Table Olives More than a Fermented Food
by
 ,
,
Giorgia Perpetuini
, 
Roberta Prete
, 
Natalia Garcia-Gonzalez
,
Mohammad Khairul Alam
and
Aldo Corsetti
* Faculty of BioScience and Technology for Food, Agriculture and Environment, University of Teramo, 641000 Teramo, Italy
*
Author to whom correspondence should be addressed.
Foods 2020, 9(2), 178; https://0-doi-org.brum.beds.ac.uk/10.3390/foods9020178
Submission received: 16 December 2019
/
Revised: 5 February 2020
/
Accepted: 7 February 2020
/
Published: 12 February 2020
(This article belongs to the Special Issue Research on Characterization and Processing of Table Olives)
Abstract
:Table olives are one of the oldest vegetable fermented foods in the Mediterranean area. Beside their economic impact, fermented table olives represent also an important healthy food in the Mediterranean diet, because of their high content of bioactive and health-promoting compounds. However, olive fermentation is still craft-based following traditional processes, which can lead to a not fully predictable final product with the risk of spontaneous alterations. Nowadays, food industries have to face consumer demands for safe and healthy products. This review offers an overview about the main technologies used for olive fermentation and the role of lactic acid bacteria and yeasts characterizing this niche during the fermentation. Particular attention is offered to the selection and use of microorganisms as starter cultures to fasten and improve the safety of table olives. The development and implementation of multifunctional starter cultures in order to obtain heath-oriented table olives is also discussed.
1. Introduction
Table olives are defined as “the sound fruit of varieties of the cultivated olive trees (Olea europaea L.) that are chosen for their production of olive whose volume, shape, flash-to-stone ratio, fine flesh, taste, firmness, and ease of detachment from the stone make them particularly suitable for processing; treated to remove their bitterness and preserved by natural fermentation; or by heat treatment, with or without the addition of preservatives; packed with or without covering liquid” [1]. Table olives are considered one of the oldest fermented vegetables in the Mediterranean basin and are an important element for the economy of several countries. Their production exceeded 2.9 million tons in the 2017/2018 season and the main producers are Spain, Egypt, Turkey, Algeria, Italy, Greece, and Portugal [2]. However, their production is increasing also in other countries, such as South America, Australia, and the Middle East [2]. Moreover, in 2010 they have been added in the Healthy Eating Pyramid of the Mediterranean diet (https://dietamediterranea.com/), because of their high content of bioactive compounds, dietary fibers, fatty acids, and antioxidants [3].
The olive fruit is a drupe which cannot be consumed directly from the tree because of the presence of a bitter compound called oleuropein. The bitterness can be removed by alkaline treatment, or by brining/salting, fermentation, and acidification [4]. According to the International Olive Oil Council (IOOC) [1], the main goals of olive processing are to improve their sensory characteristics and to ensure safety of consumption. The “trade standard applying to table olives” [1] describes the type of preparation of table olives; however, some traditional processes are still applied, such as the Castelvetrano system. This method is diffused in Sicily and mainly is based on the exploitation of the Nocellara del Belice variety. Only olives of more than 19 mm in diameter are used, which are placed in vessels and treated with a 1.8%–2.5% NaOH solution for one hour. After that, 5–8 kg of salt are added, and the olives are maintained in this brine for 10–15 days. A mild washing step is performed to avoid the total elimination of lye [5].
The main trade preparations are reported in Table 1.
2. Table Olives Associated Microbiota
Olive fermentation is a complex process involving a wide array of microorganisms and mainly lactic acid bacteria (LAB) (e.g. Lactobacillus plantarum and Lactobacillus pentosus) and yeasts (Saccharomyces cerevisiae, Wickerhamomyces anomalus, Candida boidinii, etc.) [6]. Their enzymatic activities shape the characteristics of the final products, e.g., flavor, texture, and safety [6]. Moreover, strains isolated from table olives show specific probiotic traits and are able to adhere to the fruit’s epidermis, which could thus be ingested by consumers, turning olives into a carrier for these beneficial microbes [7].
The role of LAB during olive fermentation has been investigated in detail [8,9,10,11,12,13,14,15]. The majority of studies indicated that L. plantarum and L. pentosus are the main LAB isolated from table olives [10,13,15,16]. They are facultative heterofermentative; therefore, they can produce different end products, such as lactic acid, acetic acid, and carbon dioxide or only produce lactic acid depending on the environmental conditions [13]. Hurtado et al. [13] highlighted that L. plantarum produced a higher amount of acetic acid during olive fermentation than L. pentosus, suggesting the lower ability of the latter species to preserve a homofermentative metabolism under stress conditions. The main species are reported in Figure 1. LAB are the main bacteria responsible of olive debittering thanks to their enzymatic reservoir (β-glucosidase and esterase). L. pentosus is characterized by a strong β-glucosidase activity [11]. This enzyme catalyzes oleuropein degradation and the release of glucose and aglycone. This last compound is converted to non-bitter compounds, such as elenolic acid and hydroxytyrosol, by an esterase [17]. They also play a key role in the decrease of pH and provide microbiological stability to the final product as well as an extended shelf life. The production of lactic acid induces an acidification of brine that prevent the growth of spoilage microorganisms and pathogens [17,18].
Yeasts can play a double role during olive fermentation; in fact, they are associated with the production of volatile compounds (e.g., alcohols, ethyl acetate, and acetaldehyde) and metabolites that improve the taste and aroma and the preservation characteristics of this fermented food. Moreover, they can enhance LAB growth by the release of nutritive compounds, either synthesizing vitamins, amino acids, and purins, or by metabolizing complex carbon sources [19,20,21]. Finally, they show esterase and lipase activities. The first one improves the olive taste since it is involved in the production of esters from free fatty acids, while the second one changes the free fatty acids composition of olives improving the characteristics of the final product [22]. On the other hand, yeasts may cause gas-pocket formation and softening of the olive tissue, or even package bulging, clouding of the brines, and production of off flavors and odors [20].
Microbiological studies revealed that W. anomalus, S. cerevisiae, Pichia kluyveri, and Pichia membranifaciens are the yeast mainly present in olive brine [6,20,23,24]. S. cerevisiae and several species of the Pichia genus showed antioxidant activity which protects fruits from oxidation and peroxide formation [21]. Hernandez et al. [21,25] underlined the relevance of W. anomalus during olive fermentation. In fact, it presents β-glucosidase activity, as well as produces anti-oxidant compounds and killer toxins against human pathogens and spoilage microorganisms.
Moreover, D. hansenii, P. membranifaciens, and W. anomalus showed strain-specific killer activity against spoilage yeasts [20,23,25,26].
A recent study started to study the biogeography of the microbial communities associated with Spanish-style green olive fermentations [27]. The authors studied the microbial biodiversity of 30 ten-ton fermenters of three different fermentations yards (patios) during the fermentation process. Some species were constant, representing the core microbiota of this area. L. pentosus, Pediococcus parvulus, Lactobacillus collinoides/paracollinoides, Lactobacillus coryniformis, L. plantarum, Pichia manshurica, and Candida thaimueangensis were found in every patio. In particular, cosmopolitan strains belonged to the following species: L. pentosus, P. parvulus, L. collinoides/paracollinoides, and P. manshurica.
3. Microbial Spoilage of Table Olives
Olive fermentation is still craft-based; therefore, it is not fully predictable, and some alterations can occur. During the first phase of Spanish fermentation, the Gram-negative bacteria prevail. This phase lasts until LAB grow up inducing a decrease in pH. If this reduction is not too fast, “gas pockets”, resulting in the softening and breakage of the cuticle, can appear [28]. A high pH can also favor the development of Clostridium spp., which could induce a putrid or butyric fermentation, which cause the appearance of off-flavors and off-odors [28].
The softening of olive drupe is another alteration due to the development of pectinolytic yeasts (e.g., P. manshurica, Pichia kudriavzevii,, Saccharomyces oleaginosus, etc.), molds (Aspergillus niger, Fusarium spp., and Penicillium spp.) and some bacteria (Bacillus spp., Aerobacter spp., etc.) [29]. These microorganisms release degrading enzymes, which act on pectic substances and cellulose, hemicellulose, and polysaccharides, causing the loss of the structural integrity of the olive drupe [28,29].
Seville-style table olives can undergo a defect called “white spot”. These spots develop between the skin and the flesh and are associated to the development of some L. plantarum strains [30].
Finally, when the final product is not pasteurized Propionibacterium can develop, producing acetic and propionic acids. This alteration is called “zapateria” and cause an increase in volatile acidity and the formation of cyclohexanecarboxylic acid [31] and the production of biogenic amines, such as cadaverine and tyramine [32].
4. Table Olives’ Starter Cultures
The use of starter cultures for table olives fermentation is highly recommended [17]. An appropriate inoculum reduces the effects of spoilage microorganisms, inhibits the growth of pathogenic microorganisms, and helps to achieve a controlled process, reducing debittering time and improving the sensorial and hygienic quality of the final product [17,33,34,35]. Two different types of starter cultures can be applied. Natural starter cultures are made up of microorganisms that spontaneously colonize the raw materials [3]. Their composition is often not reproducible; however, they guarantee a high biodiversity, which contributes to enrich the final product with particular sensory characteristics mostly linked to the region of origin of the raw material itself [36]. On the other hand, selected starter cultures provide numerous advantages (Table 2). They are usually represented by a single strain or by a mixture of strains previously selected on the basis of specific features: A high survival capacity in the fermentation environment (low pH, high concentrations of salts, and low fermentation substrates); high acidifying activity (through organic acid production); the ability to hydrolyze phenolic compounds (such as oleuropein); as well as the possibility of producing volatile molecules and/or specific enzymatic activities that contribute positively to the development of the sensory profile of the final product [17]. Another important characteristic of a starter culture is its ability to dominate the indigenous microbiota [17]. Dominance of the starter culture would be exerted by its fast and predominant growth under fermentation conditions and/or its ability to produce antagonistic substances [37]. In addition, for commercial purpose, it is necessary that starter cultures resist the freezing or freeze-drying process [17].
Despite these advantages, the application of starter cultures for olive fermentation is still limited [6]. Some of the most important olive varieties are still processed without their addition [3].
Among LAB species, the most often proposed as starter cultures are L. plantarum and L. pentosus [15,17,38], used alone or in combination with other bacterial or yeast species (Table 3 and Table 4).
Several studies were conducted to drive the fermentation processes and to improve the quality and sensory profiles of different table olive cultivars using both autochthonous and commercial oleuropeinolytic strains belonging to the L. plantarum group [33,34,39,40,41,42,43,44].
Different L. pentosus and L. plantarum starter cultures have been found to dominate and improve the fermentation process of green table olives in terms of processing time, microbiological quality, color stability, and aroma profile [39,40,41].
A strain of Lb. pentosus (1MO) was used as a starter to shorten the debittering process of different cultivars (cv. Itrana and Leccino) at the pilot and industrial scale [45]. The use of the selected strain L. pentosus (1MO) significantly improved the quality and safety aspects of the fermented table olives, allowing to successfully end the fermentation process within eight days, while more than one week or even months are usually required for biological spontaneous fermentation [46,47,48].
Recently, a starter culture made up of two L. pentosus strains was successfully used to debitter green table olives (cv. Itrana) [35] and was patented (Patent N0. 0001428559).
Interestingly, the use of L. plantarum strains as starter strains has been investigated also for the ability to positively affect the fermentation process in term of quality preservation and stability during storage. Sherhai et al. [42] found a protective effect of L. plantarum on fatty acid oxidation and peroxidation processes, as well as a strong antioxidant activity during the Spanish-style fermentation process. In line with that, a recent study on inoculated Nocellara Etnea table olives with six different starter cultures made up of L. plantarum, L. pentosus, and L. paracasei confirmed the dominance of L. plantarum during fermentation and its positive impact on table olives [34].
Furthermore, a sequential inoculation strategy has been proposed as a promising biotechnological tool to produce low salt Nocellara Etnea table olives. The authors reported on the use of a β-glucosidase-positive strain, L. plantarum strain, followed after 60 days by the inoculum of a L. paracasei probiotic strain. This strategy reduced the processing time, and positively affected the polyphenol content and sensory profile of the final product, which was characterized by a low salt concentration (5%) [43].
In recent years, several studies focused on the development of yeast starter cultures, both alone and in combination with LAB [20,23,26,49,50,51]. L. plantarum and L. pentosus strains have been used with excellent results in combination with an autochthonous Wickerhamomyces anomalus strain to accelerate the fermentation of Bella di Cerignola table olives [33]. A functional starter strain of L. pentosus, with and without P. membranifaciens, was successfully used to drive fermentations of Conservolea black olives, which allow producing a functional product with an improved sensory profile [52].
A sequential inoculation strategy (firstly yeasts, then bacteria) was developed by Tufariello et al. [53]. In particular, the authors tested different yeast species (S. cerevisiae, D. hansenii, and W. anomalus) in combination with L. plantarum and Leuconostoc mesenteroides in order to improve the sensory and organoleptic properties of table olives. Pilot-scale fermentations with the sequential inoculation of LAB and yeast strains reduced the fermentation time (from 180 to 90 days), as well as improved the organoleptic characteristics of the final product [53].
Other yeasts species, such as Debaryomyces spp., Pichia spp., and Rhodotorula spp., were recently investigated in order to select the appropriate strains to use in combination with LAB [3,54] (Table 2).
Bonatsou et al. [54] selected P. guilliermondii and W. anomalus among several yeast strains, isolated from black table olives, and screened for their technological and probiotic properties as promising multifunctional starters to use in real olive fermentations. The use of yeasts is also linked to their ability to favor the formation of multispecies biofilms on biotic (drupes) and abiotic (fermenter vats) surfaces [3]. Several studies showed the ability of some yeast species, such as D. hansenii, Geotrichum candidum, P. guilliermondii, and W. anomalus, to form biofilm and create a positive environment for L. pentosus growth [7,23,55,56,57,58,59].
Recently, the application of autochthonous strains has arisen to face consumers’ demand for more traditional products with a unique sensory profile and peculiar organoleptic properties [60]. Autochthonous strains, being well adapted to the raw material conditions, can easily lead the fermentation process by dominating the table olives microenvironment [3,51]. However, only few studies report the application of autochthonous starter cultures [36,53,61,62]. Martorana et al. [36] used autochthonous starter cultures as a “Pied de cuve” to ferment Nocellara del Belice olives [36]. The application of autochthonous starter cultures could be useful for achieving IGP and PDO (Protected Designation of Origin) product specifications, linking the fermented final product to the region where it comes from [3].
5. New Trend in Olive Production: Probiotic Table Olives
The concept of functional food was born in Japan around the 1980s; in 1991, the acronym FOSHU (Foods for Specified Health Use) was coined. Nowadays, the accepted definition is the one recognized by the European Union Food Information Council (EUFIC), based on which functional foods are defined as “foods similar in appearance to conventional foods that are consumed as part of a normal diet, and have demonstrated physiological benefits and/or the capacity to reduce the risk of chronic disease beyond their basic nutritional functions” [88]. Probiotics and prebiotics represent the most-used strategies for the production of functional foods [89,90,91,92,93,94]. Probiotics are defined as “live microorganisms which, when administered in adequate amounts, as part of a food or a supplement, confer a health benefit on the host” [95]. Generally, probiotics are bacteria isolated from human sources, mostly from the gastrointestinal tract [96], and mainly belong to Bifidobacterium and Lactobacillus genera [88]. Indeed, it has been recently showed that also naturally occurring food-associated microbes can reach the gut as viable cells, interact with the human host, and potentially provide benefits to gut health [97]. In this context, a diet may represent not only a source of nutrients to the body, but can be also a vehicle of exogenous microorganisms with positive effects on human health [98,99].
Table olives represent a wide reservoir of putative beneficial microbes. Thus, several studies have been conducted to assess the probiotic effects of strains isolated from different fermented olives cultivars and/or already used as starter cultures, belonging to the most widely spread species L. plantarum and L. pentosus, as well as to species less frequently used, such as L. paracasei, L. casei, and L. paraplantarum [43,85,100,101,102,103,104,105,106]. Some studies revealed that some LAB strains isolated from table olives were able to adhere to porcine jejune epithelial cells IPEC-J2 and produced antimicrobial compounds able to inhibit Helicobacter pylori, Propionibacterium spp., and Clostridium perfringens [10,70,107,108,109]. Probiotic potential, based on the ability to outcompete foodborne pathogens for cell adhesion, was also characterized in several L. pentosus isolated from different table olive cultivars (i.e., Nocellara del Belice and Aloreña green table olives) [103,110]. Strains isolated from both cultivars showed the ability to adhere to human intestinal epithelial Caco-2 [110] and vaginal cells [103], as well as the ability to auto-aggregate and co-aggregate with pathogenic bacteria, to ferment some prebiotics, and to in vivo exert protective effects in Caenorhabditis elegans [103,110]. Beside antimicrobial activity, different strains of L. pentosus and L. plantarum isolated from table olives stimulated the release of pro-inflammatory (IL-6) and anti-inflammatory (IL-10) interleukins on macrophages, suppressed the secretion of IL-8, and showed anti-proliferative activity on the HT-29 cell line [111].
Table olives of different cultivars have already been validated as a promising carrier for delivering different probiotics strains into the human GI tract [112] (patent application EP2005/0104138 [9,113]. Table olives can be considered an ideal matrix for the survival of probiotics due to the nutrients released by the fruits and the fact that drupes are coated with a hydrophobic epicuticular wax that promote microbial adhesion [6,7,52,56,112,113,114,115,116].
The probiotic L. paracasei strain LMGP22043 was able to colonize the human gut, positively influencing fecal bacteria and biochemical parameters [113]. Lavermicocca et al. [112] used table olives as carrier for the probiotic L. paracasei strain IMPC2.1. The strain was recovered in human feces after fermented olive intake, confirming the possibility to use table olives as carrier of probiotics into the human gastrointestinal tract [112]. An autochthonous potential probiotic L. pentosus strain [23,75,103] showed to be able to survive for 200 days in packed olives, confirming the possibility to incorporate probiotic strains and thus produce functional table olives [76].
The genetic basis of LAB strains adhesion on olive surfaces is still in its infancy. Perpetuini et al. [115] revealed that the sessile state represented the prevailing L. pentosus life-style during table olive fermentation and that the three genes enoA1, gpi and obaC were necessary in L. pentosus to form an organized biofilm on the olive skin. The first two genes encoded for cytosolic enzymes involved in the glycolysis pathway and in the adhesion to some specific components of olive skin, while obaC for a putative fatty acid binding protein of the DegV family, which could bind some lipids of the epicuticular wax. More recently, Pérez Montoro et al. [116] analyzed the adhesion to mucin of L. pentosus strains isolated from Aloreña green table olives. They revealed the presence of four moonlighting proteins over-produced in adhesive strains, which were not produced in non-adhesive strains. These proteins were involved in the glycolytic pathway (phosphoglycerate mutase and glucosamine-6-phosphate deaminase), stress response (small heat shock protein), and transcription (transcription elongation factor GreA). A new in silico approach confirmed that moonlighting proteins are involved in the adhesion to both the extracellular matrix (i.e., olive surface) and host cells, as well as in host immunomodulation [117]. Due to the importance of the genetic background on health-promoting traits, Calero-Delgado et al. [118] recently published the draft genome sequences of five L. pentosus strains isolated from biofilms on the skin of green table olives. In particular, most of the strains evaluated harbored two copies of the luxS gene, involved in the production of the universal bacterial communicator autoinductor-2. Genes encoding for bacteriocin, exopolysaccharide, and MucBP proteins, which could play an important role in microbe-eukaryote cell adhesion, were also found [118]. The main feature of these studied strains was their ability to adhere to the surface of olives during fermentation, forming biofilms, and turning table olives into carriers of beneficial microorganisms to consumers [114,115,119].
Recently, different studies have been focusing on the yeast microbiota associated with table olives fermentations in order to find potential probiotic candidates to be used as starter cultures [23,26,54,100,101,102,103,104,105,106]. Saccharomyces boulardii represent the only yeast with claimed probiotic effects [120]. Evidences of other yeast species showing probiotic features, mainly associated with table olive microbiota, such as D. hansenii, T. delbrueckii, K. lactis, and S. cerevisiae, are emerging [121,122,123,124].
Different Torulaspora delbrueckii and Debaryomyces hansenii strains have been found to survive in the presence of high bile salt concentrations and low pH values, as well as to have antimicrobial activity against foodborne pathogens [26]. Furthermore, Silva et al. [125] found some P. membranifaciens and Candida oleophila strains within a native yeast population of Portuguese olives to be promising candidates as multifunctional starter cultures, by having both technological (oleuropeinolytic activity) and beneficial potential (vitamins production, mycogenic, and antimicrobial activities).
In this context two important issues to be considered are the assessment of technological factors influencing the survival of probiotic starter cultures and the starter effect on olives’ sensory profile. Rodríguez-Gómez et al. [77] evaluated the effects of inoculation strategies on the survival of L. pentosus TOMC-LAB2—a potential probiotic strain when used as a starter culture in large-scale fermentations of green Spanish-style olives. They proposed an inoculation immediately after brining to reduce the presence of initial natural microbiota, the re-inoculation to replace the possible initial died starter and an early processing in the season when starter survival is higher. Concerning the second aspect, a recent study analyzed the organoleptic characteristics of traditional, spontaneously fermented green table olives and green table olives inoculated with L. pentosus TOMC-LAB2. Consumers perceived them similarly, only saltiness had a marked adverse effect [78].
Probiotics are generally carried through dairy products. However, the increased incidence of lactose intolerance, concerns over cholesterol, and the wide spread of new lifestyles (vegans and vegetarians) drove new researches toward non-dairy probiotic foods, such as fruits and vegetables, which are rich in vitamins, minerals, carbohydrates, fibers, and antioxidant compounds [126,127]. Recently, it has been shown that vegetable-derived products (i.e., fruits, fruits juices, cereals, and legumes) can act as carriers for positive microbes because of their intrinsic structure; thus, microorganisms can colonize pores, lesions, lenticels, and irregularities present on the surface [119]. Moreover, vegetables are also rich in prebiotic compounds, which protect probiotic microorganisms from the harsh GI tract conditions and are a source of nutrients that positively influences bacterial survival [128,129]. Actually, vegetable-based probiotic foods are available on the market. However, further studies are necessary to better understand the viability of selected strains in the human GI tract and their interactions with human microbiota. In vivo studies are required to assess if carried bacteria and the food matrix have a positive impact on human health. In this case, health claims could be proposed.
6. Conclusions
Table olives have a great impact on the economy of several countries. According to Bonatsou et al. [6], olives are considered in the food industry as the “food of the future”. Despite the many advances made, table olives are still produced according to ancient and local recipes, refusing the addition of starter cultures. Olive industries will face several challenges in the next future, including crop management, olive quality, production methods, and health issues. The application of starter cultures represents the main biotechnological challenge/innovation in this field. In this review the main criteria used for starter cultures selection are reported. LAB and yeasts are the main microbial groups studied and several strains have been characterized in order to develop new starter cultures. The use of autochthonous starter cultures is gaining attention since they offer several advantages in terms of adaptability to stressful niches and characterization of the final product, offering a link with the product origin. Another interesting aspect is the characterization of probiotic strains. This issue is the main research trend in this field since it responds to consumer demand for health-oriented products. The potential addition of probiotics in table olive fermentation on one hand give rise to new questions to be solved in terms of cost-effectiveness and acceptance by consumers, but on the other hand can improve the entire production process by positively affecting the aroma and sensory profile, product shelf-life, and by providing additional health-promoting properties to the consumers. Moreover, the development of probiotic table olives could have a positive economic impact, since this product is produced also in less developed countries.
In our opinion, further studies are necessary to isolate and characterize more strains from different table olive cultivars in order to prepare autochthonous starter culture collections and produce healthy products with enhanced sensory characteristics. Additional researches are also needed to implement fermentation strategies to favor the survival and dominance of starter strains and develop new starters by combining LAB and yeasts, to mimic the natural microbiota of olives. Moreover, concerning probiotic strains, further validation in in vivo trials with more complex animal or human systems should be performed to gain a deeper understanding of their potential health-promoting features for humans. Finally, further studies should develop new approaches for the treatment of wastewater produced by table olive industries in order to have healthy eco-friendly products.
Author Contributions
A.C. conceived the idea; G.P., R.P. and A.C. drafted the paper; N.G.-G. and M.K.A. prepared the tables and figure; All authors have read and agreed to the published version of the manuscript.
Funding
This work is part of the project “Tracciabilità, certificazione e tutela della qualità dell’olio di oliva e delle olive da tavola Azione 4d” supported by a grant from UNAPROL (Reg. CE No. 867/2008 Misura 4). It has received financial support from the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska-Curie grant agreement 713714 ESR 07 to Natalia Garcia-Gonzalez and from “PON Ricerca e Innovazione 2014-20,” azione I.1 “Dottorati innovativi con caratterizzazione industriale,” A.Y. 2018-19, XXXIV Cycle, for the PhD project grant of Mohammad Khairul Alam.
Conflicts of Interest
The authors declare no conflict of interest.
References
- IOOC, International Olive Oil Council. Trade Standard Applying to Table Olives; International Olive Oil Council: Madrid, Spain, 2004. [Google Scholar]
- IOOC, International Olive Oil Council. World Table Olive Figures. 2019. Available online: http://www.internationaloliveoil.org/estaticos/view/132-world-table-olive-figures (accessed on 20 January 2019).
- Campus, M.; Degirmencioglu, N.; Comunian, R. Technologies and Trends to Improve Table Olive Quality and Safety. Front. Microbiol. 2018, 9, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido-Fernández, A.; Fernández-Díez, M.J.; Adams, R.M. Table Olives: Production and Processing, 1st ed.; Chapman and Hall: London, UK, 1997. [Google Scholar]
- Rejano, L.; Montaño, A.; Casado, F.J.; Sánchez, A.H.; de Castro, A. Table olives: Varieties and variations. In Olives and Olive Oil in Health and Disease Prevention; Preedy, V.R., Watson, R.R., Eds.; Oxford Academic Press: Oxford, UK, 2010; pp. 5–15. [Google Scholar]
- Bonatsou, S.; Tassou, C.C.; Panagou, E.Z.; Nychas, G.J.E. Table olive fermentation using starter cultures with multifunctional potential. Microorganisms 2017, 5, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benítez-Cabello, A.; Romero-Gil, V.; Rodríguez-Gómez, F.; Garrido-Fernández, A.; Jiménez-Díaz, R.; Arroyo-López, F.N. Evaluation and identification of poly-microbial biofilms on natural green Gordal table olives. Antonie Van Leeuwenhoek 2015, 108, 597–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randazzo, C.L.; Restuccia, C.; Romano, A.D.; Caggia, C. Lactobacillus casei, dominant species in naturally fermented Sicilian green olives. Int. J. Microbiol. 2004, 90, 9–14. [Google Scholar] [CrossRef]
- De Bellis, P.; Valerio, F.; Sisto, A.; Lonigro, S.L.; Lavermicocca, P. Probiotic table olives: Microbial populations adhering on olive surface in fermentation sets inoculated with the probiotic strain Lactobacillus paracasei IMPC2.1 in an industrial plant. Int. J. Food Microbiol. 2010, 140, 6–13. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Altieri, C.; Corbo, M.R.; Sinigaglia, M.; Ouoba, L.I.I. Characterization of lactic acid bacteria isolated from Italian Bella di Cerignola table olives: Selection of potential multifunctional starter cultures. J. Food Sci. 2010, 75, 536–544. [Google Scholar] [CrossRef]
- Franzetti, A.; Gandolfi, I.; Gaspari, E.; Ambrosini, R.; Bestetti, G. Seasonal variability of bacteria in fine and coarse urban air particulate matter. Appl. Microbiol. Biotechnol. 2011, 90, 745–753. [Google Scholar] [CrossRef]
- Aponte, M.; Blaiotta, G.; La Croce, F.; Mazzaglia, A.; Farina, V.; Settanni, L. Use of selected autochthonous lactic acid bacteria for Spanish style table olive fermentation. Food Microbiol. 2012, 30, 8–16. [Google Scholar] [CrossRef]
- Hurtado, A.; Reguant, C.; Esteve-Zarzoso, B.; Bordons, A.; Rozès, N. Microbial population dynamics during the processing of Aberquina table olives. Food Res. Int. 2008, 41, 738–744. [Google Scholar] [CrossRef]
- Doulgeraki, A.I.; Hondrodimou, O.; Iliopoulos, V.; Panagou, E.Z. Lactic acid bacteria and yeast heterogeneity during aerobic and modified atmosphere packaging storage of natural black Conservolea olives in polyethylene pouches. Food Control 2012, 26, 49–57. [Google Scholar] [CrossRef]
- Tofalo, R.; Perpetuini, G.; Schirone, M.; Ciarrocchi, A.; Fasoli, G.; Suzzi, G.; Corsetti, A. Lactobacillus pentosus dominates spontaneous fermentation of Italian table olives. LWT Food Sci. Technol. 2014, 57, 710–717. [Google Scholar] [CrossRef]
- Campaniello, D.; Bevilacqua, A.; D’Amato, D.; Corbo, M.R.; Altieri, C.; Sinigaglia, M. Microbial characterization of table olives processed according to Spanish and natural styles. Food Technol. Biotechnol. 2005, 43, 289–294. [Google Scholar]
- Corsetti, A.; Perpetuini, G.; Schirone, M.; Tofalo, R.; Suzzi, G. Application of starter cultures to table olive fermentation: An overview on the experimental studies. Front. Microbiol. 2012, 3, 248. [Google Scholar] [CrossRef] [Green Version]
- Caggia, C.; Randazzo, C.L.; Di Salvo, M.; Romeo, F.V.; Giudici, P. Occurrence of Listeria monocytogenes in green table olives. J. Food Prot. 2004, 67, 2189–2194. [Google Scholar] [CrossRef]
- Alves, M.; Gonçalves, T.; Quintas, C. Microbial quality and yeast population dynamics in cracked green table olives’ fermentations. Food Control 2012, 23, 363–368. [Google Scholar] [CrossRef]
- Arroyo-López, F.N.; Querol, A.; Bautista-Gallego, J.; Garrido-Fernández, A. Role of yeasts in table olive production. Int. J. Food Microbiol. 2008, 128, 189–196. [Google Scholar] [CrossRef]
- Hernández, A.; Martín, A.; Aranda, E.; Pérez-Nevado, F.; Córdoba, M.G. Identification and characterization of yeast isolated from the elaboration of seasoned green table olives. Food Microbiol. 2007, 24, 346–351. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Rodriguez-Gomez, F.; Barrio, E.; Querol, A.; Garrido- Fernandez, A.; Arroyo-López, F.N. Exploring the yeast biodiversity of green table olive industrial fermentations for technological applications. Int. J. Food Microbiol. 2011, 147, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-López, F.N.; Romero-Gil, V.; Bautista-Gallego, J.; Rodríguez-Gómez, F.; Jiménez-Díaz, R.; García-García, P.; Querol, A.; Garrido-Fernández, A. Potential benefits of the application of yeast starters in table olive processing. Front. Microbiol. 2012, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Tofalo, R.; Perpetuini, G.; Schirone, M.; Suzzi, G.; Corsetti, A. Yeast biota associated to naturally fermented table olives from different Italian cultivars. Int. J. Food Microbiol. 2013, 161, 203–208. [Google Scholar] [CrossRef]
- Hernández, A.; Martín, A.; Córdoba, M.G.; Benito, M.J.; Aranda, E.; Pérez-Nevado, F. Determination of killer activity in yeasts isolated from the elaboration of seasoned green table olives. Int. J. Food Microbiol. 2008, 121, 178–188. [Google Scholar] [CrossRef]
- Psani, M.; Kotzekidou, P. Technological characteristics of yeaststrains and their potential as starter adjuncts in Greek-style black olive fermentation. World J. Microbiol. Biotechnol. 2006, 22, 1329–1336. [Google Scholar] [CrossRef]
- Lucena-Padrós, H.; Ruiz-Barba, J.L. Microbial biogeography of Spanish-style green olive fermentations in the province of Seville, Spain. Food Microbiol. 2019, 82, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Lanza, B. Abnormal fermentations in table-olive processing: Microbial origin and sensory evaluation. Front. Microbiol. 2013, 4, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golomb, B.L.; Morales, V.; Jung, A.; Yau, B.; Boundy-Mills, K.L.; Marco, M.L. Effects of pectinolytic yeast on the microbial composition and spoilage of olive fermentations. Food Microbiol. 2013, 33, 97–106. [Google Scholar] [CrossRef]
- Kailis, S.; Harris, D. Producing Table Olives; Landlinks Press: Collingwood, VIC, Australia, 2007; p. 328. [Google Scholar]
- Montano, A.; de Castro, A.; Rejano, L.; Brenes, M. 4- hydroxycyclohexanecarboxylic acid as a substrate for cyclohexane carboxylic acid production during the “Zapatera” spoilage of Spanish-style green table olives. J. Food Prot. 1996, 59, 657–662. [Google Scholar] [CrossRef]
- Garcia, P.G.; Barranco, C.R.; Quintana, M.C.; Fernandez, A.G. Biogenic amine formation and “zapatera” spoilage of fermented green olives: Effect of storage temperature and debittering process. J. Food Prot. 2004, 67, 117–123. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Campanella, D.; Cosmai, L.; Summo, C.; Rizzello, C.G.; Caponio, F. Microbiota and metabolome of un-started and started Greek-type fermentation of Bella di Cerignola table olives. Food Microbiol. 2015, 52, 18–30. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Todaro, A.; Pino, A.; Corona, O.; Caggia, C. Microbiota and metabolome during controlled and spontaneous fermentation of Nocellara Etnea table olives. Food Microbiol. 2017, 65, 136–148. [Google Scholar] [CrossRef]
- Perpetuini, G.; Caruso, G.; Urbani, S.; Schirone, M.; Esposto, S.; Ciarrocchi, A.; Prete, R.; Garcia-Gonzalez, N.; Battistelli, N.; Gucci, R.; et al. Changes in polyphenolic concentrations of table olives (cv. Itrana) produced under different irrigation regimes during spontaneous or inoculated fermentation. Front. Microbiol. 2018, 9, 1287. [Google Scholar] [CrossRef]
- Martorana, A.; Alfonzo, A.; Settanni, L.; Corona, O.; La Croce, F.; Caruso, T.; Moschetti, G.; Francesca, N. An innovative method to produce green table olives based on “pied de cuve” technology. Food Microbiol. 2015, 50, 126–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marugg, J.D. Bacteriocins, their role in developing natural products. Food Biotechnol. 1991, 5, 305–312. [Google Scholar] [CrossRef]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozès, N. Lactic acid bacteria from fermented table olives. Food Microbiol. 2012, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Comunian, R.; Ferrocino, I.; Paba, A.; Daga, E.; Campus, M.; Di Salvo, R.; Cauli, E.; Piras, F.; Zurru, E.; Cocolin, L. Evolution of microbiota during spontaneous and inoculated Tonda di Cagliari table olives fermentation and impact on sensory characteristics. LWT Food Sci. Technol. 2017, 84, 64–72. [Google Scholar] [CrossRef]
- Ramírez, E.; Medina, E.; García, P.; Brenes, M.; Romero, C. Optimization of the natural debittering of table olives. LWT Food Sci. Technol. 2017, 77, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Chranioti, C.; Kotzekidou, P.; Gerasopoulos, D. Effect of starter cultures on fermentation of naturally and alkali-treated cv. Conservolea green olives. LWT Food Sci. Technol. 2018, 89, 403–408. [Google Scholar] [CrossRef]
- Sherahi, M.H.A.; Shahidi, F.; Yazdi, F.T.; Hashemi, S.M.B. Effect of Lactobacillus plantarum on olive and olive oil quality during fermentation process. LWT Food Sci. Technol. 2018, 89, 572–580. [Google Scholar] [CrossRef]
- Pino, A.; Vaccalluzzo, A.; Solieri, L.; Romeo, F.V.; Todaro, A.; Caggia, C.; Arroyo-López, F.N.; Bautista-Gallego, J.; Randazzo, C.L. Effect of Sequential Inoculum of Beta-Glucosidase Positive and Probiotic Strains on Brine Fermentation to Obtain Low Salt Sicilian Table Olives. Front. Microbiol. 2019, 10, 174. [Google Scholar] [CrossRef]
- Caponio, F.; Difonzo, G.; Calasso, M.; Cosmai, L.; De Angelis, M. Effects of olive leaf extract addition on fermentative and oxidative processes of table olives and their nutritional properties. Food Res. Int. 2019, 116, 1306–1317. [Google Scholar] [CrossRef]
- Servili, M.; Settanni, L.; Veneziani, G.; Esposto, S.; Massitti, O.; Taticchi, A.; Urbani, S.; Montedoro, G.F.; Corsetti, A. The use of Lactobacillus pentosus 1MO to shorten the debittering process time of black table olives (Cv. Itrana and Leccino): A pilot-scale application. J. Agric. Food Chem. 2006, 54, 3869–3875. [Google Scholar] [CrossRef]
- Tassou, C.C.; Panagou, E.Z.; Katsaboxakis, K.Z. Microbiological and physicochemical changes of naturally black olives fermented at different temperatures and NaCl levels in the brines. Food Microbiol. 2002, 19, 605–615. [Google Scholar] [CrossRef]
- Ciafardini, G.; Marsilio, A.; Lanza, B.; Pozzi, N. Hydrolysis of oleuropein by Lactobacillus plantarum strains associated with olive fermentation. Appl. Environ. Microbiol. 1994, 60, 4142–4147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega Leal-Sánchez, M.; Ruiz-Barba, J.L.; Sánchez Gómez, A.H.; Rejano, L.; Jiménez-Dıíaz, R.; Garrido Fernández, A. Fermentation profile and optimization of green olive fermentation using Lactobacillus plantarum LPCO10 as a starter culture. Food Microbiol. 2003, 20, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Nisiotou, A.A.; Chorianopoulos, N.; Nychas, G.J.E.; Panagou, E.Z. Yeast heterogeneity during spontaneous fermentation of black Conservolea olives in different brine solutions. J. Appl. Microbiol. 2010, 108, 396–405. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Beneduce, L.; Sinigaglia, M.; Corbo, M.R. Selection of yeasts as starter cultures for table olives. J. Food Sci. 2013, 78, 742–751. [Google Scholar] [CrossRef]
- Bevilacqua, A.; De Stefano, F.; Augello, S.; Pignatiello, S.; Sinigaglia, M.; Corbo, M.R. Biotechnological innovations for table olives. Int. J. Food Sci. Nutr. 2015, 66, 127–131. [Google Scholar] [CrossRef]
- Grounta, A.; Doulgeraki, A.I.; Nychas, G.J.E.; Panagou, E.Z. Biofilm formation on Conservolea natural Black olives during single and combined inoculation with a functional Lactobacillus pentosus starter culture. Food Microbiol. 2016, 56, 35–44. [Google Scholar] [CrossRef]
- Tufariello, M.; Durante, M.; Ramires, F.A.; Grieco, F.; Tommasi, L.; Perbellini, E.; Falco, V.; Tasioula-Margari, M.; Logrieco, A.F.; Mita, G.; et al. New process for production of fermented black table olives using selected autochthonous microbial resources. Front. Microbiol. 2015, 6, 1007. [Google Scholar] [CrossRef] [Green Version]
- Bonatsou, S.; Benítez, A.; Rodríguez-Gómez, F.; Panagou, E.Z.; Arroyo-López, F.N. Selection of yeasts with multifunctional features for application as starters in natural black table olive processing. Food Microbiol. 2015, 46, 66–73. [Google Scholar] [CrossRef]
- Domínguez-Manzano, J.; Olmo-Ruiz, C.; Bautista-Gallego, J.; Arroyo-López, F.N.; Garrido Fernández, A.; Jiménez-Díaz, R. Biofilm formation on abiotic and biotic surfaces during Spanish style green table olive fermentation. Int. J. Food Microbiol. 2012, 157, 230–238. [Google Scholar] [CrossRef]
- Grounta, A.; Panagou, E.Z. Mono and dual species biofilm formation between Lactobacillus pentosus and Pichia membranifaciens on the surface of black olives under different sterile brine conditions. Ann. Microbiol. 2014, 64, 1757–1767. [Google Scholar] [CrossRef]
- Grounta, A.; Doulgeraki, A.I.; Panagou, E.Z. Quantification and characterization of microbial biofilm community attached on the surface of fermentation vessels used in green table olive processing. Int. J. Food Microbiol. 2015, 203, 41–48. [Google Scholar] [CrossRef] [PubMed]
- León-Romero, Á.; Domínguez-Manzano, J.; Garrido-Fernández, A.; Arroyo-López, F.N.; Jiménez Díaz, R. Formation of in vitro mixed-species biofilms by Lactobacillus pentosus and yeasts isolated from Spanish-style green table olive fermentations. Appl. Environ. Microbiol. 2016, 82, 689–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porru, C.; Rodríguez-Gómez, F.; Benítez-Cabello, A.; Jiménez-Díaz, R.; Zara, G.; Budroni, M.; Mannazzu, I.; Arroyo-Lopez, F.N. Genotyping, identification and multifunctional features of yeasts associated to Bosana naturally black table olive fermentations. Food Microbiol. 2018, 69, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Medina, E.; Ruiz-Bellido, M.A.; Romero-Gil, V.; Rodríguez-Gómez, F.; Montes-Borrego, M.; Landa, B.B.; Arroyo-López, F.N. Assessment of the bacterial community in directly brined Aloreña de Málaga table olive fermentations by metagenetic analysis. Int. J. Food Microbiol. 2016, 236, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Campus, M.; Sedda, P.; Cauli, E.; Piras, F.; Comunian, R.; Paba, A.; Daga, E.; Schirru, S.; Angioni, A.; Zurru, R.; et al. Evaluation of a single strain starter culture, a selected inoculum enrichment, and natural microflora in the processing of Tonda di Cagliari natural table olives: Impact on chemical, microbiological, sensory and texture quality. LWT Food Sci. Technol. 2015, 64, 671–677. [Google Scholar] [CrossRef]
- Tataridou, M.; Kotzekidou, P. Fermentation of table olives by oleuropeinolytic starter culture in reduced salt brines and inactivation of Escherichia coli O157:H7 and Listeria monocytogenes. Int. J. Food Microbiol. 2015, 208, 122–130. [Google Scholar] [CrossRef]
- Perricone, M.; Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Use of Lactobacillus plantarum and glucose to control the fermentation of “Bella di Cerignola” Table Olives, a traditional variety of Apulian region (Southern Italy). J. Food Sci. 2010, 75, 430–436. [Google Scholar] [CrossRef]
- Perricone, M.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Use of starter cultures in olives: A not-correct use could cause a delay of performances. Food Nutr. Sci. 2013, 4, 721–726. [Google Scholar] [CrossRef] [Green Version]
- Cosmai, L.; Campanella, D.; De Angelis, M.; Summo, C.; Paradiso, V.M.; Pasqualone, A.; Caponio, F. Use of starter cultures for table olives fermentation as possibility to improve the quality of thermally stabilized olive-based paste. LWT Food Sci. Technol. 2018, 90, 381–388. [Google Scholar] [CrossRef]
- Benincasa, C.; Muccilli, S.; Amenta, M.; Perri, E.; Romeo, F.V. Phenolic trend and hygienic quality of green table olives fermented with Lactobacillus plantarum starter culture. Food Chem. 2015, 186, 271–276. [Google Scholar] [CrossRef]
- Argyri, A.A.; Nisiotou, A.A.; Mallouchos, A.; Panagou, E.Z.; Tassou, C.C. Performance of two potential probiotic Lactobacillus strains from the olive microbiota as starters in the fermentation of heat shocked green olives. Int. J. Food Microbiol. 2014, 171, 68–76. [Google Scholar] [CrossRef]
- Blana, V.A.; Grounta, A.; Tassou, C.C.; Nychas, G.J.E.; Panagou, E.Z. Inoculated fermentation of green olives with potential probiotic Lactobacillus pentosus and Lactobacillus plantarum starter cultures isolated from industrially fermented olives. Food Microbiol. 2014, 38, 208–218. [Google Scholar] [CrossRef]
- Blana, V.A.; Polymeneas, N.; Tassou, C.C.; Panagou, E.Z. Survival of potential probiotic lactic acid bacteria on fermented green table olives during packaging in polyethylene pouches at 4 and 20 °C. Food Microbiol. 2016, 53, 71–75. [Google Scholar] [CrossRef]
- Ruiz-Barba, J.L.; Caballero-Guerrero, B.; Maldonado-Barragán, A.; Jiménez-Díaz, R. Coculture with specific bacteria enhances survival of Lactobacillus plantarum NC8, an autoinducer-regulated bacteriocin producer, in olive fermentations. Food Microbiol. 2010, 27, 413–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaltsa, A.; Papaliaga, D.; Papaioannou, E.; Kotzekidou, P. Characteristics of oleuropeinolytic strains of Lactobacillus plantarum group and influence on phenolic compounds in table olives elaborated under reduced salt conditions. Food Microbiol. 2015, 48, 58–62. [Google Scholar] [CrossRef]
- Lamzira, Z.; Asehraou, A.; Brito, D.; Oliveira, M.; Faid, M.; Peres, C. Bloater spoilage of greenolives. Food Technol. Biotechnol. 2005, 43, 373–377. [Google Scholar] [CrossRef] [Green Version]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozès, N. Evaluation of a single and combined inoculation of a Lactobacillus pentosus starter for processing cv. Arbequina natural green olives. Food Microbiol. 2010, 27, 731–740. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Arroyo-López, F.N.; Romero-Gil, V.; Rodríguez-Gómez, F.; Garcia-Garcia, P.; Garrido-Fernández, A. Chloride salt mixtures affect Gordal cv. green Spanish-style table olive fermentation. Food Microbiol. 2011, 28, 1316–1325. [Google Scholar] [CrossRef]
- Rodríguez-Gómez, F.; Bautista-Gallego, J.; Arroyo-López, F.N.; Romero-Gil, V.; Jiménez-Díaz, R.; Garrido-Fernández, A.; Garcia-Garcia, P. Table olive fermentation with multifunctional Lactobacillus pentosus strains. Food Control 2013, 34, 96–105. [Google Scholar] [CrossRef]
- Rodríguez-Gómez, F.; Romero-Gil, V.; Bautista-Gallego, J.; García-García, P.; Garrido-Fernández, A.; Arroyo-López, F.N. Production of potential probiotic Spanish-style green table olives at pilot plant scale using multifunctional starters. Food Microbiol. 2014, 44, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Gómez, F.; Romero-Gil, V.; Arroyo-López, F.N.; Roldán-Reyes, J.C.; Torres-Gallardo, R.; Bautista-Gallego, J.; Garcia-Garcia, P.; Garrido-Fernandez, A. Assessing the challenges in the application of potential probiotic lactic acid bacteria in the large-scale fermentation of Spanish-style table olives. Front. Microbiol. 2017, 8, 915. [Google Scholar] [CrossRef] [Green Version]
- López-López, A.; Moreno-Baquero, J.M.; Rodríguez-Gómez, F.; García-García, P.; Garrido-Fernández, A. Sensory Assessment by Consumers of Traditional and Potentially Probiotic Green Spanish-Style Table Olives. Front. Nutr. 2018, 5, 53. [Google Scholar] [CrossRef] [PubMed]
- de Castro, A.; Sánchez, A.H.; Cortés-Delgado, A.; López-López, A.; Montaño, A. Effect of Spanish-style processing steps and inoculation with Lactobacillus pentosus starter culture on the volatile composition of cv. Manzanilla green olives. Food Chem. 2019, 271, 543–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martorana, A.; Alfonzo, A.; Gaglio, R.; Settanni, L.; Corona, O.; La Croce, F.; Vagnoli, P.; Caruso, T.; Moschetti, N.; Francesca, N. Evaluation of different conditions to enhance the performances of Lactobacillus pentosus OM13 during industrial production of Spanish-style table olives. Food Microbiol. 2017, 61, 150–158. [Google Scholar] [CrossRef] [Green Version]
- Campus, M.; Cauli, E.; Scano, E.; Piras, F.; Comunian, R.; Paba, A.; Daga, E.; Di Salvo, R.; Sedda, P.; Angioni, A.; et al. Towards controlled fermentation of table olives: Lab starter driven process in an automatic pilot processing plant. Food Bioprocess Technol. 2017, 10, 1063–1073. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Todaro, A.; Pino, A.; Pitino, I.; Corona, O.; Mazzaglia, A.; Caggia, C. Giarraffa and Grossa di Spagna naturally fermented table olives: effect of starter and probiotic cultures on chemical, microbiological and sensory traits. Food Res. Int. 2014, 62, 1154–1164. [Google Scholar] [CrossRef]
- Ciafardini, G.; Zullo, B.A. Use of selected yeast starter cultures in industrial-scale processing of brined Taggiasca black table olives. Food Microbiol. 2019, 84, 103250. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Fava, G.; Tomaselli, F.; Romeo, F.V.; Pennino, G.; Vitello, E.; Caggia, C. Effect of kaolin and copper based products and of starter cultures on green table olive fermentation. Food Microbiol. 2011, 28, 910–919. [Google Scholar] [CrossRef]
- Pino, A.; De Angelis, M.; Todaro, A.; Van Hoorde, K.; Randazzo, C.L.; Caggia, C. Fermentation of Nocellara Etnea Table Olives by Functional Starter Cultures at Different Low Salt Concentrations. Front. Microbiol. 2018, 9, 1125. [Google Scholar] [CrossRef] [Green Version]
- Durante, M.; Tufariello, M.; Tommasi, L.; Lenucci, M.S.; Bleve, G.; Mita, G. Evaluation of bioactive compounds in black table olives fermented with selected microbial starters. J. Sci. Food Agric. 2017, 98, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; González-Ortiz, G.; Benítez-Cabello, A.; Calero-Delgado, B.; Jiménez-Díaz, R.; Martín-Orúe, S.M. The use of starter cultures in the table olive fermentation can modulate antiadhesive properties of brine exopolysaccharides against enterotoxigenic Escherichia coli. Food Funct. 2019, 10, 3738–3747. [Google Scholar] [CrossRef]
- Shah, P.N. Functional cultures and health benefits. Int. Dairy J. 2007, 17, 1262–1277. [Google Scholar] [CrossRef]
- Fuller, R. Probiotics in man and animal. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar] [PubMed]
- Fuller, R.; Gibson, G.R. Probiotics and prebiotics: microflora management for improved gut health. Clin. Microbiol. Infect. 1998, 4, 477–488. [Google Scholar] [CrossRef] [Green Version]
- Gorbach, S.L. Probiotics in the third millennium. Dig. Liver Dis. 2002, 34, S2–S7. [Google Scholar] [CrossRef]
- Guarner, F.; Malagelada, J.R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Holzapfel, W.H.; Schillinger, U. Introduction to pre- and probiotics. Food Res. Int. 2002, 35, 109–116. [Google Scholar] [CrossRef]
- Nagpal, R.; Kumar, A.; Kumar, M.; Behare, P.V.; Jain, S.; Yadav, H. Probiotics, their health benefits and applications for developing healthier foods: A review. FEMS Microbiol. Lett. 2002, 334, 1–15. [Google Scholar] [CrossRef] [Green Version]
- FAO. Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation; FAO: Rome, Italy, 2006; p. 85. [Google Scholar]
- Haller, D.; Colbus, H.; Ganzle, M.G.; Scherenbacher, P.; Bode, C.; Hammes, W.P. Metabolic and functional properties of lactic acid bacteria in the gastro-intestinal ecosystem: A comparative in vitro study between bacteria of intestinal and fermented food origin. Syst. Appl. Microbiol. 2001, 24, 218–226. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Varma, Y.; Fischbach, M.A.; Biddinger, S.B.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hara, A.M.; O’Regan, P.; Fanning, A.; O’Mahony, C.; Macsharry, J.; Lyons, A.; Bienenstock, J.; O’Mahony, L.; Shanahan, F. Functional modulation of human intestinal epithelial cell responses by Bifidobacterium infantis and Lactobacillus salivarius. Immunology 2006, 118, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.A.G.; Tsakalidou, E.; Nychas, G.J.E.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Arroyo-López, F.N.; Rantsiou, K.; Jiménez-Díaz, R.; Garrido-Fernández, A.; Cocolin, L. Screening of lactic acid bacteria isolated from fermented table olives with probiotic potential. Food Res. Int. 2013, 50, 135–142. [Google Scholar] [CrossRef]
- Botta, C.; Langerholc, T.; Cencic, A.; Cocolin, L. In vitro selection and characterization of new probiotic candidates from table olive microbiota. PLoS ONE 2014, 9, e94457. [Google Scholar] [CrossRef] [PubMed]
- Pérez Montoro, B.; Benomar, N.; Lavilla Lerma, L.; Castillo Gutiérrez, S.; Gálvez, A.; Abriouel, H. Fermented Aloreña table olives as a source of potential probiotic Lactobacillus pentosus strains. Front. Microbiol. 2016, 7, 1583. [Google Scholar] [CrossRef] [Green Version]
- Prete, R.; Tofalo, R.; Federici, E.; Ciarrocchi, A.; Cenci, G.; Corsetti, A. Food-associated Lactobacillus plantarum and yeasts inhibit the genotoxic effect of 4-Nitroquinoline-1-Oxide. Front. Microbiol. 2017, 8, 2349. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Gonzalez, N.; Prete, R.; Battista, N.; Corsetti, A. Adhesion Properties of Food-Associated Lactobacillus plantarum Strains on Human Intestinal Epithelial Cells and Modulation of IL-8 Release. Front. Microbiol. 2018, 9, 2392. [Google Scholar] [CrossRef]
- Prete, R.; Long, S.L.; Gallardo, A.L.; Gahan, C.G.; Corsetti, A.; Joyce, S.A. Beneficial bile acid metabolism from Lactobacillus plantarum of food origin. Sci. Rep. 2020, 10, 1165. [Google Scholar] [CrossRef] [Green Version]
- Delgado, A.; Brito, D.; Peres, C.; Noe-Arroyo, F.; Garrido-Fernández, A. Bacteriocin production by Lactobacillus pentosus B96 can be expressed as a function of temperature and NaCl concentration. Food Microbiol. 2005, 22, 521–528. [Google Scholar] [CrossRef]
- Kacem, M.; Karam, N.E. Microbiological study of naturally fermented Algerian green olives: Isolation and identification of lactic acid bacteria and yeasts along with the effects of brine solutions obtained at the end of olive fermentation on Lactobacillus plantarum growth. Grasas y Aceites 2006, 57, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Mokhbi, A.; Kaid-Harche, M.; Lamri, K.; Rezki, M.; Kacem, M. Selection of Lactobacillus plantarum strains for their use as starter culturesin Algerian olive fermentations. Grasas y Aceites 2009, 60, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Guantario, B.; Zinno, P.; Schifano, E.; Roselli, M.; Perozzi, G.; Palleschi, C.; Uccelletti, D.; Devirgiliis, C. In vitro and in vivo selection of potentially probiotic lactobacilli from Nocellara del Belice table olives. Front. Microbiol. 2018, 9, 595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benítez-Cabello, A.; Torres-Maravilla, E.; Bermúdez-Humarán, L.; Langella, P.; Martín, R.; Jiménez-Díaz, R.; Arroyo-López, F.N. Probiotic properties of Lactobacillus strains isolated from table olive biofilms. Probiotics Antimicrob. Proteins 2019. [Google Scholar] [CrossRef] [PubMed]
- Lavermicocca, P.; Valerio, F.; Lonigro, S.L.; de Angelis, M.; Morelli, L.; Callegari, M.L. Study of adhesion and survival of lactobacilli and bifidobacteria on table olives with the aim of formulating a new probiotic food. Appl. Environ. Microbiol. 2005, 71, 4233–4240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valerio, F.; de Candia, S.; Lonigro, S.L.; Russo, F.; Riezzo, G.; Orlando, A.; De Bellis, P.; Sisto, A.; Lavermicocca, P. Role of the probiotic strain Lactobacillus paracasei LMGP22043 carried by artichokes in influencing faecal bacteria and biochemical parameters in human subjects. J. Appl. Microbiol. 2011, 111, 155–164. [Google Scholar] [CrossRef]
- Argyri, A.A.; Nisiotou, A.A.; Pramateftaki, P.; Doulgeraki, A.I.; Panagou, E.Z.; Tassou, C.C. Preservation of green table olives fermented with lactic acid bacteria with probiotic potential under modified atmosphere packaging. LWT Food Sci. Technol. 2015, 62, 783–790. [Google Scholar] [CrossRef]
- Perpetuini, G.; Pham-Hoang, B.N.; Scornec, H.; Tofalo, R.; Schirone, M.; Suzzi, G.; Cavin, J.F.; Waché, Y.; Corsetti, A.; Licandro-Seraut, H. In Lactobacillus pentosus, the olive brine adaptation genes are required for biofilm formation. Int. J. Food Microbiol. 2016, 216, 104–109. [Google Scholar] [CrossRef]
- Pérez Montoro, B.; Benomar, N.; Caballero Gómez, N.; Ennahar, S.; Horvatovich, P.; Knapp, C.W.; Alonso, E.; Gálvez, A.; Abriouel, H. Proteomic analysis of Lactobacillus pentosus for the identification of potential markers of adhesion and other probiotic features. Food Res. Int. 2018, 111, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abriouel, H.; Pérez Montoro, B.; Casimiro-Soriguer, C.S.; Pérez Pulido, A.J.; Knapp, C.W.; Caballero Gómez, N.; Castillo-Gutiérrez, S.; Estudillo-Martínez, M.D.; Gálvez, A.; Benomar, N. Insight into potential probiotic markers predicted in Lactobacillus pentosus MP-10 genome sequence. Front. Microbiol. 2017, 8, 891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calero-Delgado, B.; Pérez-Pulido, A.J.; Benítez-Cabello, A.; Martín-Platero, A.M.; Casimiro-Soriguer, C.S.; Martínez-Bueno, M.; Arroyo-López, F.N.; Jiménez Díaz, R. Multiple genome sequences of Lactobacillus pentosus strains isolated from biofilms on the skin of fermented green table olives. Microbiol. Resour. Announc. 2019, 8, e01546-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, E.M.F.; Ramos, A.M.; Vanzela, E.S.L.; Stringheta, P.C.; Pinto, C.L.O.; Martins, J.M. Products of vegetable origin: A new alternative for the consumption of probiotic bacteria. Food Res. Int. 2013, 51, 764–770. [Google Scholar] [CrossRef]
- Sazawal, S.; Hiremath, G.; Dhingra, U.; Malik, P.; Deb, S.; Black, R.E. Efficacy of probiotics in prevention of acute diarrhoea: A meta-analysis of masked, randomised, placebo-controlled trials. Lancet Infect. Dis. 2006, 6, 374–382. [Google Scholar] [CrossRef]
- Pennacchia, C.; Blaiotta, G.; Pepe, O.; Villani, F. Isolation of Saccharomyces cerevisiae strains from different food matrices and their preliminary selection for a potential use as probiotics. J. Appl. Microbiol. 2008, 105, 1919–1928. [Google Scholar] [CrossRef]
- Kourelis, A.; Kotzamanidis, C.; Litopoulou-Tzanetaki, E.; Scouras, Z.G.; Tzanetakis, N.; Yiangou, M. Preliminary probiotic selection of dairy and human yeast strains. J. Biol. Res. 2010, 13, 93–104. [Google Scholar]
- Moslehi-Jenabian, S.; Lindegaard Pedersen, L.; Jespersen, L. Beneficial effects of probiotic and food borne yeasts on human health. Nutrients 2010, 2, 449–473. [Google Scholar] [CrossRef]
- Etienne-Mesmin, L.; Livrelli, V.; Privat, M.; Denis, S.; Cardot, J.M.; Alric, M.; Blanquet-Diot, S. Effect of a new probiotic Saccharomyces cerevisiae strain on survival of Escherichia coli O157:H7 in a dynamic gastrointestinal model. Appl. Environ. Microbiol. 2011, 77, 1127–1131. [Google Scholar] [CrossRef] [Green Version]
- Silva, T.; Reto, M.; Sol, M.; Peito, A.; Peres, C.M.; Peres, C.; Malcata, F.X. Characterization of yeasts from Portuguese brined olives, with a focus on their potentially probiotic behaviour. LWT Food Sci. Technol. 2011, 44, 1349–1354. [Google Scholar] [CrossRef]
- Granato, D.; Branco, G.F.; Nazzaro, F.; Cruz, A.G.; Faria, J.A.F. Functional foods and non-dairy probiotic food development: Trends, concepts, and products. Compr. Rev. Food Sci. Food Saf. 2010, 9, 292–302. [Google Scholar] [CrossRef]
- Vijaya Kumar, B.; Vijayendra, S.V.N.; Reddy, O.V.S. Trends in dairy and non-dairy probiotic products—A review. J. Food Sci. Technol. 2015, 52, 6112–6124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranadheera, S.; Baines, S.K.; Adams, M.C. Importance of food in probiotic efficacy. Food Res. Int. 2010, 43, 1–7. [Google Scholar] [CrossRef]
- Lamsal, B.P.; Faubion, J. The beneficial use of cereal and cereal components in probiotic foods. Food Rev. Int. 2009, 25, 103–114. [Google Scholar] [CrossRef]
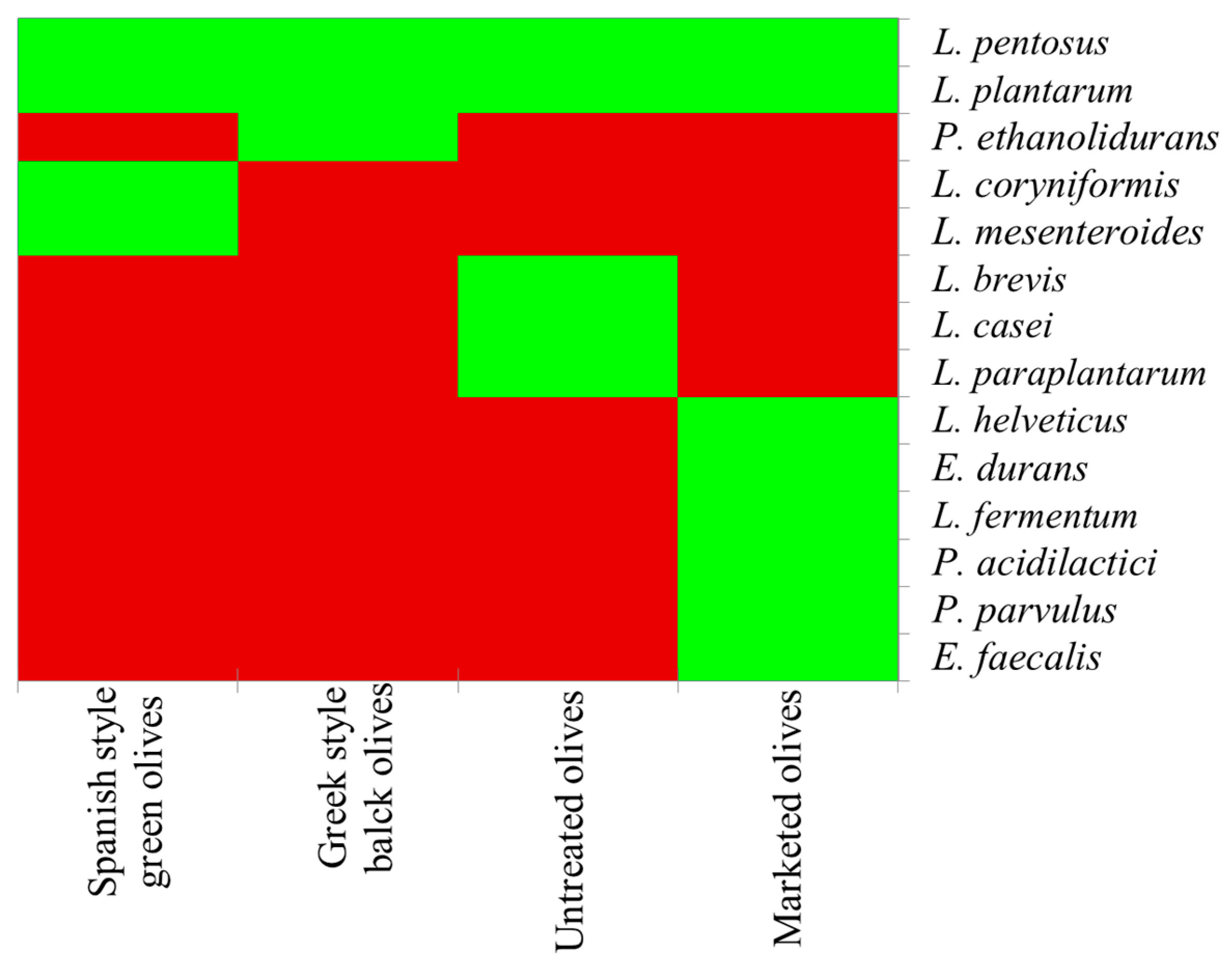
Figure 1.
LAB species detected in table olives. The green color indicates the presence of the species, while red its absence. Spanish-style olives are debittered through the addition of lye. In the Greek style, olives are put directly in brine and oleuropein is removed by the enzymatic activities of indigenous microorganisms.
Figure 1.
LAB species detected in table olives. The green color indicates the presence of the species, while red its absence. Spanish-style olives are debittered through the addition of lye. In the Greek style, olives are put directly in brine and oleuropein is removed by the enzymatic activities of indigenous microorganisms.


{kind=link}
Table 1.
Olive processing methods according to the International Olive Oil Council (IOOC) [1].
Table 1.
Olive processing methods according to the International Olive Oil Council (IOOC) [1].
| Preparation Method | Process |
|---|---|
| Treated olives | It is applied to green olives, olives turning color, or black olives. Olive debittering is achieved through an alkaline treatment (lye 2.5%–3% w/v). Olives are then placed in brine (NaCl 10%–11% w/v) where the fermentation takes place and lasts 3–7 months. Fermentation is driven by lactic acid bacteria. |
| Natural olives | It is applied to green olives, olives turning color. or black olives. Olives are placed directly in brine. With a salt concentration of about 6%–10% (w/v). Oleuropein is removed through the enzymatic activities (mainly β-glucosidase and esterase) of indigenous microorganisms. The fermentation process can last 8–12 months and it is mainly driven by yeasts and lactic acid bacteria. |
| Dehydrated and/or shriveled olives | It is applied to green olives, olives turning color, or black olives. Olives are subjected or not to a mild alkaline treatment, preserved in brine, or partially dehydrated in dry salt and/or by heating. |
| Olives darkened by oxidation | It is applied to green olives or olives turning color. Olives are preserved in brine, fermented or not, and darkened by oxidation in an alkaline medium. They are stored in hermetically sealed containers and subjected to heat sterilization. |
| Specialties | Olives prepared in a different way than those above following traditional recipes. |
Table 2.
Characteristics and advantages in the use of selected starter cultures.
| Properties | Characteristics | Advantages |
|---|---|---|
| Safety | Safe and stable activity Standardized activity Easy to manage and reproduce | Reproducibility Controlled and stable fermentation Continuous monitoring of fermentation |
| Technological | Ability to colonize olives surface (i.e., biofilm formation) Low demand for nutrients | Rapid and predominant growth High adaptation ability |
| Growth at different pH (high/low) Salt tolerance Ability to survive/growth at low temperatures | Dominance during the fermentation | |
| Biodegradation of phenolic compounds Debittering activity (i.e., oleuropeinolytic activity) High acidification activity | Reduction of fermentation time Avoided use of chemicals (microbial biotransformation) | |
| Functional | Antimicrobial activity vs. pathogens (i.e., bacteriocins production, competitive action on nutrients) Biocontrol agents vs. spoilage microorganisms (i.e., production of killer factors) | Protection from undesirable and/or pathogenic microorganisms Improvement of final product stability and shelf-life extension |
| Enzymatic activities (i.e., lipase, alkaline/acid phosphatase, β-glucosidase) Vitamins production Production of aromatic compounds | Enhancement of organoleptic, nutritional and sensory profile of the final product | |
| Probiotic | Survival under gastrointestinal conditions (i.e., low pH, gastric and pancreatic digestion, bile salts) Ability to adhere and persist in the intestinal mucosa Modulation of host immune system Antimicrobial activity against pathogens | Ensuring product safety Quality enhancement of the final product Production of a health-promoting functional food |
Table 3.
Main starter strains used for table olive fermentation.
| Bacterial Starter Cultures | Cultivar | References |
|---|---|---|
| L. plantarum | Alorena | [40] |
| Bella di Cerignola | [33,63,64,65] | |
| Carolea/Cassanese | [66] | |
| Conservolea | [41] | |
| Gordal | [40] | |
| Halkidiki | [67,68,69] | |
| Hojiblanca | [40,70] | |
| Kalamata/Chalkidikis | [62,71] | |
| Manzanilla | [40] | |
| Mele | [28] | |
| Nocellara del Belice/Nocellara Messinese | [66] | |
| Nocellara Etnea | [34] | |
| Picholine | [72] | |
| Pishomi | [42] | |
| Tonda di Cagliari | [39,61] | |
| Leccino | [44] | |
| L. pentosus | Arbequina | [73] |
| Conservolea | [41,52] | |
| Gordal | [55,74] | |
| Halkidiki | [67,68,69] | |
| Itrana | [15,35] | |
| Manzanilla | [40,75,76,77,78,79] | |
| Nocellara del Belice | [36,80] | |
| Nocellara Etnea | [34] | |
| Tonda di Cagliari | [39,61,81] | |
| L. paracasei | Bella di Cerignola | [9] |
| L. rhamnosus | Giaraffa e Grossa di Spagna | [82] |
| Yeast starter cultures | Cultivar | References |
| N. molendini-olei/C. matritensis/C. adriatica/C. diddensiae/W. anomalus/S. cerevisiae | Taggiasca | [83] |
Table 4.
Main multi-starter strains used for table olive fermentation.
| Multi-starter Cultures | Cultivar | References |
|---|---|---|
| L. plantarum/L. pentosus | Bella di Cerignola | [65] |
| Halkidiki | [67,68] | |
| Nocellara Etnea | [34] | |
| L. plantarum/L. casei | Nocellara Etnea | [84] |
| L. plantarum/L. paracasei | Giaraffa e Grossa di Spagna | [82] |
| L. plantarum/L. paracasei | Nocellara Etnea | [43] |
| Nocellara Etnea | [34,85] | |
| L. plantarum/P. pentosaceus | Green olives | [70] |
| L. plantarum/E. faecieum | Green olives | [70] |
| L. paracasei/L. pentosus | Nocellara Etnea | [34] |
| L. pentosus/L. coryniformis | Nocellara del Belice | [12] |
| L. plantarum/L. paracasei/L. rhamnosus | Giaraffa e Grossa di Spagna | [82] |
| L. plantarum/L. paracasei/L. pentosus | Nocellara Etnea | [34] |
| L. plantarum/D. hansenii | Conservolea | [53,86] |
| L. plantarum/C. famata/C. guilliermondii | Bella di Cerignola | [64] |
| L. plantarum/S. cerevisiae | Leccino | [53,86] |
| L. plantarum/W. anomalus | Cellina di Nardò | [53,86] |
| L. plantarum/W. anomalus | Bella di Cerignola | [33,65] |
| L. plantarum/W. anomalus/L. pentosus | Bella di Cerignola | [33] |
| L. pentosus/P. membranifaciens | Conservolea | [52,53] |
| L. pentosus/C. boidinii | Manzanilla | [87] |
| L. mesenteroides/S. cerevisiae | Kalamata | [53,86] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Perpetuini, G.; Prete, R.; Garcia-Gonzalez, N.; Khairul Alam, M.; Corsetti, A. Table Olives More than a Fermented Food. Foods 2020, 9, 178. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9020178
AMA Style
Perpetuini G, Prete R, Garcia-Gonzalez N, Khairul Alam M, Corsetti A. Table Olives More than a Fermented Food. Foods. 2020; 9(2):178. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9020178
Chicago/Turabian StylePerpetuini, Giorgia, Roberta Prete, Natalia Garcia-Gonzalez, Mohammad Khairul Alam, and Aldo Corsetti. 2020. "Table Olives More than a Fermented Food" Foods 9, no. 2: 178. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9020178
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.