Mosquito Surveillance for Prevention and Control of Emerging Mosquito-Borne Diseases in Portugal — 2008–2014


Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Collection
2.2. Ports and Airports
2.3. Flavivirus Survey
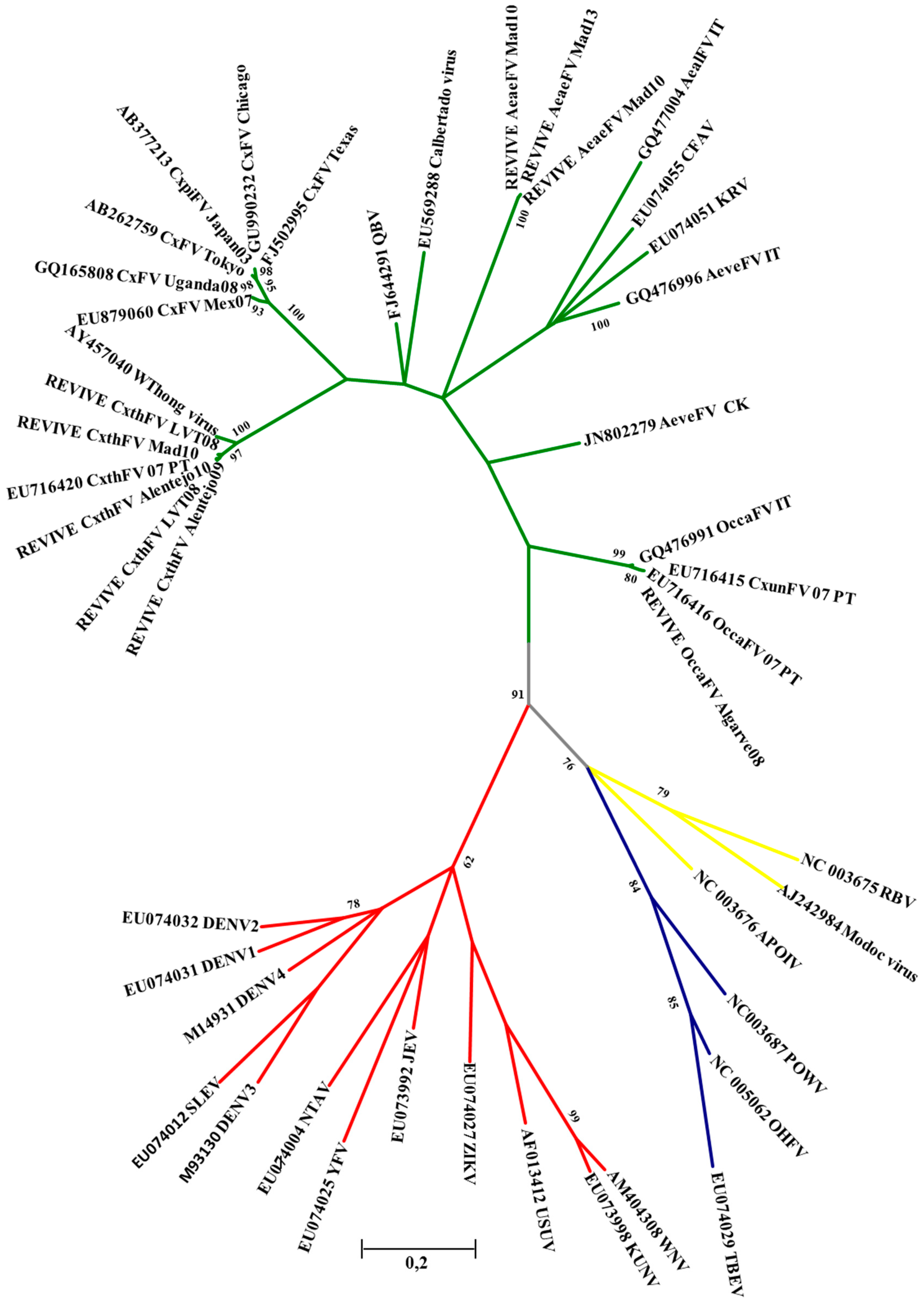
3. Results


{kind=link}
{kind=link}
| Collections | Number of | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | Total |
|---|---|---|---|---|---|---|---|---|
| Adult Collection | CDC Trap-night | 267 | 227 | 464 | 725 | 719 | 933 | 3335 |
| Adult mosquitoes | 18,310 | 23,336 | 25,790 | 12,739 | 21,127 | 15,506 | 116,808 | |
| Species number | 15 | 13 | 13 | 22 | 17 | 19 | 22 | |
| Sub-regions (NUTS III) | 19 | 11 | 15 | 19 | 20 | 20 | 20 | |
| Immature Collection | Surveyed breeding sites | 143 | 59 | 166 | 518 | 697 | 2035 | 3618 |
| Immature mosquitoes | 3338 | 2125 | 4562 | 14,526 | 25,988 | 23,850 | 74,389 | |
| Species number | 8 | 7 | 9 | 12 | 10 | 13 | 17 | |
| Sub-regions (NUTS III) | 14 | 9 | 13 | 19 | 20 | 20 | 20 |
| Species | Larvae | ♂ 1 | ♀ 2 | ♂ & ♀ 3 | Pools 4 | ♀ Pools 5 | % ♀ Pools 6 |
|---|---|---|---|---|---|---|---|
| Aedes aegypti | 2173 | 606 | 195 | 801 | 53 | 725 | 1.82 |
| Ae. eatoni | 8 | 2 | 2 | 0.00 | |||
| Anopheles algeriensis | 6 | 437 | 443 | 7 | 278 | 0.70 | |
| An. claviger | 16 | 110 | 126 | 5 | 54 | 0.14 | |
| An. maculipennis s.l. | 295 | 124 | 645 | 769 | 44 | 404 | 1.01 |
| An. plumbeus | 8 | 8 | 0.00 | ||||
| Coquillettidea richiardii | 75 | 75 | 6 | 55 | 0.14 | ||
| Culiseta annulata | 140 | 3 | 237 | 240 | 10 | 62 | 0.16 |
| Cs. longiareolata | 29,672 | 570 | 439 | 1009 | 15 | 52 | 0.13 |
| Culex hortensis | 1578 | 4 | 13 | 17 | 2 | 5 | 0.01 |
| Cx. impudicus | 120 | 3 | 3 | 0.00 | |||
| Cx. laticinctus | 1908 | 54 | 25 | 79 | 0.00 | ||
| Cx. mimeticus | 3 | 3 | 8 | 11 | 1 | 6 | 0.02 |
| Cx. modestus | 1 | 1 | 225 | 226 | 6 | 168 | 0.42 |
| Cx. perexiguus | 67 | 144 | 1303 | 1447 | 79 | 929 | 2.33 |
| Cx. pipiens | 36,531 | 4727 | 34,214 | 38,941 | 696 | 19,181 | 48.11 |
| Cx. territans | 50 | 0.00 | |||||
| Cx. theileri | 1300 | 648 | 30,197 | 30,845 | 241 | 8037 | 20.16 |
| Cx. torrentium | 359 | 0.00 | |||||
| Ochlerotatus berlandi | 9 | 9 | 2 | 17 | 0.04 | ||
| Oc. caspius | 12 | 1038 | 29,521 | 30,559 | 209 | 8456 | 21.21 |
| Oc. detritus | 172 | 409 | 10,553 | 10,962 | 40 | 1409 | 3.53 |
| Oc. geniculatus | 55 | 55 | 2 | 21 | 0.05 | ||
| Uranotaenia unguiculata | 114 | 67 | 181 | 1 | 13 | 0.03 | |
| Total | 74,389 | 8470 | 108,338 | 116,808 | 1419 | 39,872 | 100.00 |
| Points of Entry | Collections | N | Ae. aegypti | An. maculipennis | Cx. hortensis | Cx. pipiens | Cs. annulata | Cs. longiareolata | Cx. theileri | Oc. caspius | Oc. detritus | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IM 1 | AD 2 | IM | AD | IM | IM | AD | IM | IM | AD | IM | AD | IM | AD | AD | AD | IM | AD | |
| Aveiro | 13 | 62 | 55 | 96 | 1 | 37 | 55 | 14 | 43 | 1 | ||||||||
| Faro-Airport | 400 | 55 | 725 | 1934 | 4 | 12 | 599 | 338 | 6 | 139 | 1025 | 139 | ||||||
| Figueira da Foz | 6 | 63 | 240 | 10,206 | 3 | 240 | 863 | 9 | 230 | 9029 | 72 | |||||||
| Funchal (Madeira Island) | 1 | 20 | 20 | |||||||||||||||
| Machico (Madeira Island) | 1 | 7 | 7 | |||||||||||||||
| Porto-Airport | 1 | 41 | 7 | 7 | ||||||||||||||
| Matosinhos | 8 | 123 | 111 | 1378 | 10 | 1371 | 1 | 101 | 4 | 2 | ||||||||
| Portimão | 55 | 52 | 141 | 308 | 100 | 196 | 41 | 63 | 7 | 40 | ||||||||
| Setúbal | 228 | 6 | 1 | 1 | ||||||||||||||
| Sines | 23 | 15 | 423 | 32 | 1 | 23 | 16 | 3 | 395 | 4 | 4 | 5 | 4 | |||||
| Viana do Castelo | 46 | 11 | 702 | 22 | 44 | 293 | 1 | 74 | 291 | 1 | 20 | |||||||
| Vila Real de Santo António | 148 | 483 | 201 | 282 | ||||||||||||||
| Total | 930 | 428 | 2907 | 13,984 | 20 | 1 | 44 | 879 | 3091 | 74 | 4 | 1510 | 101 | 382 | 10,162 | 4 | 212 | |
| Counties | Collections | N | Ae. aegypti | Ae. eatoni | Cs. longiareolata | Cx. pipiens | Cx. theileri | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IM | AD | IM | AD | IM | AD | AD | IM | AD | IM | AD | IM | AD | |
| Calheta | 20 | 1 | 141 | 23 | 136 | 23 | 5 | 20 | |||||
| Câmara de Lobos | 33 | 123 | 805 | 464 | 715 | 338 | 1 | 82 | 31 | 8 | 88 | 33 | 6 |
| Funchal | 161 | 358 | 4154 | 984 | 1295 | 440 | 1 | 261 | 159 | 1967 | 270 | 161 | 114 |
| Machico | 4 | 57 | 57 | 4 | |||||||||
| Ponta do Sol | 1 | 14 | 12 | 2 | 1 | ||||||||
| Ribeira Brava | 2 | 8 | 2 | ||||||||||
| Santa Cruz | 6 | 77 | 15 | 51 | 11 | 6 | |||||||
| Total | 227 | 482 | 5256 | 1471 | 2173 | 801 | 2 | 451 | 190 | 1993 | 358 | 227 | 120 |
| Number of | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | Total |
|---|---|---|---|---|---|---|---|
| Pools | 214 | 164 | 230 | 328 | 251 | 232 | 1419 |
| ♀ Pools | 6785 | 5230 | 5093 | 7425 | 7947 | 7392 | 39,872 |
| Culex FV | 5 | 0 | 3 | 7 | 0 | 0 | 15 |
| Ochlerotatus FV | 1 | 1 | 1 | 4 | 0 | 0 | 7 |
| Ae. aegypti FV | 0 | 0 | 3 | 7 | 0 | 4 | 14 |

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Petric, D.; Bellini, R.; Scholte, E.-J.; Rakotoarivony, L.M.; Schaffner, F. Monitoring population and environmental parameters of invasive mosquito species in Europe. Parasit. Vectors 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Guidelines for the Surveillance of Invasive Mosquitoes in Europe; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2012.
- Angelini, R.; Finarelli, A.C.; Angelini, P.; Po, C.; Petropulacos, K.; Silvi, G.; Macini, P.; Fortuna, C.; Venturi, G.; Magurano, F.; et al. Chikungunya in north-eastern Italy: A summing up of the outbreak. Euro Surveill. 2007, 12. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=3313 (accessed on 4 November 2014).
- Grandadam, M.; Caro, V.; Plumet, S.; Thiberge, J.M.; Souares, Y.; Failloux, A.B.; Tolou, H.J.; Budelot, M.; Cosserat, D.; Leparc-Goffart, I.; et al. Chikungunya virus, southeaster France. Emerg. Infect. Dis. 2011, 17, 910–913. [Google Scholar] [CrossRef] [PubMed]
- La Ruche, G.; Souares, Y.; Armengaud, A.; Peloux-Petiot, F.; Delaunay, P.; Despres, P.; Lenglet, A.; Jourdain, F.; Leparc-Goffart, I.; Charlet, F.; et al. First two autochthonous dengue virus infections in metropolitan France, September 2010. Eur. Surveill. 2010, 15. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19676 (accessed on 4 November 2014).
- Gjenero-Margan, I.; Aleraj, B.; Krajcar, D.; Lesnikar, V.; Klobučar, A.; Pem-Novosel, I.; Kurečić-Filipović, S.; Komparak, S.; Martić, R.; Duričić, S.; et al. Autochthonous dengue fever in Croatia, August–September 2010. Eur. Surveill. 2011, 16. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19805 (accessed on 4 November 2014).
- Alves Alves, M.J.; Fernandes, P.L.; Amaro, F.; Osório, H.; Luz, T.; Parreira, P.; Andrade, G.; Zé-Zé, L.; Zeller, H. Clinical presentation and laboratory findings for the first autochthonous cases of dengue fever in Madeira island, Portugal, October 2012. Eur. Surveill. 2013, 18. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=20398 (accessed on 4 November 2014).
- Autochonous Dengue Cases in Madeira, Portugal; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2012.
- Schäfer, M.; Lundström, J.; Pfeffer, M.; Lundkvist, E.; Landin, J. Biological diversity vs. risk for mosquito nuisance and disease transmission in constructed wetlands in southern Sweden. Med. Vet. Entomol. 2004, 18, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Schaffner, F.; Bellini, R.; Petrić, D.; Scholte, E.-J.; Zeller, H.; Rakotoarivony, L.M. Development of guidelines for the surveillance of invasive mosquitoes in Europe. Parasit. Vectors 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Pervanidou, D.; Detsis, M.; Danis, K.; Mellou, K.; Papanikolaou, E.; Terzaki, I.; Baka, A.; Veneti, L.; Vakali, A.; Dougas, G.; et al. West Nile virus outbreak in humans, Greece, 2012: Third consecutive year of local transmission. Eur. Surveill. 2014, 19. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=20758 (accessed on 4 November 2014).
- Rosà, R.; Marini, G.; Bolzoni, L.; Neteler, M.; Metz, M.; Deluchi, L.; Chadwick, E.; Balbo, L.; Mosca, A.; Giacobini, M.; et al. Early warning of West Nile virus mosquito vector: Climate and land use models successfully explain phenology and abundance of Culex pipiens mosquitoes in north-western Italy. Parasit. Vectors 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Arends, J.E.; Oosterheert, J.J.; Kraaij-Dirkswager, M.M.; Kaan, J.A.; Fanoby, E.B.; Haas, P.J.; Scholte, E.J.; Kortbeek, L.M.; Sankatsing, S.U. Two cases of Plasmodium falciparum malaria in the Netherlands without recent travel to a malaria-endemic country. Amer. J. Trop. Med. Hyg. 2013, 13. [Google Scholar] [CrossRef]
- Doudier, B.; Bogreau, H.; DeVries, A.; Ponçon, N.; Stauffer, W.M.; Fontenille, D.; Rogier, C.; Parola, P. Possible autochthonous malaria from Marseille to Minneapolis. Emerg. Infect. Dis. 2007, 13, 1236–1238. [Google Scholar] [CrossRef]
- Ioannidis, A.; Nicolaou, C.; Stoupi, A.; Kossyvakis, A.; Matsoukas, P.; Liakata, M.V.; Magiorkinis, E.; Petinaki, E.; Chatzpanagiotou, S. First report of a phylogenetic analysis of an autochthonous Plasmodium vivax strain isolated from a malaria case in east Attica, Greece. Malar. J. 2013, 12. [Google Scholar] [CrossRef] [PubMed]
- Santa-Olalla, P.; Vazquez-Torres, M.C.; Latorre-Fandos, E.; Mairal-Claver, P.; Cortina-Solano, P.; Puy-Azón, A.; Sancho, A.B.; Leitmeyer, K.; Lucientes-Curdi, J.; Sierra-Moros, M.J. First autochthonous malaria case due to Plasmodium vivax since eradication, Spain, October 2010. Eur. Surveill. 2010, 15. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19684 (accessed on 4 November 2014).
- Diário da República Eletrónico. International Health Regulations, 2005. Available online: http://www.dre.pt/pdf1sdip/2008/01/01600/0063800687.PDF (accessed on 20 August 2014).
- Semenza, J.C.; Zeller, H. Integrated surveillance for prevention and control of emerging vector-borne diseases in Europe. Euro Surveill. 2014, 19. Available online: http://www.eurosurveillance.org/images/dynamic/EE/V19N13/art20757.pdf (accessed on 4 November 2014).
- Word Health Organization. Alert, Response, and Capacity Building under the International Health Regulations (IHR). Available online: http://www.who.int/ihr/ports_airports/ihr_authorized_ports_list.pdf?ua=1 (accessed on 20 August 2014).
- Alves, M.J.; Osório, H.C.; Zé-Zé, L.; Amaro, F. Relatório REVIVE 2008/2009—Programa Nacional de Vigilância de Vectores Culicídeos; Instituto Nacional de Saúde Dr. Ricardo Jorge: Lisboa, Portugal, 2010. (in Portuguese) [Google Scholar]
- Alves, M.J.; Osório, H.C.; Amaro, F.; Zé-Zé, L. Relatório REVIVE 2013—Programa Nacional de Vigilância de Vectores Culicídeos; Instituto Nacional de Saúde Dr. Ricardo Jorge: Lisboa, Portugal, 2013. (in Portuguese) [Google Scholar]
- Ribeiro, H.; Ramos, H.C. Identification keys of the mosquitoes of continental Portugal, Açores and Madeira. Eur. Mosq. Bull. 1999, 3, 1–11. [Google Scholar]
- Schaffner, E.; Anges, G.; Geoffroy, B.; Hervy, J.-P.; Rhaiem, A.; Brunhes, J. The Mosquitoes of Europe: An Identification and Training Programme; IRD Editions: Montpellier, France, 2001. [Google Scholar]
- Brise, T.; Jia, X.Y.; Huang, C.; Grady, L.J.; Lipkin, W.I. Identification of a Kunjin/West Nile-like flavivirus in brains of patients with New York encephalitis. Lancet 1999, 354, 1261–1262. [Google Scholar] [CrossRef] [PubMed]
- Briese, T.; Rambaut, A.; Pathmajeyan, M.; Bishara, J.; Weinberg, M.; Pitlik, S.; Lipkin, W.I. Phylogenetic analysis of a human isolate from the 2000 Israel West Nile virus epidemic. Emerg. Infect. Dis. 2002, 8, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffner, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Igarashi, A.; Harrap, K.A.; Casals, J.; Stollar, V. Morphological, biochemical, and serological studies on a viral agent. Virology 1976, 74, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Wilder-Smith, A.; Quam, M.; Sessions, O.; Rocklov, J.; Liu-Helmersson, J.; Franco, L.; Khan, K. The 2012 dengue outbreak in Madeira: Exploring the origins. Eur. Surveill. 2014, 19. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=20718 (accessed on 4 November 2014).
- Filipe, A.R.; Pinto, M.R. Isolation in Portugal of West Nile virus from Anopheles. Acta Virol. 1972, 16, 361. [Google Scholar] [PubMed]
- Filipe, A.R.; Pinto, M.R. Survey for antibodies to arboviruses in serum of animals from southern Portugal. Amer. J. Trop. Med. Hyg. 1969, 18, 423–426. [Google Scholar]
- Connell, J.; McKeown, P.; Garvey, P.; Cotter, S.; Conway, A.; O’Flanagan, D.; O’Herlihy, B.P.; Morgan, D.; Nicoll, A.; Lloyd, G. Two linked cases of West Nile virus (WNV) acquired by Irish tourists in the Algarve, Portugal. Eur. Surveill. 2004, 8. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=2517 (accessed on 4 November 2014).
- Parreira, R.; Severino, P.; Freitas, F.; Piedade, J.; Almeida, A.P.; Esteves, A. Two distinct introductions of the West Nile virus in Portugal disclosed by phylogenetic analysis of genomic sequences. Vector Borne Zoonotic Dis. 2007, 7, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Barros, S.C.; Ramos, F.; Fagulha, T.; Duarte, M.; Henriques, M.; Luís, T.; Fevereiro, M. Serological evidence of West Nile virus circulation in Portugal. Vet. Microbiol. 2011, 152, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.J.; Poças, J.M.; Luz, T.; Amaro, F.; Zé-Zé, L.; Osório, H.C. West Nile virus infection in Portugal: Considerations about a clinical case with febrile syndrome and rash. Rev. Port. Doenças Infec. 2012, 8, 46–51. [Google Scholar]
- Calzolari, M.; Zé-Zé, L.; Růžek, D.; Vázquez, A.; Jeffries, C.; Defilippo, F.; Osório, H.C.; Kilian, P.; Ruíz, S.; Fook, A.R.; et al. Detection of mosquito-only flaviviruses in Europe. J. Gen. Virol. 2012, 93, 1215–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phybus, O.G.; Rambaut, A.; Holmes, E.C.; Harvey, P.H. New inferences from tree shape: Numbers of missing taxa and population growth rates. Syst. Biol. 2002, 51, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Bennett, S.N.; Holmes, E.C.; de Chesse, R.; Moureau, G.; de Lamballerie, X. Isolation of a new strain of the flavivirus cell fusing agent virus in a natural mosquito population from Puerto Rico. J. Gen. Virol. 2006, 87, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Moureau, G.; Temmam, S.; Gonzalez, J.P.; Charrel, R.N.; Grard, G.; de Lamballerie, X. Real-time RT-PCR method for the universal detection and identification of flaviviruses. Vector Borne Zoonotic Dis. 2007, 7, 467–477. [Google Scholar] [CrossRef] [PubMed]
- De Lamballerie, X.; Crochu, S.; Billoir, F.; Neyts, J.; de Micco, P.; Holmes, E.C.; Gould, E.A. Genome sequence analysis of Tamana bat virus and its relationship with the genus Flavivirus. J. Gen. Virol. 2002, 83, 2443–2454. [Google Scholar] [PubMed]
- Bolling, B.G.; Olea-Popelka, F.J.; Eisen, L.; Moore, C.G.; Blair, C.D. Transmission dynamics of an insect-specific flavivirus in a naturally infected Culex pipiens laboratory colony and effects of co-infection on vector competence for West Nile virus. Virology 2012, 427, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Crochu, S.; Cook, S.; Attoui, H.; Charrel, R.N.; de Chesse, R.; Belhouchet, M.; Lemasson, J.-J.; Micco, P.; Lamballerie, X. Sequences of flavivurus-related RNA viruses persist in DNA form integrated in the genome of Aedes spp. mosquitoes. J. Gen. Virol. 2004, 85, 1971–1980. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Moureau, G.; Kitchen, A.; Gould, E.A.; Lamballerie, X.; Holmes, E.C.; Harbach, R.E. Molecular evolution of the insect-specific flaviviruses. J. Gen. Virol. 2012, 93, 223–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, F.; Cerutti, F.; Ballardini, M.; Mosca, A.; Vitale, N.; Radaelli, M.C.; Desiato, R.; Prearo, M.; Pautasso, A.; Casalone, C.; et al. Molecular characterization of flaviviruses from field-collected mosquitoes in northwestern Italy, 2011–2012. Parasit. Vectors 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Hobson-Peters, J.; Yam, A.W.; Lu, J.W.; Setoh, Y.X.; May, F.J.; Kurucz, N.; Walsh, S.; Prow, N.A.; Davis, S.S.; Weir, R.; et al. A new insect-specific flavivirus from northern Australia suppresses replication of West Nile virus and Murray Valley encephalitis virus in co-infected mosquito cells. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Kent, R.J.; Crabtree, M.B.; Miller, B.R. Transmission of West Nile virus by Culex quinquefasciatus say infected with Culex Flavivirus Izabal. PLoS Negl. Trop. Dis. 2010, 4. [Google Scholar] [CrossRef] [PubMed]
- Newman, C.M.; Cerutti, F.; Anderson, T.K.; Hamer, G.L.; Walker, E.D.; Kitron, U.D.; Ruiz, M.O.; Brawn, J.D.; Goldberg, T.L. Culex flavivirus and West Nile virus mosquito coinfection and positive ecological association in Chicago, United States. Vector Borne Zoonotic Dis. 2011, 11, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osório, H.C.; Zé-Zé, L.; Amaro, F.; Alves, M.J. Mosquito Surveillance for Prevention and Control of Emerging Mosquito-Borne Diseases in Portugal — 2008–2014. Int. J. Environ. Res. Public Health 2014, 11, 11583-11596. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph111111583
Osório HC, Zé-Zé L, Amaro F, Alves MJ. Mosquito Surveillance for Prevention and Control of Emerging Mosquito-Borne Diseases in Portugal — 2008–2014. International Journal of Environmental Research and Public Health. 2014; 11(11):11583-11596. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph111111583
Chicago/Turabian StyleOsório, Hugo C., Líbia Zé-Zé, Fátima Amaro, and Maria J. Alves. 2014. "Mosquito Surveillance for Prevention and Control of Emerging Mosquito-Borne Diseases in Portugal — 2008–2014" International Journal of Environmental Research and Public Health 11, no. 11: 11583-11596. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph111111583