Effects of Different Metals on Photosynthesis: Cadmium and Zinc Affect Chlorophyll Fluorescence in Durum Wheat


Abstract
:1. Introduction
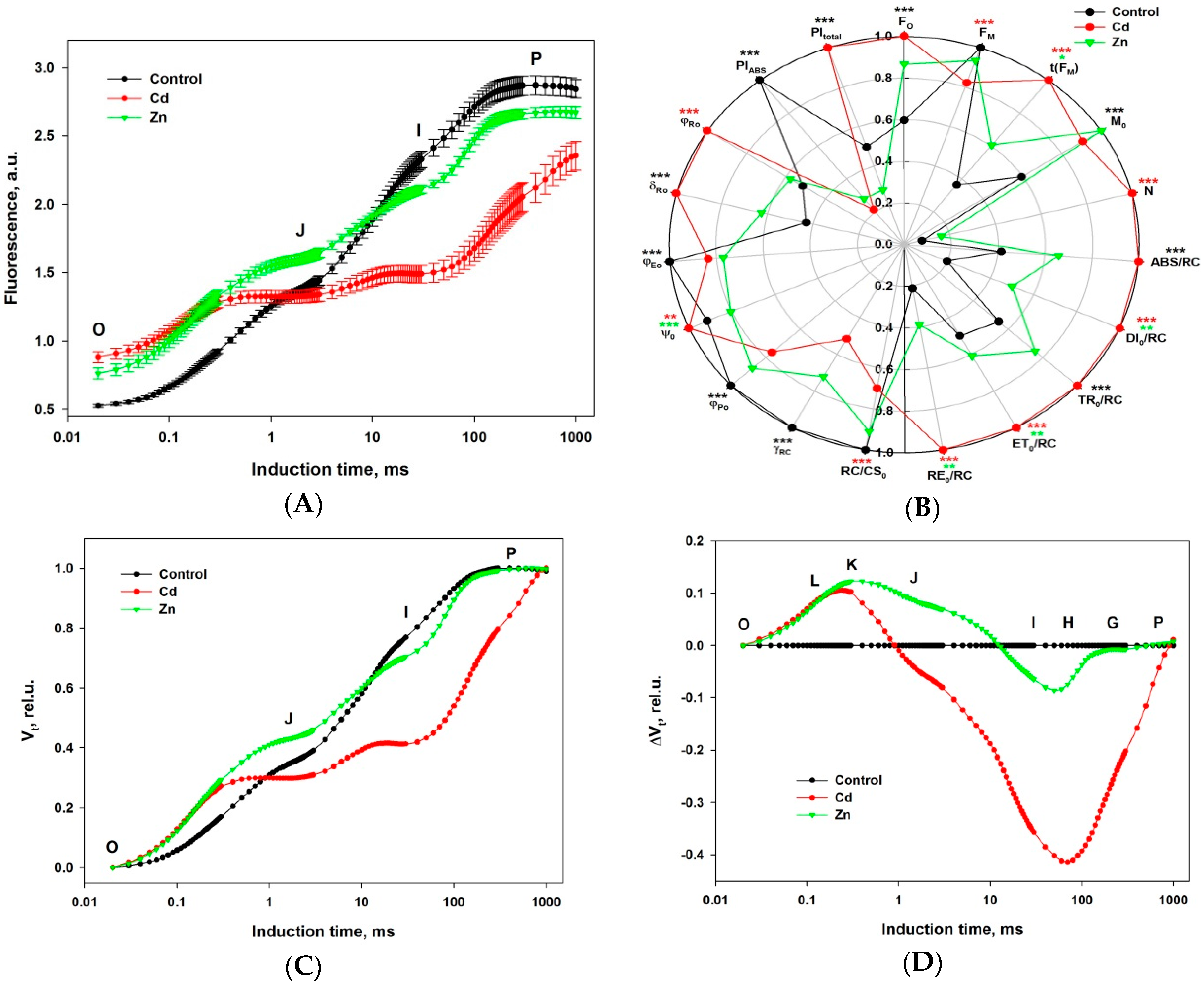
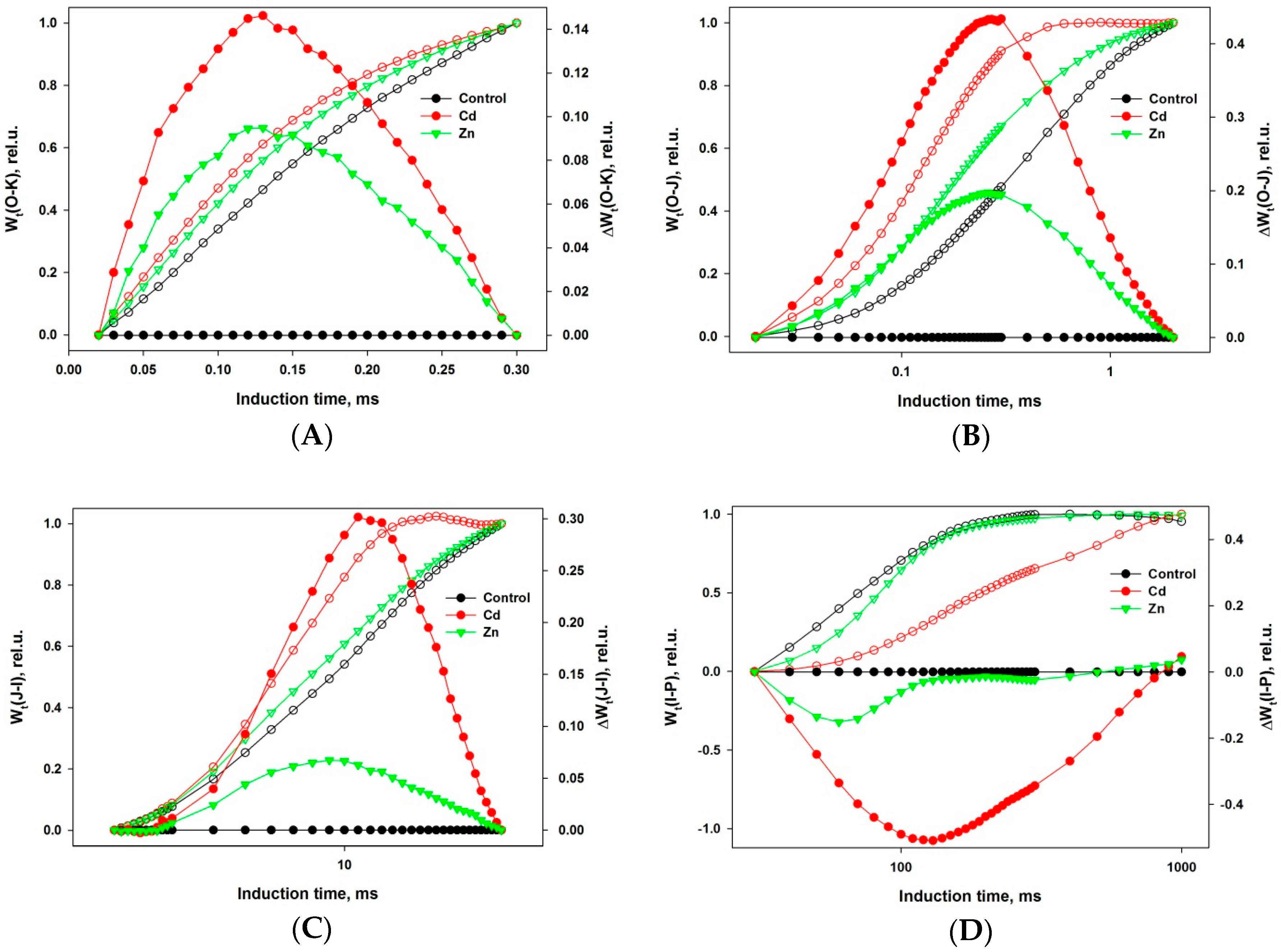
2. Results
- steeper initial rise for both metals,
- increase for Zn, decrease for Cd of the VJ (2 ms), and
- decrease (very strong for Cd) of the VI (30 ms).
- positive peak in FO-FJ,
- shoulder in FJ-FI and
- negative peak in FI-FP transient.
3. Discussion
4. Materials and Methods
Author Contributions
Conflicts of Interest
References
- Vassilev, A.; Schwitzguébel, J.-P.; Thewys, T.; van der Lelie, D.; Vangronsveld, J. The use of plants for remediation of metal-contaminated soils. Sci. World J. 2004, 4, 9–34. [Google Scholar] [CrossRef] [PubMed]
- Rout, G.R.; Das, P. Effect of Metal Toxicity on Plant Growth and Metabolism: I. Zinc. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Véronique, S., Alberola, C., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 873–884. [Google Scholar]
- Grant, C.A.; Buckley, W.T.; Bailey, L.D.; Selles, F. Cadmium accumulation in crops. Can. J. Plant Sci. 1998, 78, 1–17. [Google Scholar] [CrossRef]
- Koleva, L.; Staneva, D.; Yordanova, I.; Bineva, T.; Vassilev, A. Characterization of cadmum uptake by roots of durum wheat plants. JCEA 2008, 9, 533–538. [Google Scholar]
- Tang, L.; Aijun, Y.; Ming, Y.; Yetao, T.; Xi, L.; Rongliang, Q. Transcriptional up-regulation of genes involved in photosynthesis of the Zn/Cd hyperaccumulator Sedum alfredii in response to zinc and cadmium. Chemosphere 2016, 164, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Poschenrieder, Ch.; Tolra, R.; Barcelo, J. Can metals defend plants against biotic stress? Trends Plant Sci. 2006, 11, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Vassilev, A.; Perez-Sanz, A.; Cuypers, A.; Vangronsveld, J. Tolerance of two hydroponically grown Salix genotypes to excess Zn. J. Plant Nutr. 2007, 30, 1472–1482. [Google Scholar] [CrossRef]
- Vangronsveld, J.; Clijsters, H. Toxic effects of metals. In Plants and the Chemical Elements. Biochemistry, Uptake, Tolerance and Toxicity; Farago, M.E., Ed.; VCH Publishers: Weinheim, Germany, 1994; pp. 150–177. [Google Scholar]
- Vassilev, A.; Yordanov, I. Reductive analysis of factors limiting growth of cadmium-treated plants: A review. Bulg. J. Plant Physiol. 1997, 23, 114–133. [Google Scholar]
- Clijsters, H.; Van Assche, F. Inhibition of photosynthesis by heavy metals. Photosynth. Res. 1985, 7, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Van Assche, F.; Clijsters, H. Effects of metals on enzyme activity in plants. Plant, Cell and Environment 1990, 13, 195–206. [Google Scholar] [CrossRef]
- Barceló, J.; Poschenrieder, C. Plant water relations as affected by heavy metal stress: A review. J. Plant Nutr. 1990, 13, 1–37. [Google Scholar] [CrossRef]
- Chaney, R. Zinc phytotoxicity. In Zinc in Soils and Plants; Robson, A.D., Ed.; Kluwer Academic Publisher: Dordrecht, The Netherlands, 1993; pp. 135–150. [Google Scholar]
- Siedlecka, A. Some aspects of interactions between heavy metals and plant mineral nutrients. Acta Soc. Bot. Pol. 1995, 64, 265–272. [Google Scholar] [CrossRef]
- Cuypers, A.; Smeets, K.; Vangronsveld, J. Heavy Metal Stress in Plants. In Plant Stress Biology: From Genomics to Systems Biology; Hirt, H., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2009. [Google Scholar] [CrossRef]
- Krupa, Z. Cadmium against Higher Plant Photosynthesis—A Variety of Effects and Where Do They Possibly Come From? Z. Naturforsch. 1999, 54c, 723–729. [Google Scholar] [CrossRef]
- Cuypers, A.; Vangronsve, J.; Clijsters, H. The redox status of plant cells (AsA and GSH) is sensitive to zinc imposed oxidative stress in roots and primary leaves of Phaseolus vulgaris. Plant Physiol. Biochem. 2001, 39, 657–664. [Google Scholar] [CrossRef]
- Vassilev, A.; Nikolova, A.; Koleva, L.; Lidon, F. Effects of Excess Zn on Growth and Photosynthetic Performance of Young Bean Plants. J. Phytol. 2011, 3, 58–62. [Google Scholar]
- Van Assche, F.V.; Clijsters, H. Inhibition of photosynthesis by treatment of Phaseolus vulgaris with toxic concentration of zinc: Effects on electron transport and photophosphorylation. Physiol. Plant. 1986, 66, 717–721. [Google Scholar] [CrossRef]
- Vassilev, A.; Manolov, P. Chlorophyll fluorescence of barley (H. vulgare L.) seedlings grown in excess of Cd. Bulg. J. Plant Physiol. 1999, 25, 67–76. [Google Scholar]
- Kalaji, H.M.; Loboda, T. Photosystem II of barley seedlings under cadmium and lead stress. Soil Environ. 2007, 53, 511–516. [Google Scholar] [CrossRef]
- Sagardoy, R.; Morales, F.; Lόpez-Millán, A.F.; Abadía, A.; Abadía, J. Effects of zinc toxicity on sugar beet (Beta vulgaris L.) plants grown in hydroponic. Plant Biol. 2009, 11, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Vassilev, A.; Lidon, F.C.; Matos, M.D.; Ramalho, J.C.; Bareiro, M.G. Shoot cadmium accumulation and photosynthetic performance of barley at high Cd treatments. J. Plant Nutr. 2004, 27, 773–793. [Google Scholar] [CrossRef]
- Krupa, Z.; Öquist, G.; Huner, N.P.A. The effects of cadmium on photosynthesis of Phaseolus vulgaris—A fluorescence analysis. Physiol. Plant. 1993, 88, 626–630. [Google Scholar] [CrossRef] [PubMed]
- Vassilev, A.; Lidon, F.C.; Scotti, P.; da Graca, M.; Yordanov, I. Cadmium-induced changes in chloroplast lipids and photosystem activities of barley plants. Biol. Plant. 2004, 48, 153–156. [Google Scholar] [CrossRef]
- Cuypers, A.; Remans, T.; Weyens, N.; Colpaert, J.; Vassilev, A.; Vangronsveld, J. Soil plant relationships of metals and metalloids (Chapter 6). In Heavy Metals in Soils: Trace Metals and Metalloids in Soils and Their Bioavailability, 3rd ed.; Environmental Pollution Series; Alloway, B., Ed.; Springer Verlag: Berlin, Germany, 2012; pp. 161–195. ISBN 978-94-007-4469-1. [Google Scholar]
- Lazár, D. Parameters of photosynthetic energy partitioning. J. Plant Physiol. 2015, 175, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Srivastava, A.; Govindjee. Polyphasic chlorophyll a fluorescent transient in plants and cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Goltsev, V.N.; Kalaji, H.M.; Paunov, M.; Bąba, W.; Horaczek, T.; Mojski, J.; Kociel, H.; Allakhverdiev, S.I. Variable Chlorophyll Fluorescence and Its Use for Assessing Physiological Condition of Plant Photosynthetic Apparatus. Russ. J. Plant Physiol. 2017, 63, 869–893. [Google Scholar] [CrossRef]
- Kalaji, M.H.; Goltsev, V.N.; Żuk-Gołaszewska, K.; Zivcak, M.; Brestic, M. Chlorophyll Fluorescence: Understanding Crop Performance—Basics and Applications; CRC Press, T&F Group: Abingdon, UK, 2017; p. 222. ISBN 9781498764490. [Google Scholar]
- Kalaji, M.H.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 1–11. [Google Scholar] [CrossRef]
- Kalaji, M.H.; Schansker, G.; Brestic, M.; Bussotti, F.; Calatayud, A.; Ferroni, L.; Goltsev, V.; Guidi, L.; Jajoo, A.; Li, P.; et al. Frequently asked questions about chlorophyll fluorescence, the sequel. Photosynth. Res. 2017, 132, 13–66. [Google Scholar] [CrossRef] [PubMed]
- Stirbet, A.; Lazár, D.; Kromdijk, J. Chlorophyll a fluorescence induction: Can just a one-second measurement be used to quantify abiotic stress responses? Photosynthetica 2018, 1–19. [Google Scholar] [CrossRef]
- Luttge, U. Light-stress and crassulacean acid metabolism. Phyton 2000, 40, 65–82. [Google Scholar]
- Krause, G.H. Photoinhibition induced by low temperatures. In Photoinhibition of Photosynthesis. Environmental Plant Biology; Baker, N.R., Bowyer, J.R., Eds.; BIOS Scientific Publishers Ltd.: Oxford, UK, 1994; pp. 331–348. [Google Scholar]
- Koscielniak, J.; Biesaga-Koscielniak, J. Effects of exposure to short periods of suboptimal temperature during chili (5 degrees C) on gas exchange and chlorophyll fluorescence in maize seedlings (Zea mays L.). J. Agron. Crop. Sci. 1999, 183, 231–241. [Google Scholar] [CrossRef]
- Georgieva, K.; Yordanov, I. Temperature dependence of chlorophyll fluorescence parameters of pea seedlings. J. Plant Physiol. 1993, 142, 151–155. [Google Scholar] [CrossRef]
- Srivastava, A.; Strasser, R.J. Regulation of antenna structure and electron transport in Photosystem II of Pisum sativum under elevated temperature probed by the fast polyphasic chlorophyll a fluorescence transient: OKJIP. Biochim. Biophys. Acta 1997, 1320, 95–106. [Google Scholar] [CrossRef]
- Bukhov, N.; Carpentier, R. Heterogeneity of photosystem II reaction centers as influenced by heat treatment of barley leaves. Physiol. Plant. 2000, 110, 279–285. [Google Scholar] [CrossRef]
- Oukarroum, A.; Madidi, S.E.; Schansker, G.; Strasser, R.J. Probing the responses of barley cultivars (Hordeum vulgare L.) by chlorophyll a fluorescence OLKJIP under drought stress and re-watering. Environ. Exp. Bot. 2007, 60, 438–446. [Google Scholar] [CrossRef]
- Georgieva, K.; Maslenkova, L.; Peeva, V.; Markovska, Y.; Stefanov, D.; Tuba, Z. Comparative study on the changes in photosynthetic activity of the homoiochlorophyllous desiccation-tolerant Haberlea rhodopensis and desiccation-sensitive spinach leaves during desiccation and rehydration. Photosynth. Res. 2005, 85, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, K.; Szigeti, Z.; Sarvari, E.; Gaspar, L.; Maslenkova, L.; Peeva, V.; Peli, E.; Tuba, Z. Photosynthetic activity of homoiochlorophyllous desiccation tolerant plant Haberlea rhodopensis during dehydration and rehydration. Planta 2007, 225, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Goltsev, V.; Zaharieva, I.; Chernev, P.; Kouzmanova, M.; Kalaji, H.M.; Yordanov, I.; Krasteva, V.; Alexandrov, V.; Stefanov, D.; Allakhverdiev, S.I.; et al. Drought-induced modifications of photosynthetic electron transport in intact leaves: Analysis and use of neural networks as a tool for a rapid non-invasive estimation. Biochim. Biophys. Acta 2012, 1817, 1490–1498. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chen, L.-S.; Jiang, H.-X.; Tang, N.; Yang, L.-T.; Lin, Z.-H.; Li, Y.; Yang, G.-H. Effects of manganese-excess on CO2 assimilation, ribulose-1,5-bisphosphate carboxylase/oxygenase, carbohydrates and photosynthetic electron transport of leaves, and antioxidant systems of leaves and roots in Citrus grandis seedlings. BMC Plant Biol. 2010, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Zurek, G.; Rybka, K.; Pogrzeba, M.; Krzyżak, J.; Prokopiuk, K. Chlorophyll a Fluorescence in Evaluation of the Effect of Heavy Metal Soil Contamination on Perennial Grasses. PLoS ONE 2014, 9, e91475. [Google Scholar] [CrossRef] [PubMed]
- Cuchiara, C.C.; Silva, I.M.C.; Martinazzo, E.G.; Braga, E.J.B.; Bacarin, M.A.; Peters, J.A. Chlorophyll Fluorescence Transient Analysis in Alternanthera tenella Colla Plants Grown in Nutrient Solution with Different Concentrationsof Copper. J. Agric. Sci. 2013, 5, 8–16. [Google Scholar]
- Franić, M.; Galić, V.; Mazur, M.; Šimić, D. Effects of excess cadmium in soil on JIP-test parameters, hydrogen peroxide content and antioxidant activity in two maize inbreds and their hybrid. Photosynthetica 2017, 55, 1–10. [Google Scholar] [CrossRef]
- Tsimilli-Michael, M.; Strasser, R.J. In vivo assessment of stress impact on plant’s vitality: Applications in detecting and evaluating the beneficial role of Mycorrhization on hostplants. In Mycorrhiza: State of the Art, Genetics and Molecular Biology, Eco-Function, Biotechnology, Eco-Physiology, Structure and Systematics, 3rd ed.; Varma, A., Ed.; Springer Verlag: Berlin/Heidelberg, Germany, 2008; pp. 679–703. [Google Scholar]
- Lazár, D. The polyphasic chlorophyll a fluorescence rise measured under high intensity of exciting light. Funct. Plant Biol. 2006, 33, 9–30. [Google Scholar] [CrossRef]
- Havaux, M. Characterization of thermal damage to the photosynthetic electron transport system in potato leaves. Plant Sci. 1993, 94, 19–33. [Google Scholar] [CrossRef]
- Yamane, Y.; Kashino, Y.; Koike, H.; Satoh, K. Effects of high temperatures on photosynthetic systems in higher plants. In Photosynthesis: From Light to Biosphere; Mathis, P., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; Volume 5, pp. 849–852. [Google Scholar]
- Strasser, B. Donor side capacity of Photosystem II probed by chlorophyll a fluorescence transients. Photosynth. Res. 1997, 52, 147–155. [Google Scholar] [CrossRef]
- Steffen, R.; Eckert, H.-J.; Kelly, A.A.; Dörmann, P.; Renger, G. Investigations on the reaction pattern of photosystem II in leaves from Arabidopsis thaliana by time-resolved fluorometric analysis. Biochemistry-US 2005, 44, 3123–3133. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Tsimilli-Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim. Biophys. Acta 2010, 1797, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis. Advances in Photosynthesis and Respiration; Papageorgiou, G., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 321–362. [Google Scholar]
- Joliot, A.; Joliot, P. Etude cinetique de la reaction photochimique liberant loxygene au cours de la photosynthese. Comptes Rendus Acad. Sci. 1964, 258, 4622. [Google Scholar]
- Tsimilli-Michael, M.; Strasser, R. The energy flux theory 35 years later: Formulations and applications. Photosynth. Res. 2013, 117, 289–320. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Stirbet, A. Heterogeneity of photosystem II probed by the numerically simulated chlorophyll a fluorescence rise (O–J–I–P). Math. Comput. Simul. 1998, 48, 3–9. [Google Scholar] [CrossRef]
- Lazár, D.; Pospíšil, P.; Nauš, J. Decrease of fluorescence intensity after the K step in chlorophyll a fluorescence induction is suppressed by electron acceptors and donors to photosystem 2. Photosynthetica 1999, 37, 255–265. [Google Scholar] [CrossRef]
- Strasser, R.J.; Greppin, H. Primary reactions of photochemistry in higher plants. In Photosynthesis: Structure and Molecular Organisation of the Photosynthetic Apparatus; Akoyunoglou, G., Ed.; Balaban International Science Services: Philiadelphia, PA, USA, 1981; Volume III, pp. 717–726. [Google Scholar]
- Strasser, R.J. The grouping model of plant photosynthesis: Heterogeneity of photosynthetic units in thylakoids. In Photosynthesis: Structure and Molecular Organisation of the Photosynthetic Apparatus; Akoyunoglou, G., Ed.; Balaban International Science Services: Philiadelphia, PA, USA, 1981; Volume III, pp. 727–737. [Google Scholar]
- Guisse, B.; Srivastava, A.; Strasser, R.J. Effects of high temperature and water stress on the polyphasic chlorophyll a fluorescence transient of potato leaves. In Photosynthesis: From Light to Biosphere; Mathis, P., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 913–916. [Google Scholar]
- De Ronde, J.A.; Cress, W.A.; Krüger, G.H.J.; Strasser, R.J.; Van Staden, J. Photosynthetic response of transgenic soybean plants, containing an Arabidopsis P5CR gene, during heat and drought stress. J. Plant Physiol. 2004, 161, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Beadle, C.L. Growth analysis. In Photosynthesis and Production in a Changing Environment: A Field and Laboratory Manual; Scurlock, J.M.O., Bolhàr-Nordenkampf, H.R., Leegood, R.C., Long, S.P., Hall, D.O., Eds.; Springer Science & Business Media: Dordrecht, The Netherlands, 1993; pp. 36–46. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembrans. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanism, Regulation & Adaptation; Yunus, M., Pathre, U., Eds.; Taylor & Francis: London, UK, 2000; pp. 443–480. [Google Scholar]



{kind=link}
{kind=link}
| Treatments | Parameters | |||||
|---|---|---|---|---|---|---|
| DW (mg plant−1) Inhibition % | RGR (mg g DW−1 day−1) Inhibition % | A (µmol CO2 m−2 s−1) Inhibition % | ||||
| Control | 484 a | 0 | 140.6 a | 0 | 6.02 a | 0 |
| Cd 50 µM | 271 b | 55 | 68.8 b | 51 | 3.94 b | 35 |
| Zn 600 µM | 252 b | 58 | 64.1 b | 53 | 4.02 b | 33 |
| Treatments | Photosynthetic Pigments Content (mg g FW−1) | |||||
|---|---|---|---|---|---|---|
| Chlorophyll a | Inhibition % | Chlorophyll b | Inhibition % | Carotenoids | Inhibition % | |
| Control | 1.77 a | 0 | 0.70 a | 0 | 0.47 a | 0 |
| Cd 50 µM | 0.85 b | 52 | 0.48 b | 31 | 0.32 b | 32 |
| Zn 600 µM | 0.80 b | 55 | 0.53 b | 24 | 0.26 c | 45 |
| Chlorophyll Fluorescence Parameter | Description |
|---|---|
| Measured parameters and basic JIP-test parameters derived from the OJIP transient | |
| FO ~ F20µs | Minimum fluorescence, when all PSII reaction centers (RCs) are open; Fluorescence intensity at 20 µs |
| FJ | Fluorescence at the J-step (2 ms) of the O-J-I-P transient |
| FI | Fluorescence at the I-step (30 ms) of the O-J-I-P transient |
| FM = FP | Maximum recorded fluorescence at the P-step when all RCs are closed |
| t(FM) | Time (in ms) to reach maximal fluorescence FM |
| M0 = 4 × [(F300µs − F50µs)/(FM − F50µs)] | Approximated initial slope (in ms−1) of the fluorescent transient. This parameter is related to the rate of closure of reaction centers |
| N = Sm/Ss = Sm × M0 × (1/VJ) | Turnover number: number of QA reduction events between t = 0 and t(FM), where Sm is normalized total complementary area above the O-J-I-P transient (reflecting multiple-turnover QA reduction events), Ss—normalized total complementary area corresponding only to the O-J phase (reflecting single-turnover QA reduction events) and VJ—relative variable fluorescence at the J-step |
| Specific energy fluxes (per active, i.e., QA-reducing PSII RC) | |
| ABS/RC = M0 × (1/VJ) × (1/φPo) | Absorption flux per RC corresponding directly to its apparent antenna size—ratio between chlorophyll in antenna and chlorophyll in RC |
| DI0/RC = (ABS/RC) − (TR0/RC) | Dissipated energy flux per RC at the initial moment of the measurement, i.e., at t = 0 |
| TR0/RC = M0 × (1/VJ) | Trapping flux leading to QA reduction per RC at t = 0 |
| ET0/RC = M0 × (1/VJ) × (1 − VJ) | Electron transport flux from QA− to plastoquinone per RC at t = 0 |
| RE0/RC = M0 × (1/VJ) × (1 − VI) | Electron transport flux from QA− to the PSI end electron acceptors per RC at t = 0, where VI is the relative variable fluorescence at the I-step |
| Density of reaction centers | |
| RC/CS0 = φPo × (VJ/M0) × FO | Density of active PSII RCs. CS denotes cross section |
| Quantum yields and probabilities | |
| γRC = 1/[(ABS/RC) + 1] = RC/(ABS + RC) | Probability that PSII chlorophyll molecule functions as RC |
| φPo = TR0/ABS = [1 − (FO/FM)] | Maximum quantum yield of primary PSII photochemistry (at t = 0) |
| ψ0 = ET0/TR0 = 1 − VJ | Probability (at t = 0) that a trapped exciton moves an electron into the electron transport chain beyond QA− |
| φEo = ET0/ABS = φPo × ψEo | Quantum yield (at t = 0) for electron transport from QA− to plastoquinone |
| δRo = RE0/ET0 = (1 − VI)/(1 − VJ) | Efficiency/probability (at t = 0) with which an electron from the intersystem carriers moves to reduce end electron acceptors at the PSI acceptor side |
| φRo = RE0/ABS = φEo × δRo | Quantum yield (at t = 0) for reduction of end electron acceptors at the PSI acceptor side |
| Performance indexes | |
| PIABS = γRC/(1 − γRC) × φPo/(1 − φPo) × ψEo/(1 − ψEo) | Performance index of PSII based on absorption |
| PItotal = PIABS × δRo/(1 − δRo) | Performance index of electron flux to the final PSI electron acceptors, i.e., of both PSII and PSI |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paunov, M.; Koleva, L.; Vassilev, A.; Vangronsveld, J.; Goltsev, V. Effects of Different Metals on Photosynthesis: Cadmium and Zinc Affect Chlorophyll Fluorescence in Durum Wheat. Int. J. Mol. Sci. 2018, 19, 787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030787
Paunov M, Koleva L, Vassilev A, Vangronsveld J, Goltsev V. Effects of Different Metals on Photosynthesis: Cadmium and Zinc Affect Chlorophyll Fluorescence in Durum Wheat. International Journal of Molecular Sciences. 2018; 19(3):787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030787
Chicago/Turabian StylePaunov, Momchil, Lyubka Koleva, Andon Vassilev, Jaco Vangronsveld, and Vasilij Goltsev. 2018. "Effects of Different Metals on Photosynthesis: Cadmium and Zinc Affect Chlorophyll Fluorescence in Durum Wheat" International Journal of Molecular Sciences 19, no. 3: 787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030787