Metabolomic Signature in Sera of Multiple Sclerosis Patients during Pregnancy

 , , ,
, , ,  and
and
Abstract
:
1. Introduction
2. Results
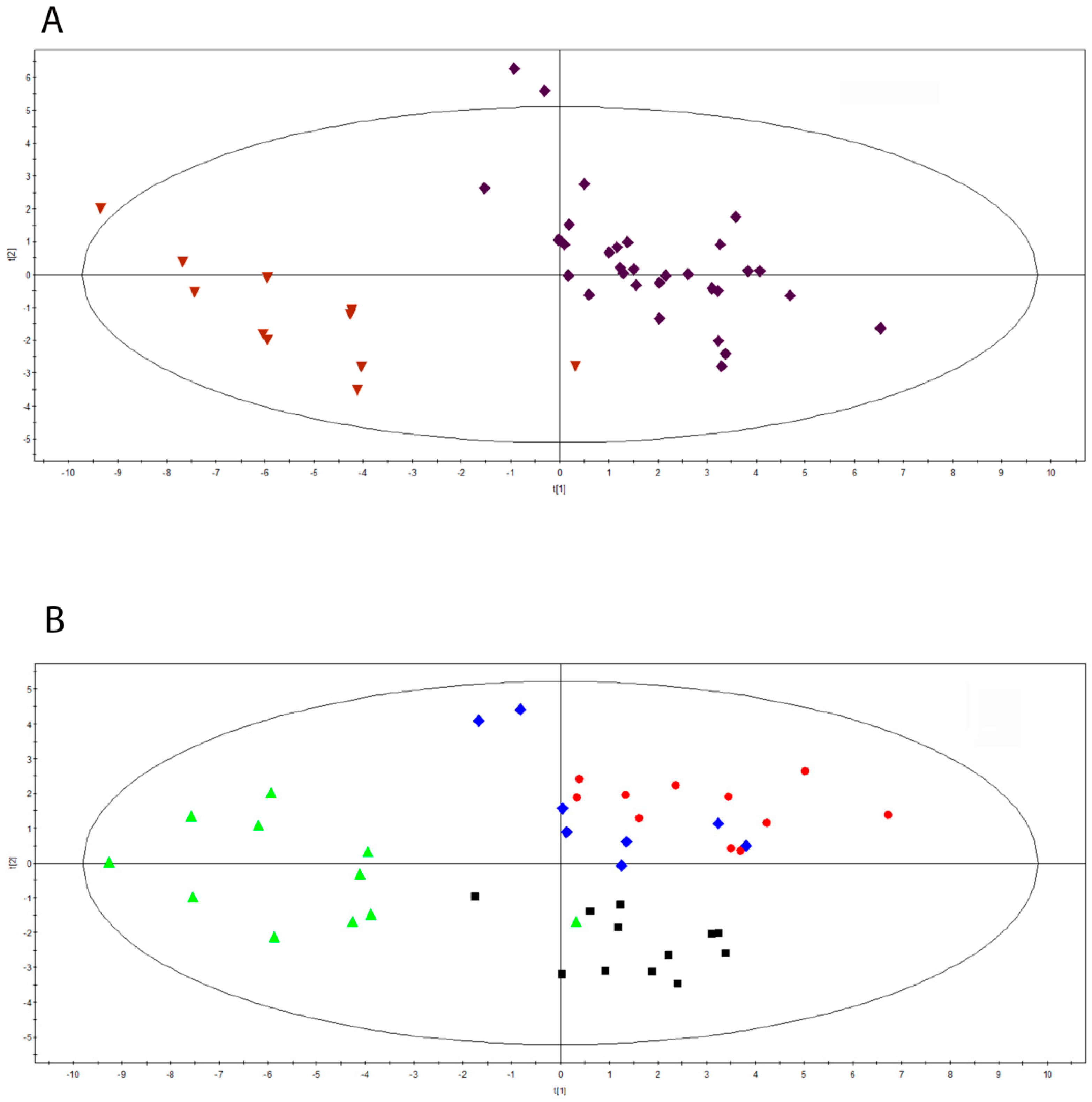
2.1. Serum Metabolomics Investigation during Pregnancy in Multiple Sclerosis
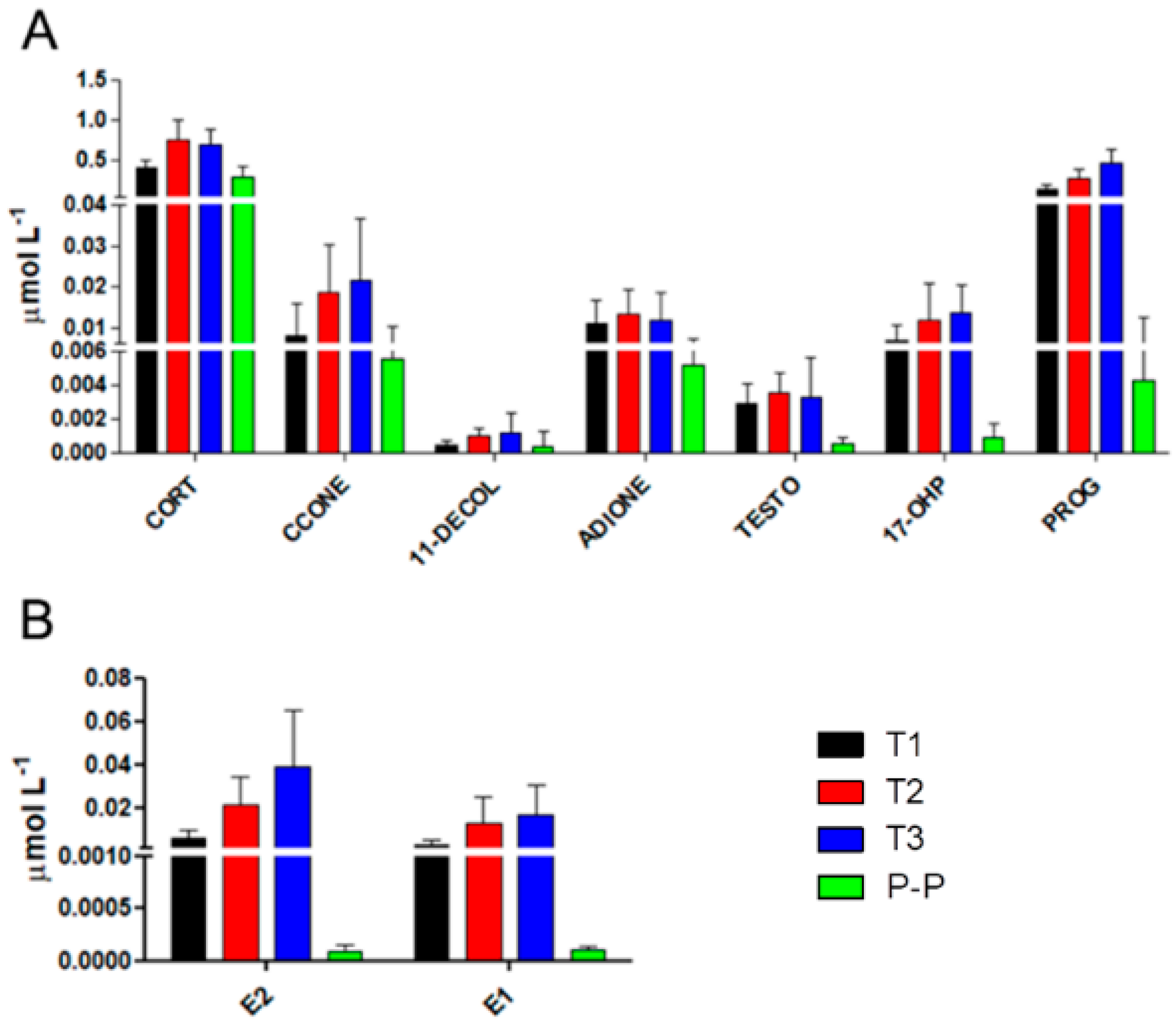
2.2. Characterization of Hormonal Fluctuations during Pregnancy in Multiple Sclerosis
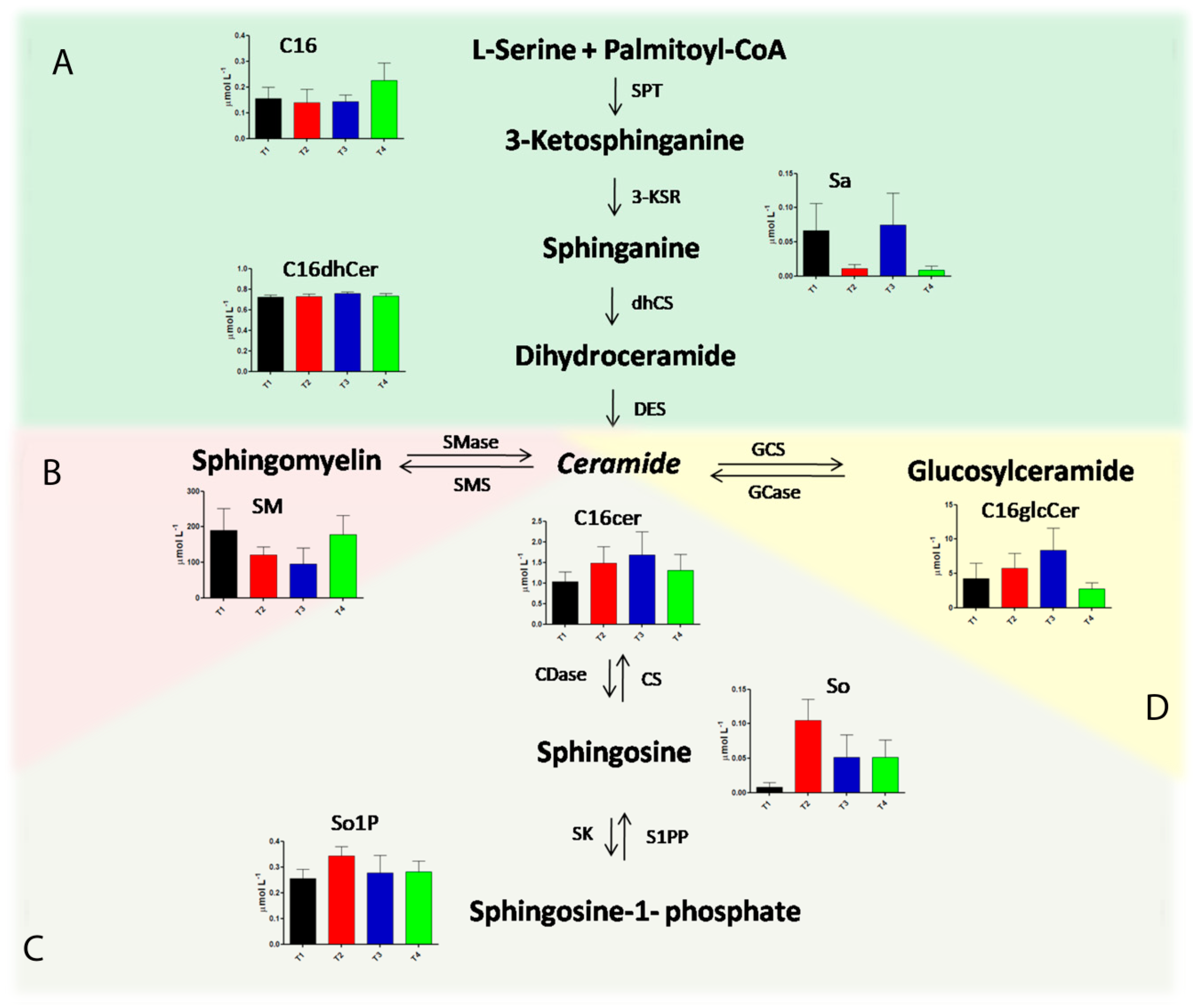
2.3. Alteration in Sphingolipids and Ceramides Metabolic Pathways during Pregnancy in Multiple Sclerosis
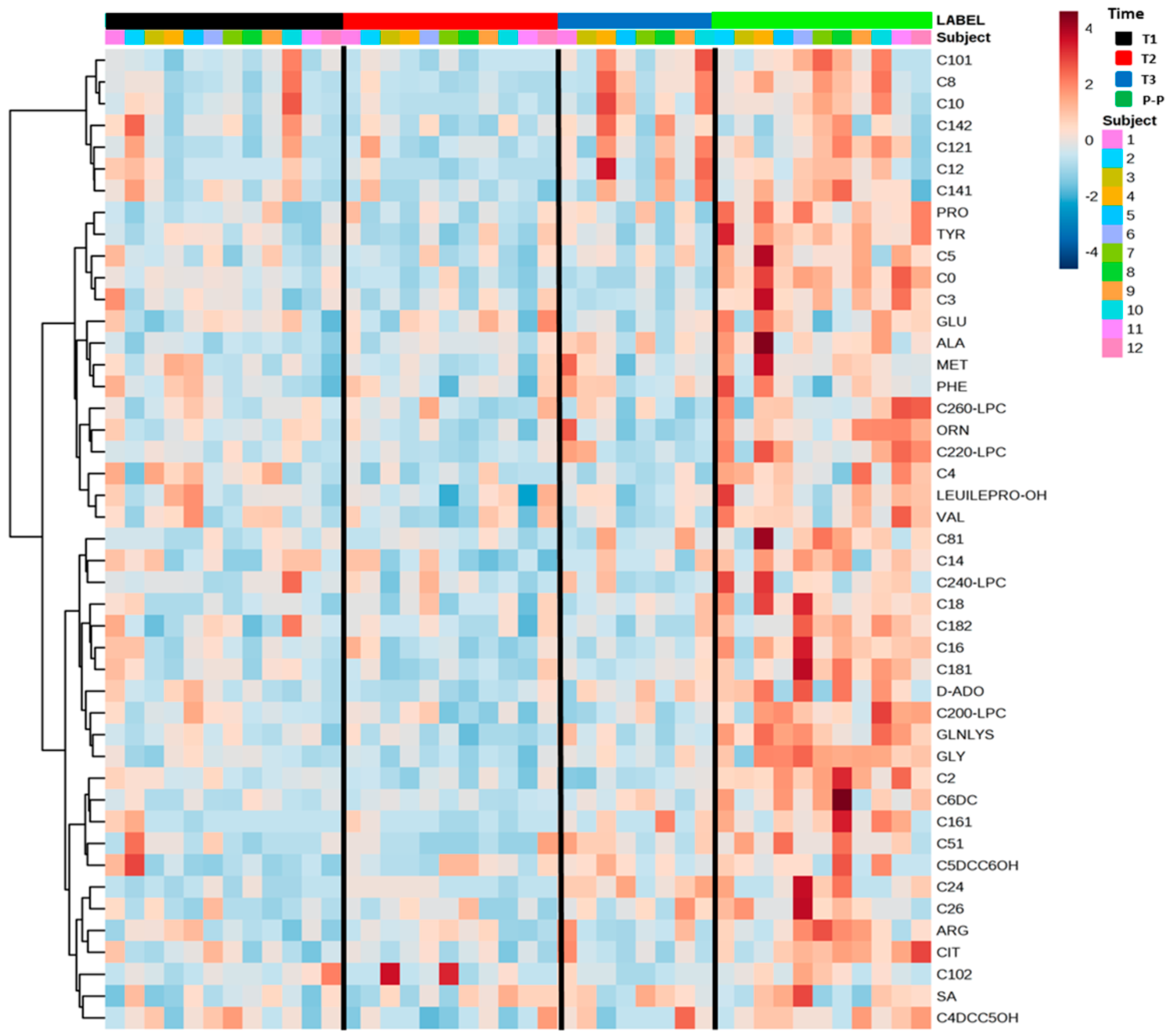
2.4. Serum Amino Acids and Acylcarnitines Profiles of Multiple Sclerosis in Pregnancy
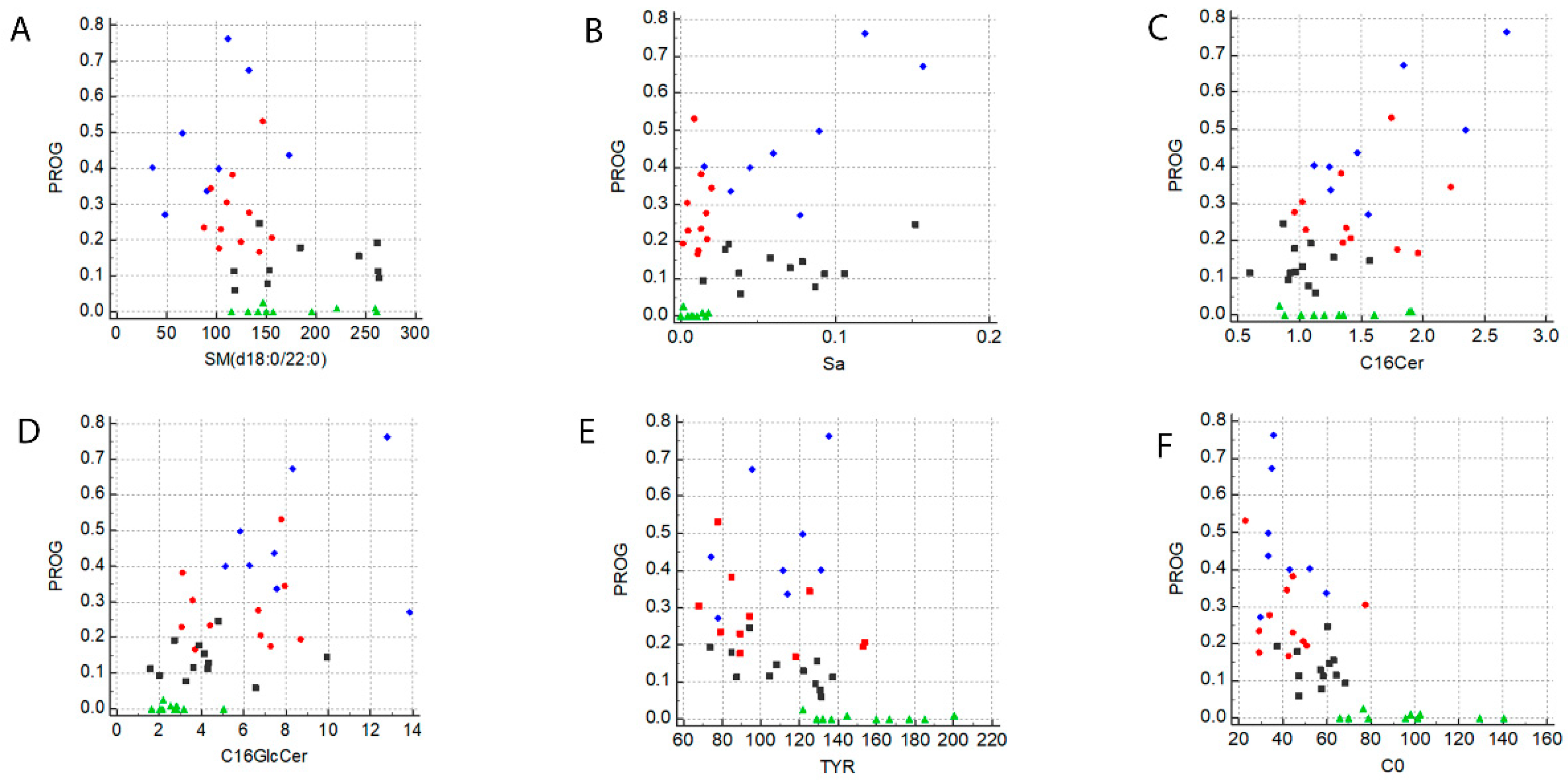
2.5. A Specific Metabolic Profile Correlates with Progesterone Levels
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Patients
4.3. Samples Collection
4.4. Materials
4.5. Standard Solutions, Calibrators, and Quality Controls
4.6. Sample Preparation and LC-MS/MS Analysis for Steroids Determination
4.7. Sample Preparation and LC-MS/MS Analysis for Estrogens Determination
4.8. Sample Preparation and LC-MS/MS Analysis for Sphingolipids and Ceramides Determination
4.9. Targeted Metabolomics for AAs and ACCs Determination
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lassmann, H.; Bruck, W.; Lucchinetti, C.F. The immunopathology of multiple sclerosis: An overview. Brain Pathol. 2007, 17, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Voskuhl, R.R.; Gold, S.M. Sex-related factors in multiple sclerosis susceptibility and progression. Nat. Rev. Neurol. 2012, 8, 255–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch-Henriksen, N.; Thygesen, L.C.; Stenager, E.; Laursen, B.; Magyari, M. Incidence of MS has increased markedly over six decades in Denmark particularly with late onset and in women. Neurology 2018, 90, e1954–e1963. [Google Scholar] [CrossRef] [PubMed]
- Voskuhl, R.; Momtazee, C. Pregnancy: Effect on Multiple Sclerosis, Treatment Considerations, and Breastfeeding. Neurotherapeutics 2017, 14, 974–984. [Google Scholar] [CrossRef] [PubMed]
- Confavreux, C.; Hutchinson, M.; Hours, M.M.; Cortinovis-Tourniaire, P.; Moreau, T. Rate of pregnancy-related relapse in multiple sclerosis. Pregnancy in Multiple Sclerosis Group. N. Engl. J. Med. 1998, 339, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Schwendimann, R.N.; Alekseeva, N. Gender issues in multiple sclerosis. Int. Rev. Neurobiol. 2007, 79, 377–392. [Google Scholar] [PubMed]
- Brann, D.W.; Dhandapani, K.; Wakade, C.; Mahesh, V.B.; Khan, M.M. Neurotrophic and neuroprotective actions of estrogen: Basic mechanisms and clinical implications. Steroids 2007, 72, 381–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spence, R.D.; Voskuhl, R.R. Neuroprotective effects of estrogens and androgens in CNS inflammation and neurodegeneration. Front. Neuroendocrinol. 2012, 33, 105–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, S.M.; Voskuhl, R.R. Estrogen and testosterone therapies in multiple sclerosis. Prog. Res. 2009, 175, 239–251. [Google Scholar] [Green Version]
- McGaha, T.L.; Huang, L.; Lemos, H.; Metz, R.; Mautino, M.; Prendergast, G.C.; Mellor, A.L. Amino acid catabolism: A pivotal regulator of innate and adaptive immunity. Immunol. Rev. 2012, 249, 135–157. [Google Scholar] [CrossRef] [PubMed]
- Schoderbeck, M.; Auer, B.; Legenstein, E.; Genger, H.; Sevelda, P.; Salzer, H.; Marz, R.; Lohninger, A. Pregnancy-related changes of carnitine and acylcarnitine concentrations of plasma and erythrocytes. J. Perinat. Med. 1995, 23, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Metrustry, S.J.; Karhunen, V.; Edwards, M.H.; Menni, C.; Geisendorfer, T.; Huber, A.; Reichel, C.; Dennison, E.M.; Cooper, C.; Spector, T.; et al. Metabolomic signatures of low birthweight: Pathways to insulin resistance and oxidative stress. PLoS ONE 2018, 13, e0194316. [Google Scholar] [CrossRef] [PubMed]
- Rutkowsky, J.M.; Knotts, T.A.; Ono-Moore, K.D.; McCoin, C.S.; Huang, S.; Schneider, D.; Singh, S.; Adams, S.H.; Hwang, D.H. Acylcarnitines activate proinflammatory signaling pathways. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1378–E1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, S.; Fassbender, K. Spingolipids in Multiple Sclerosis. Cell. Physiol. Biochem. 2010, 26, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Pieragostino, D.; Cicalini, I.; Lanuti, P.; Ercolino, E.; di Ioia, M.; Zucchelli, M.; Zappacosta, R.; Miscia, S.; Marchisio, M.; Sacchetta, P.; et al. Enhanced release of acid sphingomyelinase-enriched exosomes generates a lipidomics signature in CSF of Multiple Sclerosis patients. Sci. Rep. 2018, 8, 3071. [Google Scholar] [CrossRef] [PubMed]
- Brennan, P.J.; Tatituri, R.V.; Brigl, M.; Kim, E.Y.; Tuli, A.; Sanderson, J.P.; Gadola, S.D.; Hsu, F.F.; Besra, G.S.; Brenner, M.B. Invariant natural killer T cells recognize lipid self antigen induced by microbial danger signals. Nat. Immunol. 2011, 12, 1202–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalazar, G.; Preston, S.; Zigmond, E.; Ben Yaacov, A.; Ilan, Y. Glycolipids as immune modulatory tools. Mini Rev. Med. Chem. 2006, 6, 1249–1253. [Google Scholar] [CrossRef] [PubMed]
- Casella, G.; Colombo, F.; Finardi, A.; Descamps, H.; ILL-Raga, G.; Spinelli, A.; Podini, P.; Bastoni, M.; Martino, G.; Muzio, L.; et al. Extracellular vesicles containing IL-4 modulate neuroinflammation in a mouse model of multiple sclerosis. Mol. Ther. 2018, 26, 2107–2118. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.G., Jr.; Young, J.A.; Ray, S.K.; Wang, G.; Purohit, S.; Banik, N.L.; Dasgupta, S. Sphingosine toxicity in EAE and MS: Evidence for ceramide generation via serine-palmitoyltransferase activation. Neurochem. Res. 2017, 42, 2755–2768. [Google Scholar] [CrossRef] [PubMed]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Pieragostino, D.; Agnifili, L.; Cicalini, I.; Calienno, R.; Zucchelli, M.; Mastropasqua, L.; Sacchetta, P.; Del Boccio, P.; Rossi, C. Tear film steroid profiling in dry eye disease by liquid chromatography tandem mass spectrometry. Int. J. Mol. Sci. 2017, 18, 1349. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Calton, L.; Brown, H.A.; Gillingwater, S.; Wallace, A.M.; Petrucci, F.; Ciavardelli, D.; Urbani, A.; Sacchetta, P.; Morris, M. Confirmation of congenital adrenal hyperplasia by adrenal steroid profiling of filter paper dried blood samples using ultra-performance liquid chromatography-tandem mass spectrometry. Clin. Chem. Lab. Med. 2011, 49, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Calton, L.; Hammond, G.; Brown, H.A.; Wallace, A.M.; Sacchetta, P.; Morris, M. Serum steroid profiling for congenital adrenal hyperplasia using liquid chromatography-tandem mass spectrometry. Clin. Chim. Acta 2010, 411, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Zhao, Y.Y.; Dielschneider, R.F.; Gibson, S.B.; Curtis, J.M. An LC/MS/MS method for the simultaneous determination of individual sphingolipid species in B cells. J. Chromatogr. B 2016, 1031, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Del Boccio, P.; Perrotti, F.; Rossi, C.; Cicalini, I.; Di Santo, S.; Zucchelli, M.; Sacchetta, P.; Genovesi, D.; Pieragostino, D. Serum lipidomic study reveals potential early biomarkers for predicting response to chemoradiation therapy in advanced rectal cancer: A pilot study. Adv. Radiat. Oncol. 2017, 2, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Bonomini, M.; Di Liberato, L.; Del Rosso, G.; Stingone, A.; Marinangeli, G.; Consoli, A.; Bertoli, S.; De Vecchi, A.; Bosi, E.; Russo, R.; et al. Effect of an L-carnitine-containing peritoneal dialysate on insulin sensitivity in patients treated with CAPD: A 4-month, prospective, multicenter randomized trial. Am. J. Kidney Dis. 2013, 62, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Di Liberato, L.; Arduini, A.; Rossi, C.; Di Castelnuovo, A.; Posari, C.; Sacchetta, P.; Urbani, A.; Bonomini, M. L-Carnitine status in end-stage renal disease patients on automated peritoneal dialysis. J. Nephrol. 2014, 27, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Marzano, V.; Consalvo, A.; Zucchelli, M.; Levi Mortera, S.; Casagrande, V.; Mavilio, M.; Sacchetta, P.; Federici, M.; Menghini, R.; et al. Proteomic and metabolomic characterization of streptozotocin-induced diabetic nephropathy in TIMP3-deficient mice. Acta Diabetol. 2018, 55, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Stohr, R.; Cavalera, M.; Menini, S.; Mavilio, M.; Casagrande, V.; Rossi, C.; Urbani, A.; Cardellini, M.; Pugliese, G.; Menghini, R.; et al. Loss of TIMP3 exacerbates atherosclerosis in ApoE null mice. Atherosclerosis 2014, 235, 4438–4443. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | VIP Value | Kruskal–Wallis p-Value | T1 vs. T2 | T2 vs. T3 | T1 vs. T3 | T1 vs. P-P | T2 vs. P-P | T3 vs. P-P |
|---|---|---|---|---|---|---|---|---|
| So | 2.29 | <0.0001 | 0.0001 | ns | 0.05 | 0.05 | ns | ns |
| Sa1P | 1.80 | <0.0001 | 0.0001 | 0.01 | ns | 0.01 | ns | ns |
| C16Cer | 1.60 | <0.01 | 0.05 | ns | 0.05 | ns | ns | ns |
| C24dHCer | 1.57 | <0.001 | 0.01 | ns | 0.01 | 0.05 | ns | ns |
| C0 | 1.41 | <0.0001 | ns | ns | ns | 0.05 | 0.0001 | 0.0001 |
| So1P | 1.41 | <0.01 | 0.0001 | ns | ns | ns | 0.05 | ns |
| C16GlcCer | 1.32 | <0.0001 | ns | ns | 0.05 | ns | 0.01 | 0.0001 |
| C24 | 1.32 | <0.001 | ns | ns | 0.01 | 0.01 | ns | ns |
| C2 | 1.32 | <0.0001 | ns | ns | ns | 0.01 | 0.0001 | 0.0001 |
| GLY | 1.27 | <0.001 | ns | ns | ns | 0.01 | 0.0001 | ns |
| ALA | 1.25 | <0.01 | ns | ns | 0.05 | 0.01 | ns | ns |
| C18Cer | 1.23 | <0.01 | ns | ns | 0.01 | ns | ns | ns |
| GLNLYS | 1.21 | <0.001 | ns | ns | ns | 0.05 | 0.0001 | 0.05 |
| TYR | 1.09 | <0.001 | ns | ns | ns | 0.05 | 0.01 | 0.05 |
| PRO | 1.08 | <0.01 | ns | ns | ns | 0.01 | 0.05 | ns |
| Sa | 1.02 | <0.0001 | 0.01 | 0.01 | ns | 0.0001 | ns | 0.0001 |
| C18:1 | 1.00 | <0.01 | ns | ns | ns | ns | 0.01 | ns |
| Metabolite | ANOVA | T1 vs. T2 | T2 vs. T3 | T1 vs. T3 | T1 vs. P-P | T2 vs. P-P | T3 vs. P-P |
|---|---|---|---|---|---|---|---|
| CORT | <0.0001 | <0.01 | ns | <0.05 | ns | <0.0001 | <0.0001 |
| CCONE | <0.001 | ns | ns | <0.05 | ns | <0.05 | <0.01 |
| 11-DECOL | <0.01 | ns | ns | ns | ns | <0.01 | <0.05 |
| ADIONE | <0.005 | ns | ns | ns | <0.05 | <0.01 | ns |
| TESTO | <0.0001 | ns | ns | ns | <0.01 | <0.0001 | <0.01 |
| 17-OHP | <0.0001 | ns | ns | ns | <0.05 | <0.0001 | <0.0001 |
| PROG | <0.0001 | ns | ns | <0.01 | ns | <0.0001 | <0.0001 |
| E2 | <0.0001 | ns | ns | <0.01 | ns | <0.0001 | <0.0001 |
| E1 | <0.0001 | ns | ns | ns | ns | <0.0001 | <0.0001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, C.; Cicalini, I.; Zucchelli, M.; Di Ioia, M.; Onofrj, M.; Federici, L.; Del Boccio, P.; Pieragostino, D. Metabolomic Signature in Sera of Multiple Sclerosis Patients during Pregnancy. Int. J. Mol. Sci. 2018, 19, 3589. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113589
Rossi C, Cicalini I, Zucchelli M, Di Ioia M, Onofrj M, Federici L, Del Boccio P, Pieragostino D. Metabolomic Signature in Sera of Multiple Sclerosis Patients during Pregnancy. International Journal of Molecular Sciences. 2018; 19(11):3589. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113589
Chicago/Turabian StyleRossi, Claudia, Ilaria Cicalini, Mirco Zucchelli, Maria Di Ioia, Marco Onofrj, Luca Federici, Piero Del Boccio, and Damiana Pieragostino. 2018. "Metabolomic Signature in Sera of Multiple Sclerosis Patients during Pregnancy" International Journal of Molecular Sciences 19, no. 11: 3589. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113589