Endpoint Visual Detection of Three Genetically Modified Rice Events by Loop-Mediated Isothermal Amplification

Abstract
:1. Introduction
2. Results and Discussion
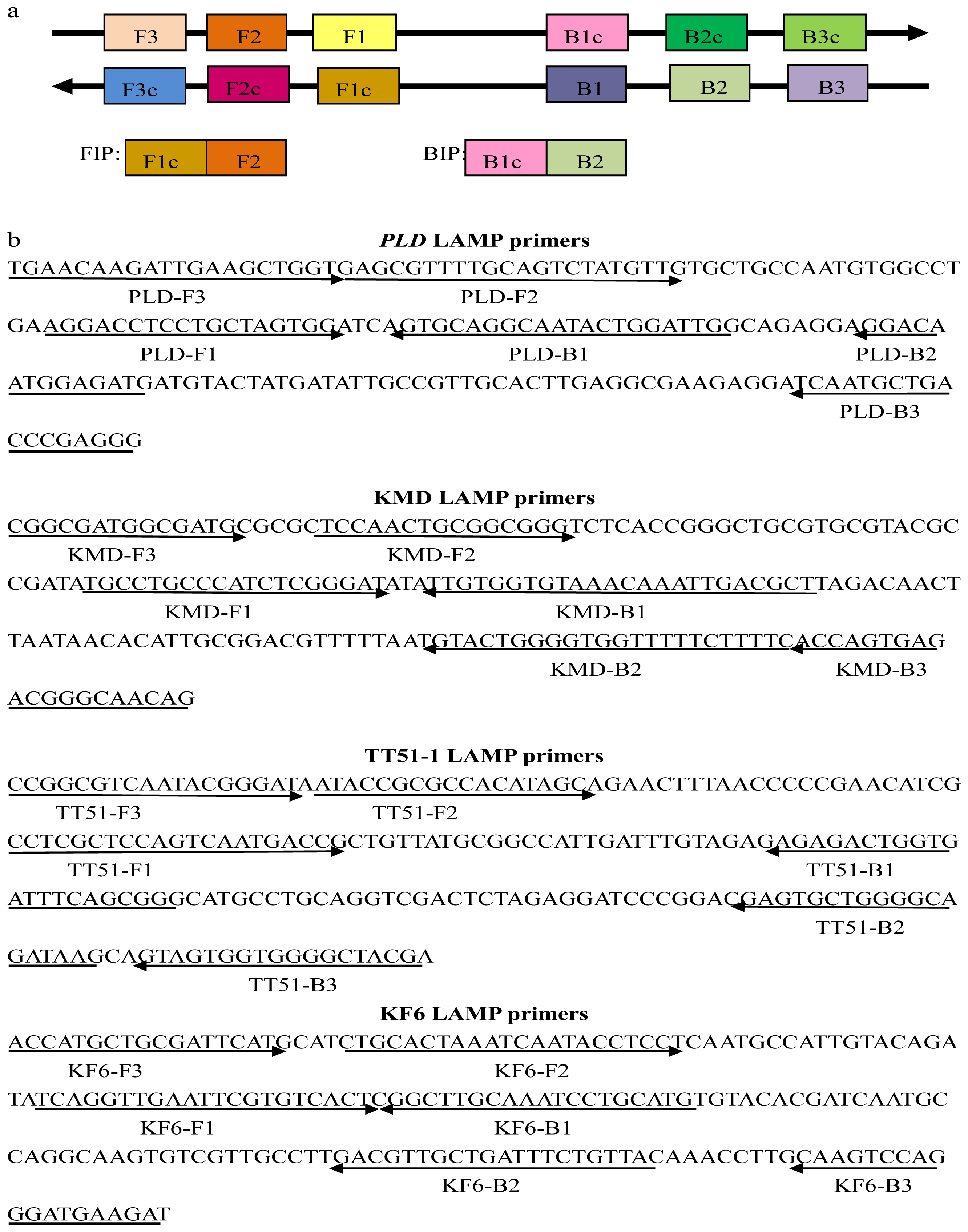
2.1. Primer Design for the Target Sequences
2.2. Optimization of the LAMP Reactions
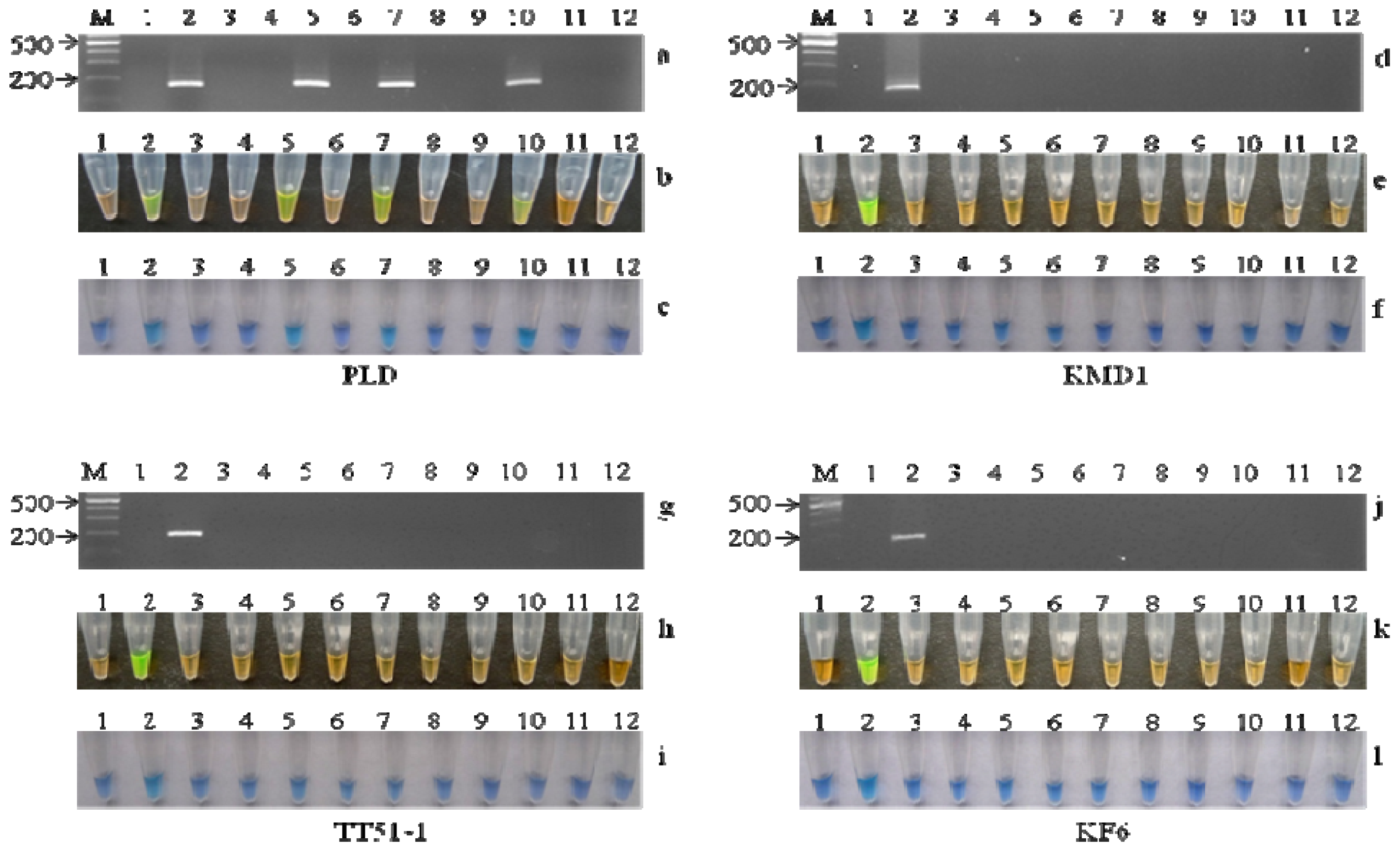
2.3. Specificity of LAMP
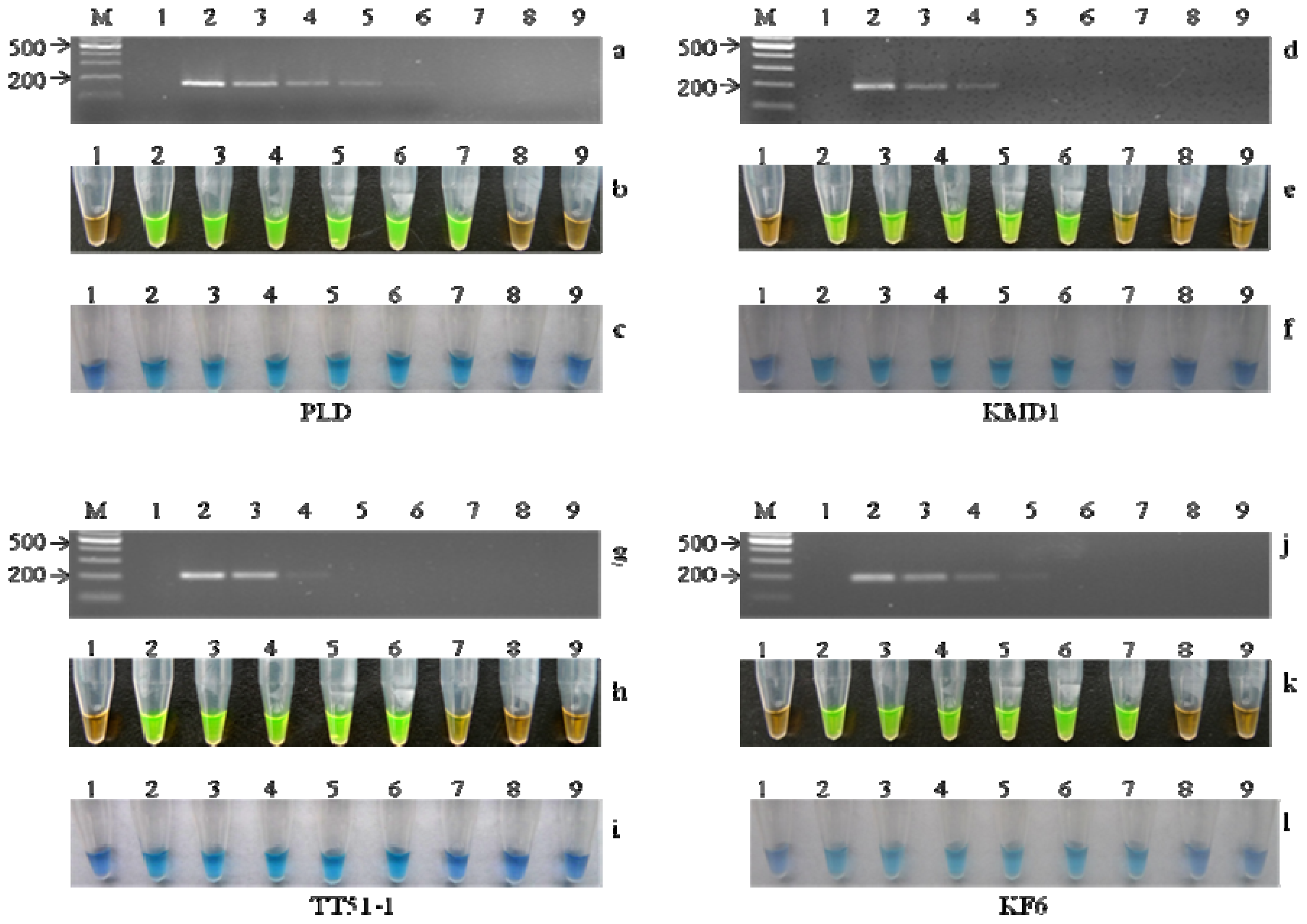
2.4. Sensitivity of LAMP
3. Experimental Section
3.1. Plant Materials
3.2. DNA Extraction
3.3. Primer Design
3.4. Conventional PCR
3.5. LAMP Assay
4. Conclusions
Acknowledgments
References
- James, C. Global Status of Commercialized Biotech./GM Crops; ISAAA Brief No. 43; International Service for the Acquisition of Agri-biotech Applications (ISAAA): Ithaca, NY, USA, 2011. [Google Scholar]
- Shimamoto, K.; Terada, R.; Izawa, T.; Fujimoto, H. Fertile transgenic rice plants regenerated from transformed protoplasts. Nature 1989, 338, 274–276. [Google Scholar]
- Datta, S.K.; Datta, K.; Soltanifar, N.; Donn, G.; Potrykus, I. Herbicide-resistant Indica rice plants from IRRI breeding line IR72 after PEG-mediated transformation of protoplasts. Plant Mol. Biol 1992, 20, 619–629. [Google Scholar]
- Oard, J.H.; Linscombe, S.D.; Braverman, M.P.; Jodari, F.; Blouin, D.C.; Leech, M.; Kohli, A.; Vain, P.; Cooley, J.C.; Christou, P. Development, feld evaluation and agronomic performance of transgenic herbicide resistant rice. Mol. Breed 1996, 2, 359–368. [Google Scholar]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.S.; Pi, L.Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.X.; Zhu, L.H.; et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar]
- Zhai, W.X.; Chen, C.Y.; Zhu, X.F.; Chen, X.W.; Zhang, D.H.; Li, X.B.; Zhu, L.H. Analysis of tDNA-Xa21 loci and bacterial blight resistance effects of the transgene Xa21 in transgenic rice. Theor. Appl. Genet 2004, 109, 534–542. [Google Scholar]
- Sivamani, E.; Huet, H.; Shen, P.; Ong, C.A.; de Kochko, A.; Fauquet, C.; Beachy, R.N. Rice plant (Oryza sativa L.) containing rice tungro spherical virus (RTSV) coat protein transgenes are resistant to virus infection. Mol. Breed 1999, 5, 177–185. [Google Scholar]
- Tyagi, A.K.; Mohanty, A. Rice transformation for crop improvement and functional genomics. Plant Sci 2000, 158, 1–18. [Google Scholar]
- Bajaj, S.; Mohanty, A. Recent advances in rice biotechnology—towards genetically superior transgenic rice. Plant Biotechnol. J 2005, 3, 275–307. [Google Scholar]
- Ye, X.; Al-Babili, S.; Kloti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Engineering the provitamin A (beta-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science 2000, 287, 303–305. [Google Scholar]
- Wu, G.; Cui, H.; Ye, G.; Xia, Y.; Sardana, R.; Cheng, X.; Li, Y.; Altosaar, I.; Shu, Q. Inheritance and expression of the cry1Ab gene in Bt (Bacillus thuringiensis) transgenic rice. Theor. Appl. Genet 2002, 104, 727–734. [Google Scholar]
- Tu, J.; Datta, K.; Alam, M.F.; Fan, Y.; Khush, G.S.; Datta, S.K. Expression and function of a hybrid Bt toxin gene in transgenic rice conferring resistance to insect pests. Plant Biotechnol 1998, 15, 195–203. [Google Scholar]
- Tu, J.; Zhang, G.; Datta, K.; Xu, C.; He, Y.; Zhang, Q.; Khush, G.S.; Datta, S.K. Field performance of transgenic elite commercial hybrid rice expressing Bacillus thuringiensis delta-endotoxin. Nat. Biotechnol 2000, 18, 1101–1104. [Google Scholar]
- Tu, J.; Datta, K.; Oliva, N.; Zhang, G.; Xu, C.; Khush, G.S.; Zhang, Q.; Datta, S.K. Site-independently integrated transgenes in the elite restorer rice line Minghui 63 allow removal of a selectable marker from the gene of interest by self-segregation. Plant Biotechnol. J 2003, 1, 155–165. [Google Scholar]
- Li, D.; Fu, Q.; Wang, F.; Yao, Q.; Lai, F.; Wu, J.C.; Zhang, Z.T. Resistance of transgenic rice containing both sck and cry1Ac genes against Chilo suppressalis and Cnaphalocrocis medinalis. Chin. J. Rice Sci 2004, 18, 43–47. [Google Scholar]
- Zhang, Y.J.; Zhao, H.Y.; Wu, K.M.; Zhao, K.J.; Peng, Y.F.; Guo, Y.Y. Expression of Cry1Ac/CpTI transgenic rice and its resistance in different stages to Chilo suppressalis. J. Agric. Biotechnol 2004, 12, 76–79. [Google Scholar]
- James, C. Global Status of Commercialized Transgenic Crops; ISAAA Briefs No. 41; International Service for the Acquisition of Agri-biotech Applications (ISAAA): Ithaca, NY, USA, 2009. [Google Scholar]
- Paine, J.A.; Shipton, C.A.; Chaggar, S.; Howells, R.M.; Kennedy, M.J.; Vernon, G.; Wright, S.Y.; Hinchliffe, E.; Adams, J.L.; Silverstone, A.L.; et al. Improving the nutritional value of Golden Rice through increased pro-vitamin A content. Nat. Biotechnol 2005, 23, 482–487. [Google Scholar]
- Potrykus, P.K.; Burkhardt, S.K.; Datta, J.; Fütterer, G.C.; Ghosh-Biswas, A.; Klöti, A.; Spangenberg, G.; Wünn, J. Genetic engineering of indica rice in support of sustained production of affordable and high quality food in developing countries. Euphytica 1995, 85, 441–449. [Google Scholar]
- Yang, L.; Pan, A.; Zhang, K.; Guo, J.; Yin, C.; Chen, J.; Huang, C.; Zhang, D. Identification and quantification of three genetically modified insect resistant cotton lines using conventional and TaqMan real-time polymerase chain reaction methods. J. Agric. Food Chem 2005, 53, 6222–6229. [Google Scholar]
- Babekova, R.; Funk, T.; Pecoraro, S.; Engel, K.H.; Busch, U. Development of an event-specific realtime PCR detection method for the transgenic Bt rice line KMD1. Eur. Food Res. Technol 2009, 228, 707–716. [Google Scholar]
- Wu, G.; Wu, Y.H.; Nie, S.J.; Zhang, L.; Xiao, L.; Cao, Y.L.; Lu, C.M. Real-time PCR method for detection of the transgenic rice event TT51–1. Food Chem 2010, 119, 417–422. [Google Scholar]
- Su, C.; Xie, J.; Wang, X.; Peng, Y. Integrated structure and event-specific real-time detection of transgenic cry1Ac/SCK rice Kefeng 6. Eur. Food Res. Technol 2011, 232, 351–359. [Google Scholar]
- De Franchis, R.; Cross, N.C.; Foulkes, N.S.; Cox, T.M. A potent inhibitor of Taq polymerase copurifies with human genomic DNA. Nucleic Acids Res 1988, 16, 10355. [Google Scholar]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res 2000, 28, e63. [Google Scholar]
- Ohtsuka, K.; Yanagawa, K.; Takatori, K.; Kudo, Y.H. Detecton of Salmonella entericain naturally contaminated liquid eggs by loop-mediated isothermal amplification, and characterization of Salmonella isolates. Appl. Environ. Microb 2005, 71, 6730–6735. [Google Scholar]
- Misawa, Y.; Yoshida, A.; Saito, R.; Yoshida, H.; Okuzumi, K.; Ito, N.; Okada, M.; Moriya, K.; Koike, K. Application of loop-mediated isothermal amplification technique to rapid and direct detection of methicillin-resistant Staphylococcus aureus (MRSA) in blood cultures. J. Infect. Chemother 2007, 13, 134–140. [Google Scholar]
- Imai, M.; Ninomiya, A.; Minekawa, H.; Notomi, T.; Ishizaki, T.; van Tu, P.; Tien, N.T.K.; Tashiro, M.; Odagiri, T. Rapid diagnosis of H5N1 avian influenza virus infection by newly developed influenza H5 hemagglutinin gene-specific loop-mediated isothermal amplification method. J. Virol. Methods 2007, 141, 173–180. [Google Scholar]
- Wang, Y.; Yuan, X.; Li, Y.; Yu, K.; Yang, J.; Xu, H.; Zhang, Y.; Liao, M.; Qin, Z. Rapid detection of newly isolated Tembusu-related Flavivirus by reverse-transcription loop-mediated isothermal amplification assay. Virol. J. 2011, 8. [Google Scholar] [CrossRef]
- Chen, R.; Tong, Q.; Zhang, Y.; Lou, D.; Kong, Q.; Lu, S.; Zhuo, M.; Wen, L.; Lu, S. Loop-mediated isothermal amplification: Rapid detection of Angiostrongylus cantonensis infection in Pomacea canaliculata. Parasit Vectors 2011, 4. [Google Scholar] [CrossRef]
- Kong, Q.M.; Lu, S.H.; Tong, Q.B.; Lou, D.; Chen, R.; Zheng, B.; Kumagai, T.; Wen, L.Y.; Ohta, N.; Zhou, X.N. Loop-mediated isothermal amplification (LAMP): Early detection of Toxoplasma gondii infection in mice. Parasit Vectors 2012, 5. [Google Scholar] [CrossRef]
- Lee, D.; La Mura, M.; Allnutt, T.R.; Powell, W. Detection of genetically modified organisms (GMOs) using isothermal amplification of target DNA sequences. BMC Biotechnol. 2009, 9. [Google Scholar] [CrossRef]
- Guan, X.Y.; Guo, J.C.; Shen, P.; Yang, L.T.; Zhang, D.B. Visual and rapid detection of two genetically modified soybean events using Loop-mediated Isothermal Amplification method. Food Anal. Methods 2010, 3, 313–320. [Google Scholar]
- Liu, M.; Luo, Y.; Tao, R.; He, R.; Jiang, K.; Wang, B.; Wang, L. Sensitive and rapid detection of genetic modified soybean (Roundup Ready) by loop-mediated isothermal amplification. Biosci. Biotechnol. Biochem 2009, 73, 2365–2369. [Google Scholar]
- Kiddle, G.; Hardinge, P.; Buttigieg, N.; Gandelman, O.; Pereira, C.; McElgunn, C.J.; Rizzoli, M.; Jackson, R.; Appleton, N.; Moore, C.; et al. GMO detection using a bioluminescent real time reporter (BART) of loop mediated isothermal amplification (LAMP) suitable for field use. BMC Biotechnol. 2012, 12. [Google Scholar] [CrossRef]
- European Network of GMO Laboratories (ENGL). Event-specific method for the quantification of rice line LLRICE62 using real-time PCR protocol. 2008. Available online: http://gmo-crl.jrc.ec.europa.eu/summaries/LLRICE62_val_report.pdf accessed on 5 May 2012.
- Goto, M.; Honda, E.; Ogura, A.; Nomoto, A.; Hanaki, K. Colorimetric detection of loop-mediated isothermal amplification reaction by using hydroxy naphthol blue. Biotechniques 2009, 46, 167–172. [Google Scholar]
- Arumuganathan, K.; Earle, E. Nuclear DNA content of some important plant species. Plant Mol. Biol. Rep 1991, 9, 208–218. [Google Scholar]
- European Network of GMO Laboratories (ENGL). Definition of minimum performance requirements for analytical methods of GMO testing. 2008. Available online: http://gmo-crl.jrc.ec.europa.eu/doc/Min_Perf_Requirements_Analytical_methods.pdf accessed on 5 May 2012.
- Primer Explorer V4 Software. Operation procedure for designing regular/loop primers. Available online: http://primerexplorer.jp/elamp4.0.0/index.html accessed on 10 January 2012.
- Rigano, L.A.; Marano, M.R.; Castagnaro, A.P.; do Amaral, A.M.; Vojnov, A.A. Rapid and sensitive detection of Citrus Bacterial Canker by loop-mediated isothermal amplification combined with simple visual evaluation methods. BMC Microbiol 2010, 10, 176. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers name | Sequence (5′-3′) | Target | Amplicon size (bp) | Reference |
|---|---|---|---|---|
| PLD-F3 | TGAACAAGATTGAAGCTGGTG | PLD gene | 183 | This work |
| PLD-B3 | CCCTCGGGTCAGCATTGA | |||
| PLD-FIP | TCCACTAGCAGGAGGTCCTTTTTAGCGTT TTGCAGTCTATGTTG | |||
| PLD-BIP | GTGCAGGCAATACTGGATTGGTTTTCCT CTTCGCCTCAAGTGC | |||
| KMD1-F3 | CGGCGATGGCGATGC | Junction of KMD1 | 188 | This work |
| KMD1-B3 | CTGTTGCCCGTCTCACTGGT | |||
| KMD1-FIP | TATCCCGAGATGGGCAGGCATTTTTCCA ACTGCGGCGGGT | |||
| KMD1-BIP | TTGTGGTGTAAACAAATTGACGCTTTTTT GAAAAGAAAAACCACCCCAGTAC | |||
| TT51-F3 | CCGGCGTCAATACGGGATA | Junction of TT51-1 | 202 | This work |
| TT51-B3 | TCGTAGCCCCACCACTAC | |||
| TT51-FIP | CGGTCATTGACTGGAGCGAGGTTTTATA CCGCGCCACATAGCA | |||
| TT51-BIP | AGAGACTGGTGATTTCAGCGGGTTTTCT TATCTGCCCCAGCACTC | |||
| KF6-F3 | ACCATGCTGCGATTCATG | Junction of KF6 | 187 | This work |
| KF6-B3 | ATCTTCATCCCTGGACTTG | |||
| KF6-FIP | GAGTGACACGAATTCAACCTGATTTTTG CACTAAATCAATACCTCCT | |||
| KF6-BIP | GGCTTGCAAATCCTGCATGTTTTTGTAAC AGAAATCAGCAACGT | |||
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, X.; Wang, X.; Jin, N.; Zhou, Y.; Huang, S.; Miao, Q.; Zhu, Q.; Xu, J. Endpoint Visual Detection of Three Genetically Modified Rice Events by Loop-Mediated Isothermal Amplification. Int. J. Mol. Sci. 2012, 13, 14421-14433. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms131114421
Chen X, Wang X, Jin N, Zhou Y, Huang S, Miao Q, Zhu Q, Xu J. Endpoint Visual Detection of Three Genetically Modified Rice Events by Loop-Mediated Isothermal Amplification. International Journal of Molecular Sciences. 2012; 13(11):14421-14433. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms131114421
Chicago/Turabian StyleChen, Xiaoyun, Xiaofu Wang, Nuo Jin, Yu Zhou, Sainan Huang, Qingmei Miao, Qing Zhu, and Junfeng Xu. 2012. "Endpoint Visual Detection of Three Genetically Modified Rice Events by Loop-Mediated Isothermal Amplification" International Journal of Molecular Sciences 13, no. 11: 14421-14433. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms131114421