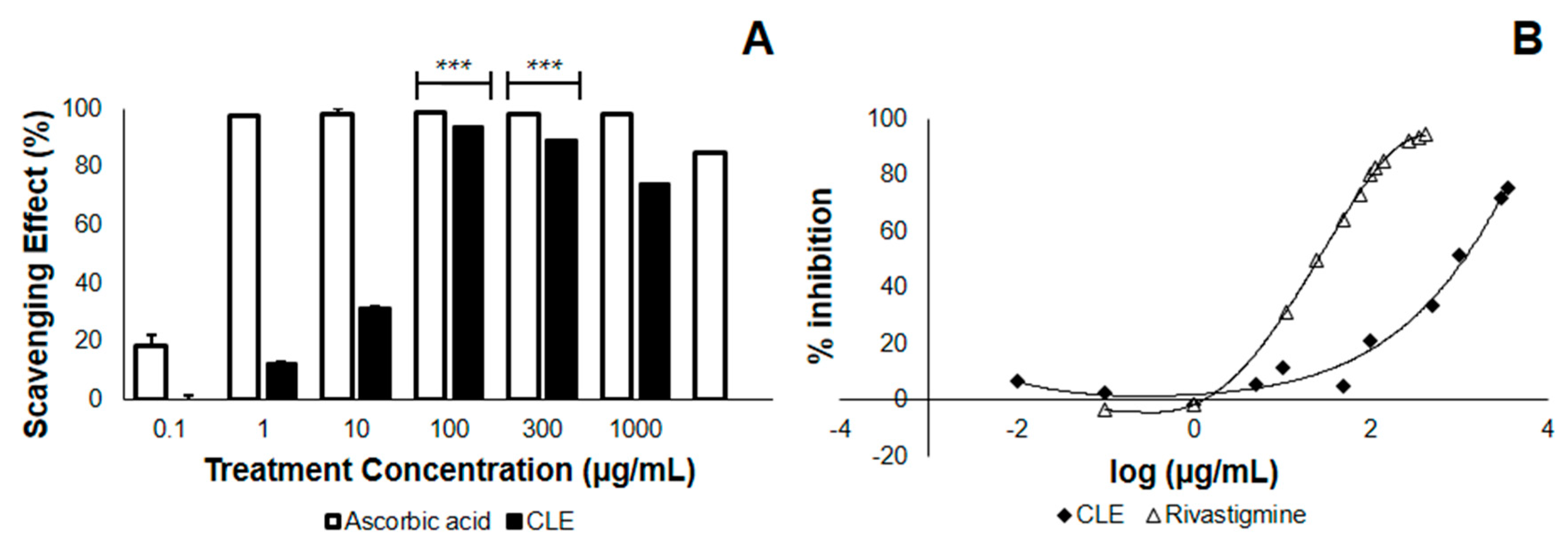
3.1. Coconut Leaf Extract (CLE) Neutralizes DPPH Radicals in a Dose-Dependent Manner
3.2. CLE Inhibits AChE Less Effectively than Does Rivastigmine In Vitro
3.3. CLE is Non-Lethal to Wild-type and Transgenic C. elegans CL2006 and CL4176
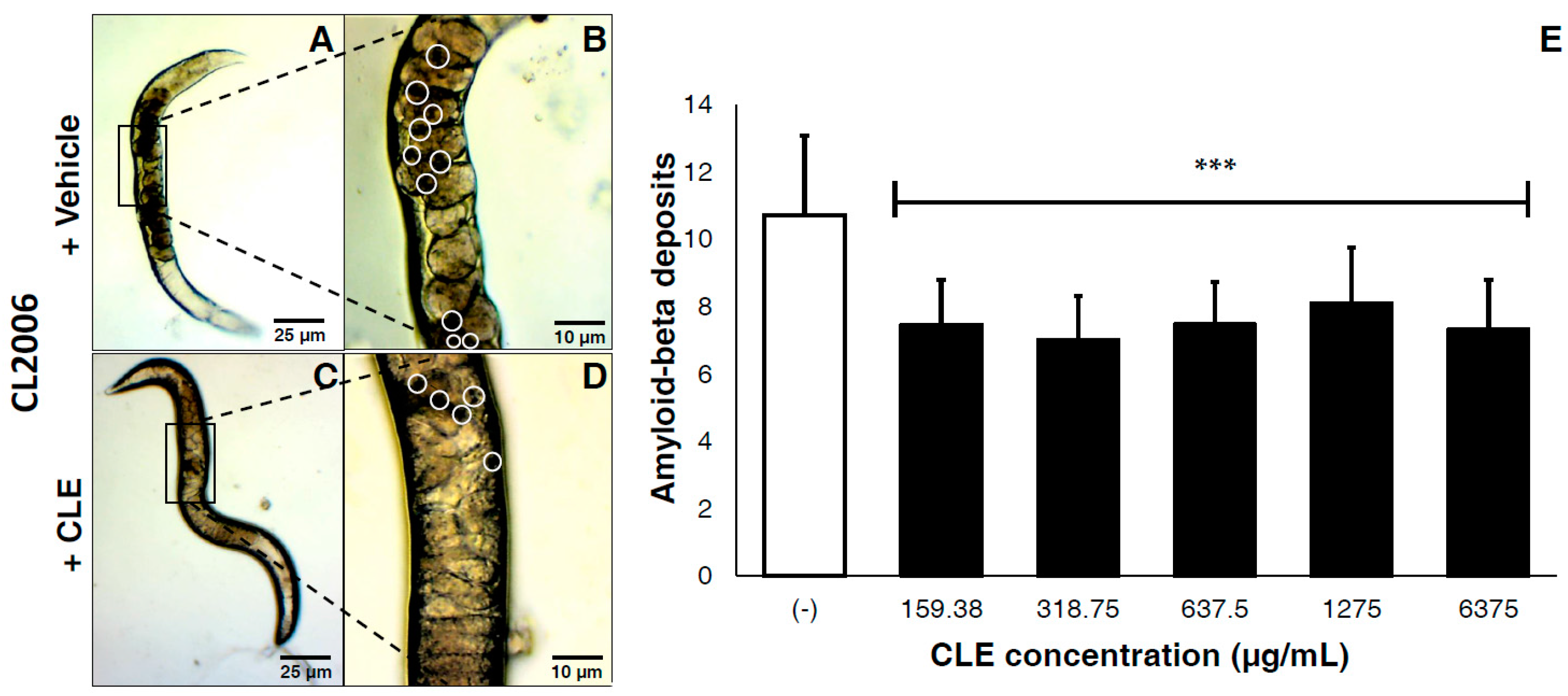
3.4. CLE Significantly Reduces Aβ Aggregate Deposits in Transgenic C. elegans Strains CL2006 and CL4176
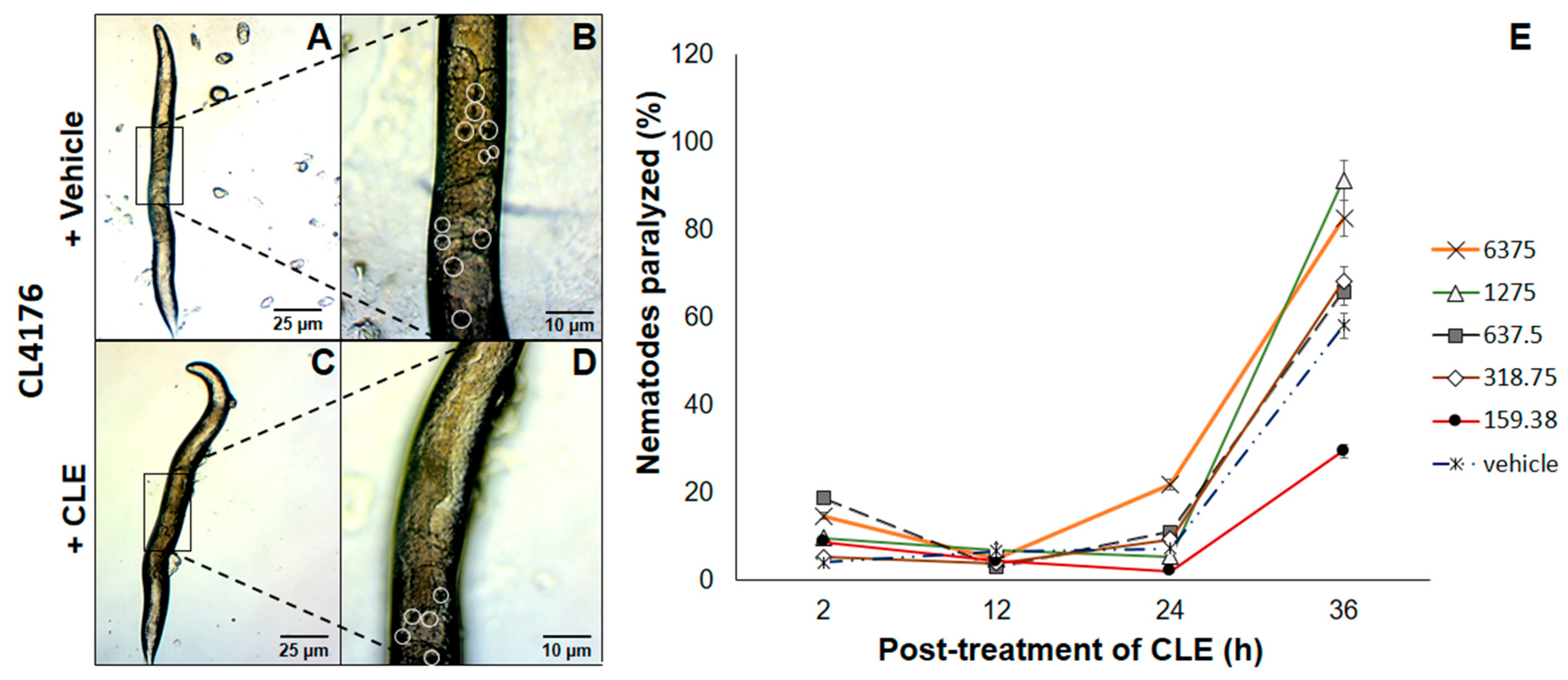
3.5. Paralysis of CL4176 C. elegans Strains Was Partially Relieved by Exposure to CLE Prior to Heat Activation of Aβ Expression
3.6. Phytochemicals Present in CLE Possibly Interact with Aβ1–42 Peptide
4. Discussion
4.1. C. elegans as a Model for Alzheimer’s Disease (AD) and Sporadic Inclusion Body Myositis (sIBM)
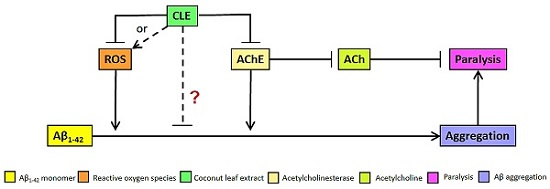
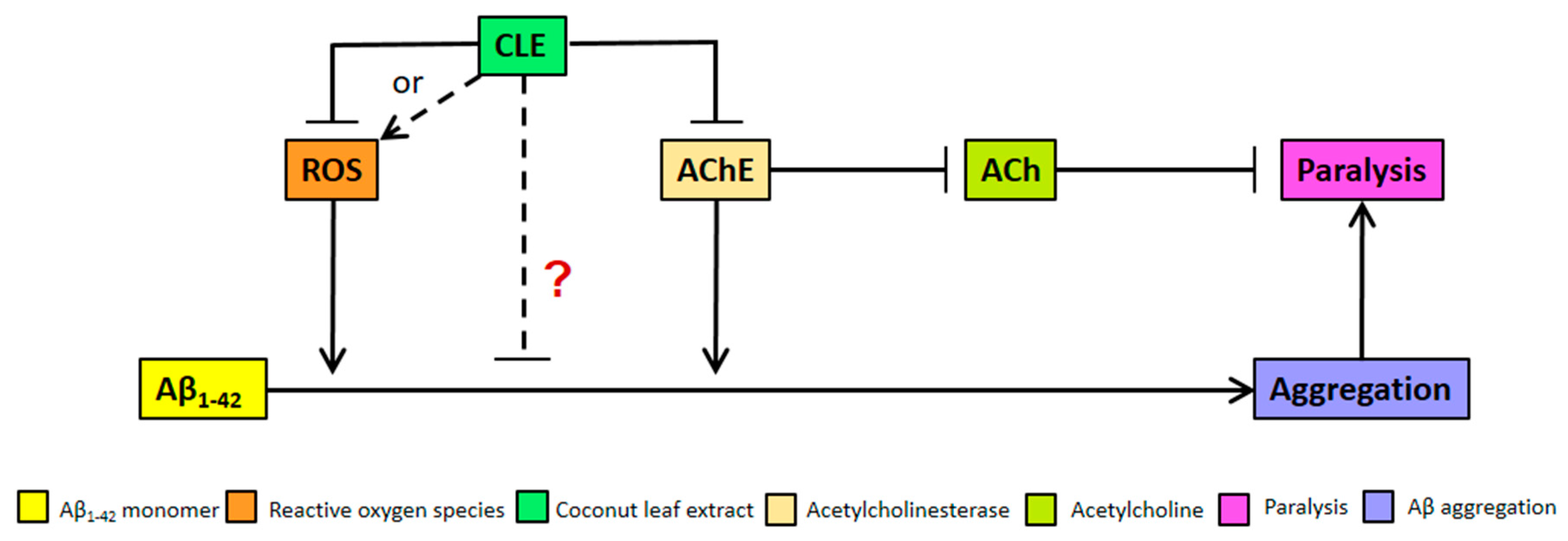
4.2. Reduction of Aβ1–42 Aggregation by CLE Is Independent of Free Radical Scavenging and AChE Inhibition
4.3. Reduction of Aβ-Induced Paralysis by CLE Is Independent of Free Radical Scavenging and AChE Inhibition
4.4. Anthocyanins, Tannins and Glycosides Are Candidate Compounds Against Aβ1–42 Induced Pathology
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- El Gaamouch, F.; Jing, P.; Xia, J.; Cai, D. Alzheimer’s Disease Risk Genes and Lipid Regulators. J. Alzheimer’s Dis. 2016, 53, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.L.; Narasimhan, S.; Changolkar, L.; He, Z.; Stieber, A.; Zhang, B.; Gathagan, R.J.; Iba, M.; McBride, J.D.; Trojanowski, J.Q.; et al. Unique pathological tau conformers from Alzheimer’s brains transmit tau pathology in nontransgenic mice. J. Exp. Med. 2016, 213, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Nilson, A.N.; English, K.C.; Gerson, J.E.; Whittle, T.B.; Crain, C.N.; Xue, J.; Sengupta, U.; Castillo-Carranza, D.L.; Zhang, W.; Gupta, P.; et al. Tau Oligomers Associate with Inflammation in the Brain and Retina of Tauopathy Mice and in Neurodegenerative Diseases. J. Alzheimer’s Dis. 2016, 55, 1083–1099. [Google Scholar] [CrossRef] [PubMed]
- Crary, J. Primary age-related tauopathy and the amyloid cascade hypothesis: The exception that proves the rule. J. Neurol. Neuromed. 2016, 1, 53–57. [Google Scholar]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Laurent, C.; Dorotheé, G.; Hunot, S.; Martin, E.; Monnet, Y.; Duchamp, M.; Dong, Y.; Légeron, F.P.; Leboucher, A.; Burnouf, S.; et al. Hippocampal T cell infiltration promotes neuroinflammation and cognitive decline in a mouse model of tauopathy. Brain 2016, 140, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Tardivel, M.; Bégard, S.; Bousset, L.; Dujardin, S.; Coens, A.; Melki, R.; Buée, L.; Colin, M. Tunneling nanotube (TNT)-mediated neuron-to neuron transfer of pathological Tau protein assemblies. Acta Neuropathol. Commun. 2016, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Karpati, G.; O’Ferrall, E. Sporadic inclusion body myositis: Pathogenic considerations. Ann. Neurol. 2009, 65, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Dalakas, M.C. Sporadic inclusion body myositis—Diagnosis, pathogenesis and therapeutic strategies. Nat. Clin. Pract. Neurol. 2006, 2, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.H.; Brumfield, K.A.; Kasperbauer, J.L.; Basford, J.R. Dysphagia in inclusion body myositis: Clinical features, management, and clinical outcome. Am. J. Phys. Med. Rehabil. 2008, 87, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; de la Rubia Orti, J.E.; Selvi, S.; Sancho Castillo, S.; Rochina, M.J.; Manresa Ramon, R.; Montoya-Castilla, I. Coconut Oil: Non-Alternative Drug Treatment Against Alzheimer’s Disease. Nutr. Hosp. 2015, 32, 2822–2827. [Google Scholar]
- Nafar, F.; Mearow, K.M. Coconut oil attenuates the effects of amyloid-β on cortical neurons in vitro. J. Alzheimer’s Dis. 2014, 39, 233–237. [Google Scholar]
- Nonaka, Y.; Takagi, T.; Inai, M.; Nishimura, S.; Urashima, S.; Honda, K.; Aoyama, T.; Terada, S. Lauric Acid Stimulates Ketone Body Production in the KT-5 Astrocyte Cell Line. J. Oleo Sci. 2016, 65, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Dedea, L. Can coconut oil replace caprylidene for Alzheimer disease? J. Am. Acad. Phys. Assist. 2012, 25, 19. [Google Scholar] [CrossRef]
- Fernando, W.M.; Martins, I.J.; Goozee, K.G.; Brennan, C.S.; Jayasena, V.; Martins, R.N. The role of dietary coconut for the prevention and treatment of Alzheimer’s disease: Potential mechanisms of action. Br. J. Nutr. 2015, 114, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.W.C.; Lim, Y.Y.; Omar, M. Antioxidant and antibacterial activity of leaves of Etlingera species (Zingiberaceae) in Peninsular Malaysia. Food Chem. 2007, 104, 1586–1593. [Google Scholar] [CrossRef]
- Stiernagle, T. Maintenance of C. elegans. Available online: http://www.wormbook.org/chapters/www_strainmaintain/strainmaintain.html (accessed on 20 December 2016).
- Lefèvre, G.; Callegari, F.; Gstegier, S.; Xiong, Y. Effects of Renal Impairment on Steady-State Plasma Concentrations of Rivastigmine: A Population Pharmacokinetic Analysis of Capsule and Patch Formulations in Patients with Alzheimer’s Disease. Drugs Aging 2016, 33, 725–736. [Google Scholar] [CrossRef] [PubMed]
- George, L.; Ellman, K.; Courtney, D.; Andres, V., Jr.; Featherstone, R. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 90–95. [Google Scholar]
- Qiao, Y.; Zhao, Y.; Wu, Q.; Sun, L.; Ruan, Q.; Chen, Y.; Wang, M.; Duan, J.; Wang, D. Full toxicity assessment of Genkwa Flos and the underlying mechanism in nematode Caenorhabditis elegans. PLoS ONE 2014, 9, e91825. [Google Scholar] [CrossRef] [PubMed]
- Lublin, A.L.; Link, C.D. Alzheimer’s Disease Drug Discovery: In vivo screening using C. elegans as a model for β-amyloid peptide-induced toxicity. Drug Discov. Today Technol. 2013, 10, e115–e119. [Google Scholar] [CrossRef] [PubMed]
- Ryu, D.; Mouchiroud, L.; Andreux, P.A.; Katsyuba, E.; Moullan, N.; Nicolet-Dit-Felix, A.A.; Williams, E.G.; Jha, P.; Lo Sasso, G.; Huzard, D.; et al. Urolithin A induces mitophagy and prolongs lifespan in C. elegans and increases muscle function in rodents. Nat. Med. 2016, 22, 879–888. [Google Scholar] [PubMed]
- Valero, T. Mitochondrial biogenesis: Pharmacological approaches. Curr. Pharm. Des. 2014, 20, 5507–5509. [Google Scholar] [CrossRef] [PubMed]
- Tomas-Barberan, F.A.; Gonzales-Sarrias, A.; Garcia-Villalba, R.; Nunez-Sanchez, M.A.; Selma, M.V.; Garcia-Conesa, M.T.; Espin, J.C. Urolithins, the rescue of ‘old’ metabolites to understand a ‘new’ concept: Metabotypes as a nexus between phenolic metabolism, microbiota dysbiosis and host health status. Mol. Nutr. Food Res. 2016, 61, 1500901. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Aronson, W.J.; Zhang, Y.; Henning, S.M.; Moro, A.; Lee, R.P.; Sartippour, M.; Harris, D.M.; Rettig, M.; Suchard, M.A.; et al. Pomegrenate ellagitannin-derived metabolites inhibit prostate cancer growth and localize to the mouse prostate gland. J. Agric. Food Chem. 2007, 55, 7732–7737. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, M.T.; Farbood, Y.; Sameri, M.J.; Sarkaki, A.; Naghizadeh, B.; Rafeirad, M. Neuroprotective effects of oral gallic acid against oxidative stress induced by 6-hydroxydopamine in rats. Food Chem. 2013, 138, 1028–1033. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.X.; Li, Y.B.; Zhao, R.P. Epigallocatechin Gallate Attenuates β-Amyloid Generation and Oxidative Stress Involvement of PPARγ in N2a/APP695 Cells. Neurochem. Res. 2017, 42, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Gul, F.; Khan, K.M.; Adhikari, A.; Zafar, S.; Akram, M.; Khan, H.; Saeed, M. Antimicrobial and antioxidant activities of a new metabolite from Quercus incana. Nat. Prod. Res. 2016, 21, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.Y.; Bae, K.; Seong, Y.H.; Song, K.S. Green tea catechins as a BACE1 (β-secretase) inhibitor. Bioorg. Med. Chem. Lett. 2003, 13, 3905–3908. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.; Kantham, S.; Rao, V.M.; Palanivelu, M.K.; Pham, H.L.; Shaw, P.N.; McGeary, R.P.; Ross, B.P. Metal chelation radical scavenging and inhibition of Aβ42 fibrillation by food constituents in relation to Alzheimer’s disease. Food Chem. 2016, 199, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Smith, T. Pharmakokinetics, bioavailability and serum levels of cardiac glycosides. J. Am. Coll. Cardiol. 1985, 5, 43A–50A. [Google Scholar] [CrossRef]
- Alexander, A.; Marfil, V.; Li, C. Use of Caenorhabditis elegans as a model to study Alzheimer’s disease and other neurodegenerative diseases. Front. Genet. 2014, 5, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Diomede, L.; Rigacci, S.; Romeo, M.; Stefani, M.; Salmona, M. Oleuropein Aglycone Protects Transgenic C. elegans Strains Expressing Aβ42 by Reducing Plaque Load and Motor Deficit. PLoS ONE 2013, 8, e58893. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Barclay, J.; Burgoyne, R.; Morgan, A. Using C. elegans to discover therapeutic compounds for ageing-associated neurodegenerative diseases. Chem. Cent. J. 2015, 9, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Markaki, M.; Tavernarakis, N. Modeling human diseases in Caenorhabditis elegans. Biotechnol. J. 2010, 5, 1261–1276. [Google Scholar] [CrossRef] [PubMed]
- Link, C.D. Expression of human β-amyloid peptide in transgenic Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 1995, 92, 9368–9372. [Google Scholar] [CrossRef] [PubMed]
- David, D.; Ollikainen, N.; Trinidad, J.; Cary, M.; Burlingame, A.; Kenyon, C. Widespread Protein Aggregation as an Inherent Part of Aging in C. elegans. PLoS Biol. 2010, 8, e1000450. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.H.; Hyon-Lee; Lee, K.M. The Possible Role of Antioxidant Vitamin C in Alzheimer’s Disease Treatment and Prevention. Am. J. Alzheimer’s Dis. 2013, 28, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Mukherjee, A.; Swarnakar, S.; Das, N. Nanocapsulated Ascorbic Acid Combating Cerebral Ischemia Reperfusion-Induced Oxidative Injury in Rat Brain. Curr. Alzheimer Res. 2016, 13, 1363–1373. [Google Scholar] [CrossRef] [PubMed]
- Sil, S.; Ghosh, T.; Gupta, P.; Ghosh, R.; Kabir, S.N.; Roy, A. Dual Role of Vitamin C on the Neuroinflammation Mediated Neurodegeneration and Memory Impairments in Colchicine Induced Rat Model of Alzheimer’s Disease. J. Mol. Neurosci. 2016, 60, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Rietjens, I.; Boersma, M.; de Haan, L.; Spenkelink, B.; Awad, H.; Cnubben, N.; van Zanden, J.; van der Woude, H.; Alink, G.; Koeman, J. The pro-oxidant chemistry of the natural antioxidants vitamin C, vitamin E, carotenoids and flavonoids. Environ. Toxicol. Pharmacol. 2002, 11, 321–333. [Google Scholar] [CrossRef]
- Putchala, M.C.; Ramani, P.; Sherlin, H.J.; Premkumar, P.; Natesan, A. Ascorbic acid and its pro-oxidant activity as a therapy for tumours of oral cavity—A systematic review. Arch. Oral. Biol. 2013, 58, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Ginotra, Y.P.; Ramteke, S.N.; Walke, G.R.; Rapole, S.; Kulkarni, P.P. Histidine availability is decisive in ROS-mediated cytotoxicity of copper complexes of Aβ1-16 peptide. Free Radic. Res. 2016, 50, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, K.; Wong, J.M.; Vann, P.H.; Sumien, N. Exercise, but not antioxidants, reversed the ApoE4-associated motor impairments in adult GFAP-ApoE mice. Behav. Brain Res. 2016, 305, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Ohlow, M.J.; Sohre, S.; Granold, M.; Schreckenberger, M.; Moosmann, B. Why Have Clinical Trials of Antioxidants to Prevent Neurodegeneration Failed?—A Cellular Investigation of Novel Phenothiazine-Type Antioxidants Reveals Competing Objectives for Pharmaceutical Neuroprotection. Pharm. Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-K.; Kim, T.H.; Lee, S.-J. Mechanisms of aging-related proteinopathies in Caenorhabditis elegans. Exp. Mol. Med. 2016, 48, e263. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, W.; Ebert, P.R. Metformin Attenuates Aβ Pathology Mediated Through Levamisole Sensitive Nicotinic Acetylcholine Receptors in a C. elegans Model of Alzheimer’s Disease. Mol. Neurobiol. 2016, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kikis, E. The struggle by Caenorhabditis elegans to maintain proteostasis during aging and disease. Biol. Direct 2016, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- Cuanalo-Contreras, K.; Park, W.; Murkherjee, A.; Millán-Pérez, P.; Soto, C. Delaying aging in Caenorhabditis elegans with protein aggregation inhibitors. Biochem. Biophys. Res. Commun. 2017, 482, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, X.; Zhang, X.; Jin, Q.; Li, J. Phenolic Profiles, Antioxidant Activities, and Neuroprotective Properties of Mulberry (Morus atropurpurea Roxb.) Fruit Extracts from Different Ripening Stages. J. Food Sci. 2016, 81, C2439–C2446. [Google Scholar] [CrossRef] [PubMed]
- Di Giovanni, S.; Eleuteri, S.; Paleologou, K.E.; Yin, G.; Zweckstetter, M.; Carrupt, P.A.; Lashuel, H.A. Entacapone and tolcapone, two catechol O-methyltransferase inhibitors, block fibril formation of α-synuclein and β-amyloid and protect against amyloid-induced toxicity. J. Biol. Chem. 2010, 285, 14941–14954. [Google Scholar] [CrossRef] [PubMed]
- Du, X.T.; Wang, L.; Wang, Y.J.; Andreasen, M.; Zhan, D.W.; Feng, Y.; Li, M.; Zhao, M.; Otzen, D.; Xue, D.; et al. Aβ1-16 can aggregate and induce the production of reactive oxygen species, nitric oxide, and inflammatory cytokines. J. Alzheimer’s Dis. 2011, 27, 401–413. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group | CLE Concentration (μg/mL) | C. elegans Strain | Temperature |
|---|---|---|---|
| Aβ1–42 Aggregation Assay | |||
| 1 | (−) | CL2006 | 20 °C |
| 2 | 159.38 | ||
| 3 | 318.75 | ||
| 4 | 637.5 | ||
| 5 | 1275 | ||
| 6 | 6375 | ||
| Aβ1–42 Paralysis Assay | |||
| 1 | (−) | CL4176 | Pre-treatment period: 10–16 °C Post-treatment period: 24 °C |
| 2 | 159.38 | ||
| 3 | 318.75 | ||
| 4 | 637.5 | ||
| 5 | 1275 | ||
| 6 | 6375 | ||
| Treatment Group | Concentration (μg/mL) | EC50/IC50 (μg/mL) | Positive Control | |
|---|---|---|---|---|
| DPPH Scavenging Assay | ||||
| CLE | Control | Ascorbic acid | ||
| 1 | 0.1 | 18.11 | 0.23 | |
| 2 | 1 | |||
| 3 | 10 | |||
| 4 | 100 | |||
| 5 | 300 | |||
| 6 | 1000 | |||
| AChE Inhibition Assay | ||||
| 1 | 0.01 | 3218.56 | 24.52 | Rivastigmine |
| 2 | 0.1 | |||
| 3 | 1 | |||
| 4 | 5 | |||
| 5 | 10 | |||
| 6 | 50 | |||
| 7 | 100 | |||
| 8 | 500 | |||
| 9 | 1000 | |||
| 10 | 3000 | |||
| 11 | 3500 | |||
| Test Parameter | Method | Result |
|---|---|---|
| Alkaloids | Mayer/Meyer test | Alkaloids absent |
| Anthraquinones | Bornträger test | Anthraquinones absent |
| Cardenolides and bufadienolides | Keller-Kiliani test | Glycoside compounds present |
| Flavonoids | Bate-Smith and Metcalf test | Anthocyanins present |
| Tannins and polyphenolic compounds | Ferric chloride test | Hydrolyzable tannins present |
| Saponins | Froth test | Saponins absent |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manalo, R.V.; Silvestre, M.A.; Barbosa, A.L.A.; Medina, P.M. Coconut (Cocos nucifera) Ethanolic Leaf Extract Reduces Amyloid-β (1-42) Aggregation and Paralysis Prevalence in Transgenic Caenorhabditis elegans Independently of Free Radical Scavenging and Acetylcholinesterase Inhibition. Biomedicines 2017, 5, 17. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines5020017
Manalo RV, Silvestre MA, Barbosa ALA, Medina PM. Coconut (Cocos nucifera) Ethanolic Leaf Extract Reduces Amyloid-β (1-42) Aggregation and Paralysis Prevalence in Transgenic Caenorhabditis elegans Independently of Free Radical Scavenging and Acetylcholinesterase Inhibition. Biomedicines. 2017; 5(2):17. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines5020017
Chicago/Turabian StyleManalo, Rafael Vincent, Maries Ann Silvestre, Aza Lea Anne Barbosa, and Paul Mark Medina. 2017. "Coconut (Cocos nucifera) Ethanolic Leaf Extract Reduces Amyloid-β (1-42) Aggregation and Paralysis Prevalence in Transgenic Caenorhabditis elegans Independently of Free Radical Scavenging and Acetylcholinesterase Inhibition" Biomedicines 5, no. 2: 17. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines5020017