P120 Catenin Isoforms Differentially Associate with Breast Cancer Invasion and Metastasis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Antibodies and Dyes
2.3. Subcellular Protein Fractionation
2.4. Cytoskeleton Extraction
2.5. Co-Immunoprecipitation with p120 Isoform-Specific Antibodies
2.6. Western Blot Analysis
2.7. Immunofluorescence of Cultured Cells
2.8. Gap Closure Assay
2.9. Spheroid Invasion in 3D Culture
2.10. In Silico Splicing Analysis of Human Breast Cancer RNA-seq Data
2.11. P120 Isoform Expression in Human Breast Cancer Samples
2.12. H&E and Immunofluorescence of Histological Sections
2.13. Analysis of p120 Isoform Expression in Human Breast Cancer Sections
3. Results
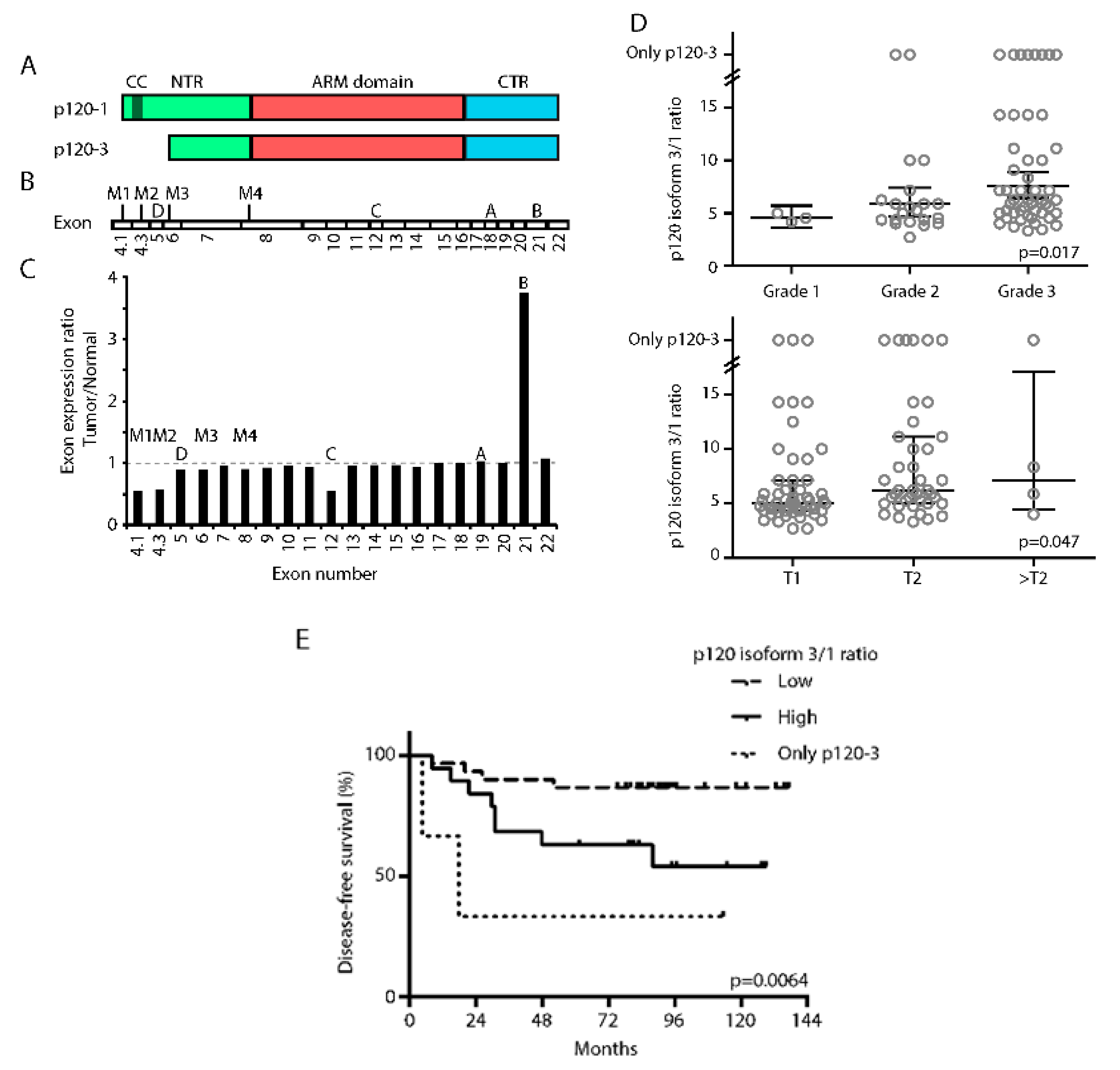
3.1. High p120-3/p120-1 Isoform Expression Ratio Predicts Poor Prognosis
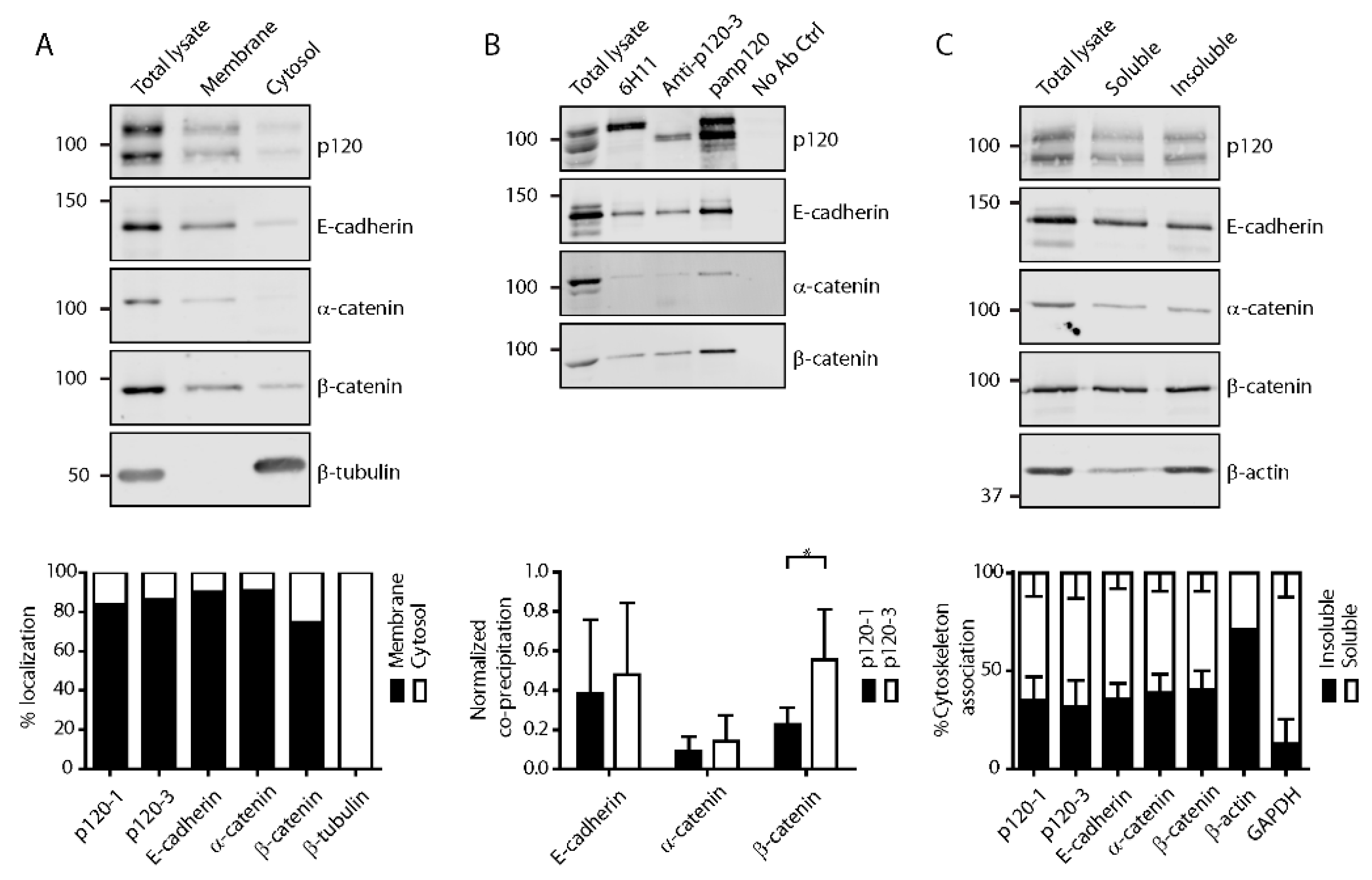
3.2. P120 Isoforms Equally Associate with Adherens Junctions in Confluent Monolayers
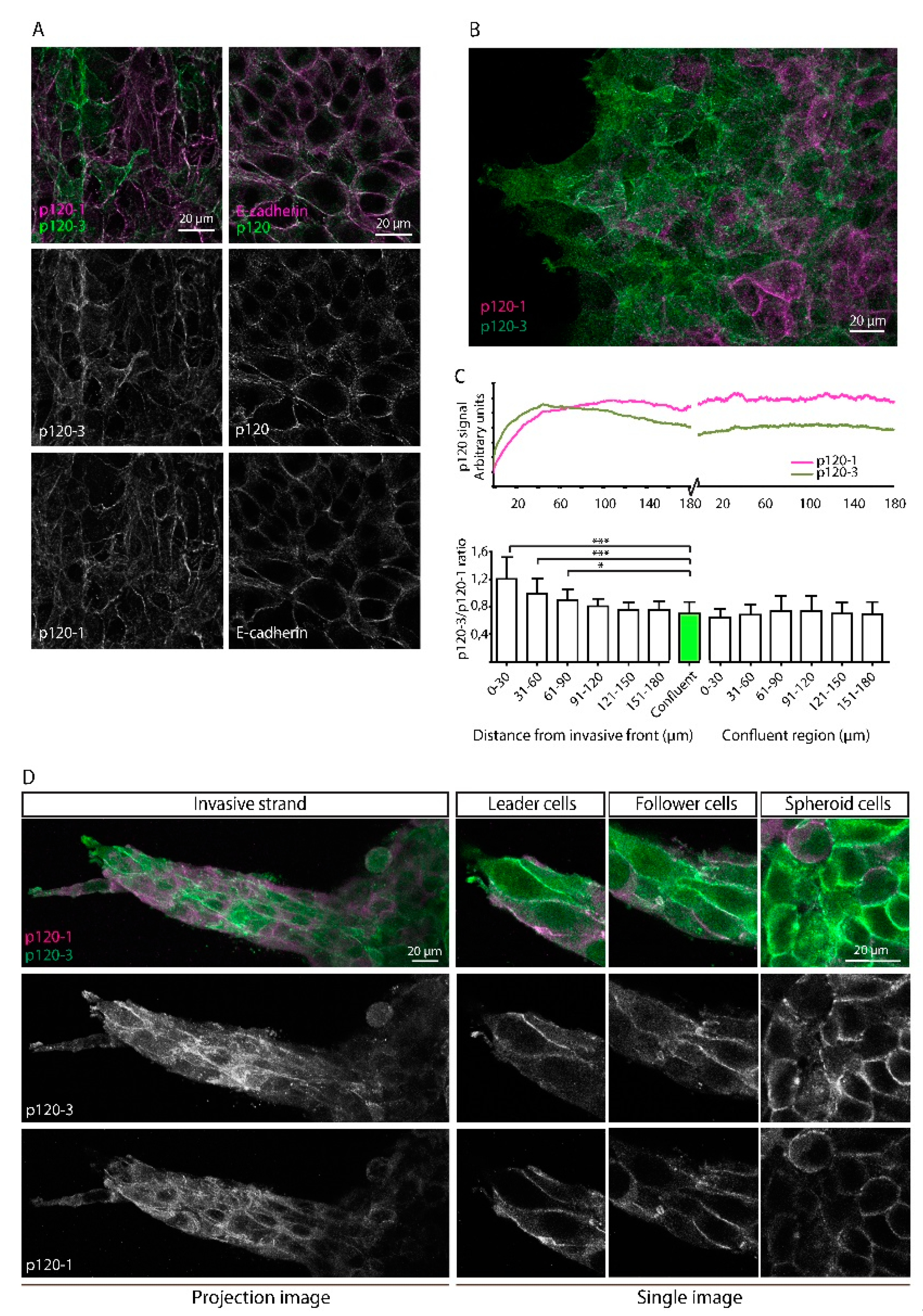
3.3. Upregulation of p120-3 at the Invasive Front of Moving Multicellular Sheets
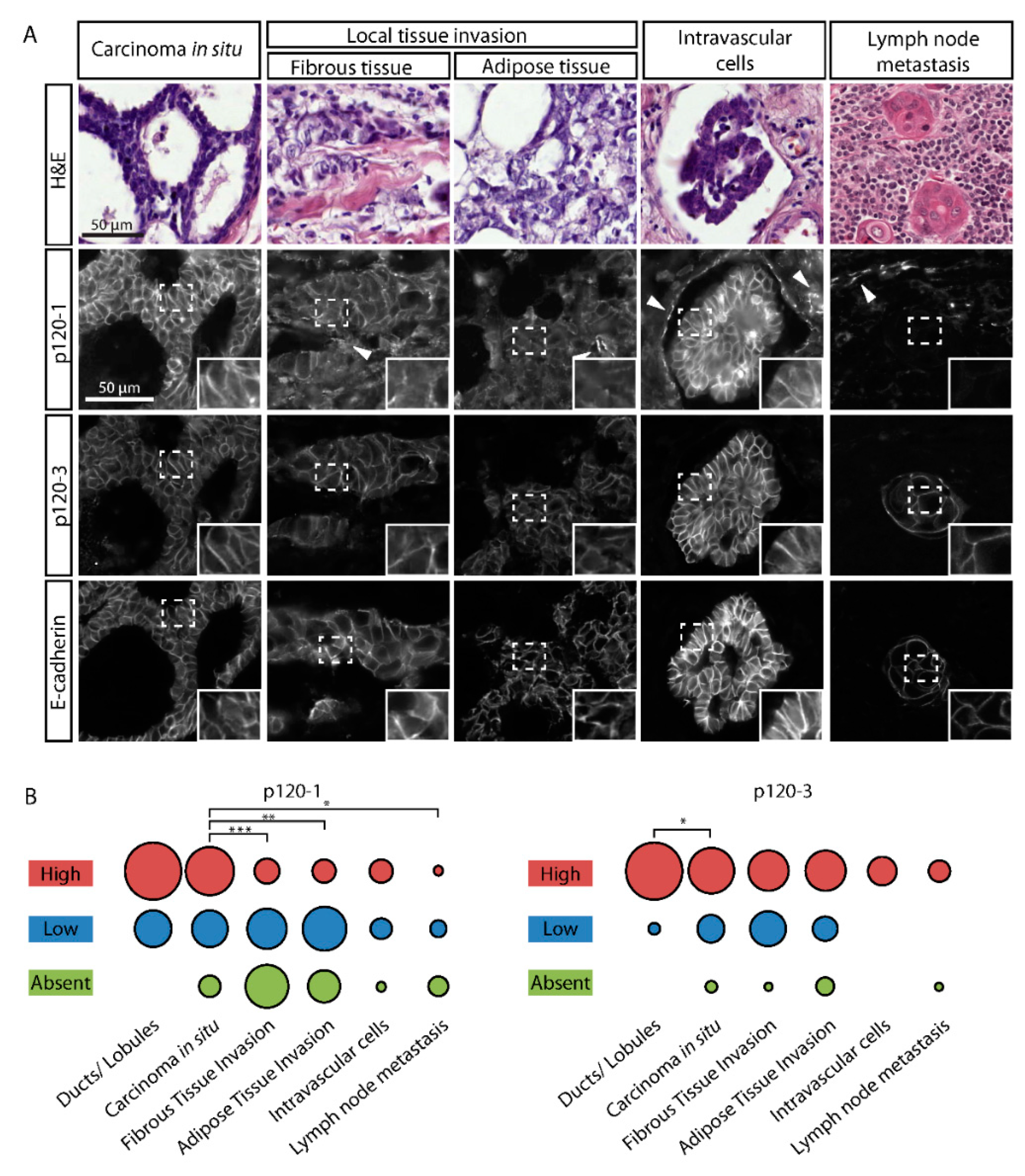
3.4. P120-3 Is Expressed throughout the Metastatic Cascade, Whereas p120-1 Is Reduced in Invasive Regions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davis, M.A.; Ireton, R.C.; Reynolds, A.B. A core function for p120-catenin in cadherin turnover. J. Cell Biol. 2003, 163, 525–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoreson, M.A.; Reynolds, A.B. Altered expression of the catenin p120 in human cancer: Implications for tumor progression. Differentiation 2002, 70, 583–589. [Google Scholar] [CrossRef]
- Talvinen, K.; Tuikkala, J.; Nykanen, M.; Nieminen, A.; Anttinen, J.; Nevalainen, O.S.; Hurme, S.; Kuopio, T.; Kronqvist, P. Altered expression of p120catenin predicts poor outcome in invasive breast cancer. J. Cancer Res. Clin. Oncol. 2010, 136, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Kokubu, A.; Sekine, S.; Kanai, Y.; Hirohashi, S. Cytoplasmic p120ctn regulates the invasive phenotypes of E-cadherin-deficient breast cancer. Am. J. Pathol. 2004, 164, 2269–2278. [Google Scholar] [CrossRef]
- Chen, T.; Wang, C.; Wu, F.; Zhang, X.; Yang, H.; Deng, X.; He, Q.; Li, W.; Li, G. Altered localization of p120 catenin in the cytoplasm rather than the membrane correlates with poor prognosis in esophageal squamous cell carcinoma. PLoS ONE 2015, 10, e0118645. [Google Scholar] [CrossRef]
- Sarrio, D.; Perez-Mies, B.; Hardisson, D.; Moreno-Bueno, G.; Suarez, A.; Cano, A.; Martin-Perez, J.; Gamallo, C.; Palacios, J. Cytoplasmic localization of p120ctn and E-cadherin loss characterize lobular breast carcinoma from preinvasive to metastatic lesions. Oncogene 2004, 23, 3272–3283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaya, K.; Sudo, H.; Maeda, G.; Kawashiri, S.; Imai, K. Concomitant loss of p120-catenin and beta-catenin membrane expression and oral carcinoma progression with E-cadherin reduction. PLoS ONE 2013, 8, e69777. [Google Scholar] [CrossRef]
- Ye, X.; Tam, W.L.; Shibue, T.; Kaygusuz, Y.; Reinhardt, F.; Ng Eaton, E.; Weinberg, R.A. Distinct EMT programs control normal mammary stem cells and tumour-initiating cells. Nature 2015, 525, 256–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, T.N.D.; Perez White, B.E.; Zhao, H.; Mortazavi, F.; Tonetti, D.A. Protein kinase C alpha enhances migration of breast cancer cells through FOXC2-mediated repression of p120-catenin. BMC Cancer 2017, 17, 832. [Google Scholar] [CrossRef]
- Li, Z.; Yin, S.; Zhang, L.; Liu, W.; Chen, B. Prognostic value of reduced E-cadherin expression in breast cancer: A meta-analysis. Oncotarget 2017, 8, 16445–16455. [Google Scholar] [CrossRef]
- Davis, M.A.; Reynolds, A.B. Blocked acinar development, E-cadherin reduction, and intraepithelial neoplasia upon ablation of p120-catenin in the mouse salivary gland. Dev. Cell 2006, 10, 21–31. [Google Scholar] [CrossRef]
- Perez-Moreno, M.; Song, W.; Pasolli, H.A.; Williams, S.E.; Fuchs, E. Loss of p120 catenin and links to mitotic alterations, inflammation, and skin cancer. Proc. Natl. Acad. Sci. USA 2008, 105, 15399–15404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stairs, D.B.; Bayne, L.J.; Rhoades, B.; Vega, M.E.; Waldron, T.J.; Kalabis, J.; Klein-Szanto, A.; Lee, J.S.; Katz, J.P.; Diehl, J.A.; et al. Deletion of p120-catenin results in a tumor microenvironment with inflammation and cancer that establishes it as a tumor suppressor gene. Cancer Cell 2011, 19, 470–483. [Google Scholar] [CrossRef]
- Schackmann, R.C.; Klarenbeek, S.; Vlug, E.J.; Stelloo, S.; van Amersfoort, M.; Tenhagen, M.; Braumuller, T.M.; Vermeulen, J.F.; van der Groep, P.; Peeters, T.; et al. Loss of p120-catenin induces metastatic progression of breast cancer by inducing anoikis resistance and augmenting growth factor receptor signaling. Cancer Res. 2013, 73, 4937–4949. [Google Scholar] [CrossRef]
- van Hengel, J.; Van den Broeke, C.; Pieters, T.; Libbrecht, L.; Hofmann, I.; van Roy, F. Inactivation of p120 catenin in mice disturbs intrahepatic bile duct development and aggravates liver carcinogenesis. Eur. J. Cell Biol 2016, 95, 574–584. [Google Scholar] [CrossRef]
- Tenhagen, M.; Klarenbeek, S.; Braumuller, T.M.; Hofmann, I.; van der Groep, P.; Ter Hoeve, N.; van der Wall, E.; Jonkers, J.; Derksen, P.W. P120-Catenin Is Critical for the Development of Invasive Lobular Carcinoma in Mice. J. Mammary Gland Biol. Neoplasia 2016, 21, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Peglion, F.; Etienne-Manneville, S. P120catenin alteration in cancer and its role in tumour invasion. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2013, 368, 20130015. [Google Scholar] [CrossRef] [PubMed]
- Venhuizen, J.H.; Jacobs, F.J.C.; Span, P.N.; Zegers, M.M. P120 and E-cadherin: Double-edged swords in tumor metastasis. Semin. Cancer Biol. 2019. [Google Scholar] [CrossRef]
- Schackmann, R.C.; Tenhagen, M.; van de Ven, R.A.; Derksen, P.W. P120-catenin in cancer-mechanisms, models and opportunities for intervention. J. Cell Sci. 2013, 126, 3515–3525. [Google Scholar] [CrossRef]
- Matise, L.A.; Palmer, T.D.; Ashby, W.J.; Nashabi, A.; Chytil, A.; Aakre, M.; Pickup, M.W.; Gorska, A.E.; Zijlstra, A.; Moses, H.L. Lack of transforming growth factor-beta signaling promotes collective cancer cell invasion through tumor-stromal crosstalk. Breast Cancer Res. 2012, 14, R98. [Google Scholar] [CrossRef]
- Liu, X.; Yi, C.; Wen, Y.; Radhakrishnan, P.; Tremayne, J.R.; Dao, T.; Johnson, K.R.; Hollingsworth, M.A. Interactions between MUC1 and p120 catenin regulate dynamic features of cell adhesion, motility, and metastasis. Cancer Res. 2014, 74, 1609–1620. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, I.R.; Hooper, S.; Serrels, A.; McGarry, L.; Ozanne, B.W.; Harrington, K.; Frame, M.C.; Sahai, E.; Brunton, V.G. P120-catenin is required for the collective invasion of squamous cell carcinoma cells via a phosphorylation-independent mechanism. Oncogene 2007, 26, 5214–5228. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.J.; Padmanaban, V.; Silvestri, V.; Schipper, K.; Cohen, J.D.; Fairchild, A.N.; Gorin, M.A.; Verdone, J.E.; Pienta, K.J.; Bader, J.S.; et al. Polyclonal breast cancer metastases arise from collective dissemination of keratin 14-expressing tumor cell clusters. Proc. Natl. Acad. Sci. USA 2016, 113, E854–E863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bronsert, P.; Enderle-Ammour, K.; Bader, M.; Timme, S.; Kuehs, M.; Csanadi, A.; Kayser, G.; Kohler, I.; Bausch, D.; Hoeppner, J.; et al. Cancer cell invasion and EMT marker expression: A three-dimensional study of the human cancer-host interface. J. Pathol. 2014, 234, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.A.; Ilina, O.; Gritsenko, P.G.; Bult, P.; Span, P.N.; Friedl, P. Collective invasion in ductal and lobular breast cancer associates with distant metastasis. Clin. Exp. Metastasis 2017, 34, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Kahles, A.; Lehmann, K.V.; Toussaint, N.C.; Huser, M.; Stark, S.G.; Sachsenberg, T.; Stegle, O.; Kohlbacher, O.; Sander, C.; Cancer Genome Atlas Research, N.; et al. Comprehensive Analysis of Alternative Splicing Across Tumors from 8,705 Patients. Cancer Cell 2018, 34, 211–224 e216. [Google Scholar] [CrossRef] [PubMed]
- Keirsebilck, A.; Bonne, S.; Staes, K.; van Hengel, J.; Nollet, F.; Reynolds, A.; van Roy, F. Molecular cloning of the human p120ctn catenin gene (CTNND1): Expression of multiple alternatively spliced isoforms. Genomics 1998, 50, 129–146. [Google Scholar] [CrossRef]
- Montonen, O.; Aho, M.; Uitto, J.; Aho, S. Tissue distribution and cell type-specific expression of p120ctn isoforms. J. Histochem. Cytochem. 2001, 49, 1487–1496. [Google Scholar] [CrossRef] [PubMed]
- Venhuizen, J.H.; Sommer, S.; Span, P.N.; Friedl, P.; Zegers, M.M. Differential expression of p120-catenin 1 and 3 isoforms in epithelial tissues. Scientific reports 2019, 9, 90. [Google Scholar] [CrossRef]
- Yanagisawa, M.; Huveldt, D.; Kreinest, P.; Lohse, C.M.; Cheville, J.C.; Parker, A.S.; Copland, J.A.; Anastasiadis, P.Z. A p120 catenin isoform switch affects Rho activity, induces tumor cell invasion, and predicts metastatic disease. J. Biol. Chem. 2008, 283, 18344–18354. [Google Scholar] [CrossRef]
- Daniel, J.M.; Reynolds, A.B. The catenin p120(ctn) interacts with Kaiso, a novel BTB/POZ domain zinc finger transcription factor. Mol. Cell Biol. 1999, 19, 3614–3623. [Google Scholar] [CrossRef] [PubMed]
- Markham, N.O.; Doll, C.A.; Dohn, M.R.; Miller, R.K.; Yu, H.; Coffey, R.J.; McCrea, P.D.; Gamse, J.T.; Reynolds, A.B. DIPA-family coiled-coils bind conserved isoform-specific head domain of p120-catenin family: Potential roles in hydrocephalus and heterotopia. Mol. Biol. Cell 2014, 25, 2592–2603. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.X.; Wang, Y.; Liu, Y.; Jiang, G.Y.; Li, Q.C.; Wang, E.H. P120-catenin isoform 3 regulates subcellular localization of Kaiso and promotes invasion in lung cancer cells via a phosphorylation-dependent mechanism. Int. J. Oncol. 2011, 38, 1625–1635. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chen, Y.S.; Dashwood, W.M.; Li, Q.; Lohr, C.V.; Fischer, K.; Ho, E.; Williams, D.E.; Dashwood, R.H. Divergent roles of p120-catenin isoforms linked to altered cell viability, proliferation, and invasiveness in carcinogen-induced rat skin tumors. Mol. Carcinog. 2017, 56, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Ohkubo, T.; Ozawa, M. The transcription factor Snail downregulates the tight junction components independently of E-cadherin downregulation. J. Cell Sci. 2004, 117, 1675–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warzecha, C.C.; Sato, T.K.; Nabet, B.; Hogenesch, J.B.; Carstens, R.P. ESRP1 and ESRP2 are epithelial cell-type-specific regulators of FGFR2 splicing. Mol. cell 2009, 33, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, I.M.; Cheng, A.W.; Flytzanis, N.C.; Balsamo, M.; Condeelis, J.S.; Oktay, M.H.; Burge, C.B.; Gertler, F.B. An EMT-driven alternative splicing program occurs in human breast cancer and modulates cellular phenotype. PLoS Genet. 2011, 7, e1002218. [Google Scholar] [CrossRef] [PubMed]
- Warzecha, C.C.; Jiang, P.; Amirikian, K.; Dittmar, K.A.; Lu, H.; Shen, S.; Guo, W.; Xing, Y.; Carstens, R.P. An ESRP-regulated splicing programme is abrogated during the epithelial-mesenchymal transition. EMBO J. 2010, 29, 3286–3300. [Google Scholar] [CrossRef] [PubMed]
- Gottgens, E.L.; Span, P.N.; Zegers, M.M. Roles and Regulation of Epithelial Splicing Regulatory Proteins 1 and 2 in Epithelial-Mesenchymal Transition. Int. Rev. Cell Mol. Biol. 2016, 327, 163–194. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Boekhorst, V.T.; Odenthal, J.; Bianchi, R.; van Helvert, S.; Ikenberg, K.; Ilina, O.; Stoma, S.; Xandry, J.; Jiang, L.Y.; et al. Hypoxia Induces a HIF-1-Dependent Transition from Collective-to-Amoeboid Dissemination in Epithelial Cancer Cells. Curr. Biol. 2017, 27, 392–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebestyen, E.; Zawisza, M.; Eyras, E. Detection of recurrent alternative splicing switches in tumor samples reveals novel signatures of cancer. Nucleic Acids Res. 2015, 43, 1345–1356. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Hoadley, K.A.; Parker, J.S.; Perou, C.M. Identification of mRNA isoform switching in breast cancer. BMC Genomics 2016, 17, 181. [Google Scholar] [CrossRef] [PubMed]
- Staddon, J.M.; Smales, C.; Schulze, C.; Esch, F.S.; Rubin, L.L. P120, a p120-related protein (p100), and the cadherin/catenin complex. J. Cell Biol. 1995, 130, 369–381. [Google Scholar] [CrossRef]
- Reynolds, A.B.; Daniel, J.; McCrea, P.D.; Wheelock, M.J.; Wu, J.; Zhang, Z. Identification of a new catenin: The tyrosine kinase substrate p120cas associates with E-cadherin complexes. Mol. Cell Biol. 1994, 14, 8333–8342. [Google Scholar] [CrossRef]
- Thoreson, M.A.; Anastasiadis, P.Z.; Daniel, J.M.; Ireton, R.C.; Wheelock, M.J.; Johnson, K.R.; Hummingbird, D.K.; Reynolds, A.B. Selective uncoupling of p120(ctn) from E-cadherin disrupts strong adhesion. J. Cell Biol. 2000, 148, 189–202. [Google Scholar] [CrossRef]
- Harris, T.J.; Tepass, U. Adherens junctions: From molecules to morphogenesis. Nat. Rev. Mol. Cell Biol. 2010, 11, 502–514. [Google Scholar] [CrossRef] [PubMed]
- Roczniak-Ferguson, A.; Reynolds, A.B. Regulation of p120-catenin nucleocytoplasmic shuttling activity. J. Cell Sci. 2003, 116, 4201–4212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, Y.Y.; Reynolds, A.B. Identification of murine p120 isoforms and heterogeneous expression of p120cas isoforms in human tumor cell lines. Cancer Res. 1996, 56, 2633–2640. [Google Scholar]
- Paredes, J.; Correia, A.L.; Ribeiro, A.S.; Schmitt, F. Expression of p120-catenin isoforms correlates with genomic and transcriptional phenotype of breast cancer cell lines. Cell Oncol. 2007, 29, 467–476. [Google Scholar] [PubMed]
- Miao, Y.; Liu, N.; Zhang, Y.; Liu, Y.; Yu, J.H.; Dai, S.D.; Xu, H.T.; Wang, E.H. P120ctn isoform 1 expression significantly correlates with abnormal expression of E-cadherin and poor survival of lung cancer patients. Med. Oncol. 2010, 27, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, H.-T.; Dai, S.-D.; Wei, Q.; Yuan, X.-M.; Wang, E.-H. Reduction of p120(ctn) isoforms 1 and 3 is significantly associated with metastatic progression of human lung cancer. Apmis 2007, 115, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Cichon, M.A.; Nelson, C.M.; Radisky, D.C. Regulation of epithelial-mesenchymal transition in breast cancer cells by cell contact and adhesion. Cancer Inform. 2015, 14, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.Y.; Park, J.I.; Cho, K.; Gu, D.; Ji, H.; Artandi, S.E.; McCrea, P.D. Shared molecular mechanisms regulate multiple catenin proteins: Canonical Wnt signals and components modulate p120-catenin isoform-1 and additional p120 subfamily members. J. Cell Sci. 2010, 123, 4351–4365. [Google Scholar] [CrossRef] [PubMed]
- van Hengel, J.; Vanhoenacker, P.; Staes, K.; van Roy, F. Nuclear localization of the p120(ctn) Armadillo-like catenin is counteracted by a nuclear export signal and by E-cadherin expression. Proc. Natl. Acad. Sci. USA 1999, 96, 7980–7985. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Kojima, S.; Borisy, G.G.; Green, K.J. P120 catenin associates with kinesin and facilitates the transport of cadherin-catenin complexes to intercellular junctions. J. Cell Biol. 2003, 163, 547–557. [Google Scholar] [CrossRef] [PubMed]
- van de Ven, R.A.; de Groot, J.S.; Park, D.; van Domselaar, R.; de Jong, D.; Szuhai, K.; van der Wall, E.; Rueda, O.M.; Ali, H.R.; Caldas, C.; et al. P120-catenin prevents multinucleation through control of MKLP1-dependent RhoA activity during cytokinesis. Nat. Commun. 2016, 7, 13874. [Google Scholar] [CrossRef] [PubMed]
- Winklbauer, R. Cell adhesion strength from cortical tension -an integration of concepts. J. Cell Sci. 2015, 128, 3687–3693. [Google Scholar] [CrossRef] [PubMed]
- Kourtidis, A.; Ngok, S.P.; Pulimeno, P.; Feathers, R.W.; Carpio, L.R.; Baker, T.R.; Carr, J.M.; Yan, I.K.; Borges, S.; Perez, E.A.; et al. Distinct E-cadherin-based complexes regulate cell behaviour through miRNA processing or Src and p120 catenin activity. Nat. Cell Biol. 2015, 17, 1145–1157. [Google Scholar] [CrossRef] [Green Version]
- Short, S.P.; Kondo, J.; Smalley-Freed, W.G.; Takeda, H.; Dohn, M.R.; Powell, A.E.; Carnahan, R.H.; Washington, M.K.; Tripathi, M.; Payne, D.M.; et al. P120-Catenin is an obligate haploinsufficient tumor suppressor in intestinal neoplasia. J. Clin. Investig. 2017. [Google Scholar] [CrossRef]
- Reichert, M.; Bakir, B.; Moreira, L.; Pitarresi, J.R.; Feldmann, K.; Simon, L.; Suzuki, K.; Maddipati, R.; Rhim, A.D.; Schlitter, A.M.; et al. Regulation of Epithelial Plasticity Determines Metastatic Organotropism in Pancreatic Cancer. Dev. Cell 2018, 45, 696–711. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venhuizen, J.-H.; Span, P.N.; van den Dries, K.; Sommer, S.; Friedl, P.; Zegers, M.M. P120 Catenin Isoforms Differentially Associate with Breast Cancer Invasion and Metastasis. Cancers 2019, 11, 1459. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101459
Venhuizen J-H, Span PN, van den Dries K, Sommer S, Friedl P, Zegers MM. P120 Catenin Isoforms Differentially Associate with Breast Cancer Invasion and Metastasis. Cancers. 2019; 11(10):1459. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101459
Chicago/Turabian StyleVenhuizen, Jan-Hendrik, Paul N. Span, Koen van den Dries, Sebastian Sommer, Peter Friedl, and Mirjam M. Zegers. 2019. "P120 Catenin Isoforms Differentially Associate with Breast Cancer Invasion and Metastasis" Cancers 11, no. 10: 1459. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101459