1. Introduction
The enteric nervous system (ENS) is responsible for the intrinsic innervation of the gastrointestinal (GI) tract. It is situated in the wall of the entire digestive tract, from the esophagus to the rectum. The enteric plexuses consist of millions of neurons, which play various roles and express a range of neurotransmitters and/or neuromodulators [
1]. Because of the high level of autonomy in the activity of the enteric neurons, the ENS is considered to be the “second” or “intestinal brain” [
2].
Depending on the animal species and the fragment of the GI tract, the ENS structure shows some differences. In the esophagus and stomach, the enteric neurons of large mammals (e.g., pigs) are grouped into two ganglionated plexuses: the myenteric plexus (MP)—located between longitudinal and circular muscle layers and the submucous plexus placed near the lamina propria of the mucosal layer. In the small and large intestines, the submucous plexus is additionally divided into the outer submucous plexus (OSP) and inner submucous plexus (ISP), which are placed in the submucosal layer: the OSP near the inner side of the circular muscle layer and the ISP near the lamina propria of the mucosa [
3,
4,
5].
It should be emphasized that enteric neurons are very diverse in terms of morphological, functional and electrophysiological properties, but the most important criterion of neuronal classification in the ENS is the neurochemical characterization of nerve cells [
3,
4]. To date, several dozen neuronal active substances have been found in the enteric neurons. They may play various functions and are involved in all aspects of gastrointestinal physiology, such as motility, excretive activity, intestinal blood flow and ion transport [
1,
6]. Previous studies have shown that the same neuronal cells may contain even a few active substances, which usually play similar roles [
7,
8] and studies on the co-localization of these substances contribute to a better understanding of their functions.
It is also known that enteric neurons may undergo changes concerning their structural, functional and chemical phenotype as a result of adaptive and/or neuroprotective responses to various pathological and physiological agents [
7,
9,
10,
11,
12,
13,
14], but knowledge of these issues is still fragmentary. In particular, little is known about the reactions of the enteric neurons to the maturation and aging of the organism [
15] and many aspects of this problem are still completely unknown. The second area of knowledge about the ENS which still remains obscure concerns sex-dependent differences in its organization. Despite some suggestions that, in males and females, the enteric neurons contain various active substances and respond differently to pathological factors [
16], exact studies on sex-dependent differences in the neurochemical characterization of neuronal cells in the ENS have not been conducted.
One neuronal substance which plays an important and multidirectional role in the ENS is calcitonin gene-related peptide (CGRP). Since the first identification of CGRP in 1982, the wide distribution of this peptide has been described in both the central and peripheral nervous systems [
17,
18]. In the GI tract of various mammal species, including humans, the presence of CGRP has been noted in all types of enteric plexuses from the esophagus to the rectum [
11,
19,
20,
21]. At first, CGRP is known as one of the most important factors participating in the conduction of sensory and pain stimuli [
12]. However, other studies have reported that CGRP is also involved in the suppression of gastric acid secretion, the relaxation of the GI muscular membrane, vasodilatation processes and regulation of the absorption of nutrients from the intestines [
22,
23,
24]. Moreover, it is also known that the expression of this neuropeptide in enteric nervous structures undergoes changes under the influence of some pathological stimuli [
7,
9], which suggests the participation of CGRP in adaptive and/or neuroprotective processes and the probable protective function of this substance against harmful factors [
25]. Nevertheless, many aspects of the CGRP functions in the ENS are still not fully understood. The previous reports on the impact of physiological conditions, such as the development and maturation of the digestive tract on the population of CGRP-positive nervous structures are rather scarce and apply only to nerve fibers [
10,
26,
27], and the influence of gender on these populations has not yet been studied.
Therefore, the aim of the present study was to investigate the impact of selected physiological states (such as age and gender) on the number, distribution and neurochemical characterization of CGRP-positive nervous structures in the wall of the porcine descending colon. Both the selection of domestic pig as an experimental animal and the descending colon as a fragment of the GI tract included in the study were not accidental. Due to the anatomical, neurochemical and electrophysiological similarities in the ENS organization between humans and pigs [
28], this species is considered to be an optimal animal model (significantly better than using rodents) for studies on the influence of various stimuli on the ENS in humans. In turn, the descending colon is where numerous pathological states occur, and many of them are connected with the aging of the organism [
26] and/or depend on gender [
29]. The present study not only contributes to the elucidation of the exact roles of CGRP in the maturation of the GI tract in individuals of the opposite sex, but it may also have practical applications. Namely, it may be the first step to a better understanding of the roles of the ENS during disorders connected with age and gender, which may contribute to improving the treatment of various intestinal diseases in the future.
3. Discussion
Previous studies have reported that the ENS may undergo changes under physiological stimuli, but the existing knowledge relating to these issues is rather scarce. It is known that nerve structures in the GI tract may be influenced by one’s diet, environmental factors or the composition of the gut microbiota [
30,
31,
32], but the main factors which may affect the ENS are maturation and aging of the organism [
26,
27]. Studies conducted on humans and rodents have shown that maturation and aging not only substantially affect the number and morphology of the enteric neurons, but may also change the synaptic profiles and expression of selected neuronal substances in the ENS, and the character of these changes depends on the mammal species, the segment of the GI tract and the type of active substance studied [
15,
33,
34].
The results obtained during the current study have shown age-dependent differences in the size of the population of CGRP-positive enteric neuronal structures and their neurochemical characterization in the porcine descending colon. Because the study was performed on adults and not older animals, these changes probably result from the development and maturation of the GI tract. On the other hand, their exact mechanisms are still unclear.
Given that the ENS undergoes major developmental and adaptive modifications during the maturation of the organism, including changes in neuronal morphology and electrophysiological properties, as well as the organization of the enteric synapses [
35], the changes in the population of CGRP-LI nervous structures, first of all, may be connected with adaptive activities of this peptide, which monitor the condition of the intestine and maintenance of gut homeostasis [
36,
37]. This is more likely since, on the one hand, previous studies have reported maturation-dependent changes in length, number and appearance of axons and dendrites of the enteric neurons [
35]. However, CGRP participates in physiological neuronal development through its involvement in the differentiation of neurons and the promotion of dendrite formation [
38,
39].
The next mechanism of observed differences may be connected with the participation of CGRP in the regulation of blood vessel functions. It is commonly known that the growth and maturation of the body require considerable vascular remodeling [
40]. In turn, CGRP is known as a factor that not only regulates vascular functions and blood flow in the intestine [
41], but also as a substance taking part in vasculogenesis during the development and maturation of the body through its influence on the expression of the vascular endothelial growth factor (VEGFA) [
42,
43].
The possibility that that changes in the number of CGRP-LI colonic nervous structures may result from other causes cannot be excluded. Because CGRP is involved in sensory stimuli conduction [
12], it may be connected with the maturation-dependent remodeling of the sensory neurons described in previous studies [
18]. In turn, the participation of CGRP in the control of the intestinal motility [
24] suggests that observed fluctuations result from developmental age-dependent changes in the muscles of the GI tract and in intestinal motility [
35,
44]. Moreover, it is known that clear changes in the enteric nervous structures may be caused by the influence of the intestinal microbiota [
31,
45], and the relatively well-known differences in the composition of gut flora in young and adult animals [
46] might have contributed to the observed changes. Another cause of the fluctuations in the number of CGRP-LI nervous structures (which among others is also connected with gut microbiota) may be connected with immunological processes [
45]. This is reinforced by the relatively well-known maturation-dependent remodeling of the immune system [
47], close correlations between the ENS and immune cells [
48], as well as the participation of CGRP in immunological reactions [
49].
It should be underlined that the exact direct mechanism of differences in the number of CGRP-LI enteric nervous structures noted in this study is also unknown. It may have resulted from fluctuations in various stages of protein synthesis. Nevertheless, based on the present results, which show a decrease in the number of CGRP-LI colonic nerve fibers along with a slight increase in the percentage of CGRP-LI neurons in females and without changes in such neurons in males, the most probable reasons for the noted fluctuations seem to be development-dependent changes in the intra-neuronal transport [
50,
51].
The observations made during the present study have also shown that the maturation of the body affects not only the number of CGRP-LI colonic nervous structures, but also changes (in a way, even more substantially) their neurochemical characterization. The obtained results, showing the presence of all active substances studied in CGRP-positive enteric neurons, have confirmed the considerable differentiation of such neuronal cells that were known from previous studies [
7]. Interestingly, neuronal factors noted in CGRP-LI neurons during the present investigation often play opposing roles, which could mean that the CGRP is engaged in various mechanisms within the GI tract. For example, acetylcholine is the main excitatory neuromediator within the ENS, while nitric oxide is considered to be one of the most important inhibitory factors within the GI tract [
52].
It is most likely that the participation of substances, which co-localize with CGRP in the enteric neurons in homeostasis maintenance in the colon during the growth and maturation of the gastrointestinal tract [
36,
37], have given rise to changes noted in the present study. The present study has shown that the degree of co-localization of CGRP with the majority of substances studied in the colonic enteric neurons is lower in adult animals. This is in agreement with the previous observations that the expression of acetylcholine, nitric oxide, GAL and SP in various parts of the nervous system was lower in older individuals [
15]. Moreover, the differences in the degree of co-localization of CGRP and other studied substances may result from the same causes and mechanisms as mentioned above for changes in CGRP-LI nervous structures.
The second aim of the present study was to investigate, for the first time, the sex-dependent differences in CGRP-LI enteric nervous structures in the descending colon. It should be underlined that the existing information about differences in the ENS organization (including number, phenotype and activity of the enteric nervous structures) between males and females is much more limited and fragmentary than knowledge about changes in the ENS with the maturation and aging of the body [
37]. Moreover, previous studies concerning this issue are inconclusive. Some of them have reported that there are no diversities between male and female ENS in certain species [
53], while others have shown such differences in reaction to some pathological factors and stress conditions [
16,
54].
Generally, during the present study, the percentage of CGRP-positive enteric neurons was similar in young animals of both sexes, while it was higher in the adult females than in males. It should be highlighted that the age of the adult animals was the age of sexual maturity, although the males were castrated at around one week of age, which was the cause of different levels of reproductive hormones in males and females before and after puberty. Thus, the differences observed during the present study in animals of opposite genders probably result from various levels of female hormones. It is all the more probable since it is known that female sex hormones may affect a wide range of functions of the GI tract via various types of estrogen receptors located among others on the enteric neurons in both males and females [
55,
56].
Primarily female sex hormones in the ENS stimulate enteric neurons to produce selected neuroactive substances and protect them against damage factors [
57,
58]. Moreover, these hormones may affect the intestinal motility, blood flow and functions of the mucosal layer [
16,
29,
37]. It is also known that female sex hormones regulate somatic and visceral sensitivity, especially in response to different pathological stimuli [
57], and some studies suggest that the estrogenic compounds regulating the release of CGRP (the major sensory factor) could, at least in part, provide a rational explanation for the gender differences in visceral pain sensitivity, which is higher in females [
58]. It should be underlined that CGRP is involved in the majority of above-mentioned gastro-intestinal processes affected by female sex hormones, including intestinal motility, protective mechanisms and mesenteric blood flow [
22,
23,
24], which may be at the heart of the observed changes.
The observations made in the present study of gender differences (similar to age-dependent changes) concerned not only the percentage of CGRP-LI nervous structures, but also their neurochemical characterization. These differences were more visible in adult animals and they are probably connected with the above-discussed influence of female sex hormones on the GI tract and the ENS. The changes in the degree of co-localization of CGRP with other substances may have resulted from the role of CGRP as a neuromodulator, influencing the secretion of other neuronal factors [
52]. However, they may also be connected with the direct influence of aging and/or gender on the synthesis and distribution of studied the neuromediators.
4. Materials and Methods
The experiment was performed on tissues collected from 20 pigs slaughtered at a commercial slaughterhouse. Fragments of the descending colon (from the same place in all animals, located about 20 cm before the anus) were taken immediately after the death of the pigs using a typical commercial method (carbon dioxide) and bowel removal. Depending on the age and sex of the animals, samples were divided into four groups (five animals in each). Descending colons were collected from young pigs before puberty (about 10 weeks old), the compounded group A (samples from females) and group B (males). Group C contained tissues from females after puberty (7–8 months old) and group D from mature males (also 7-8 months old). Both young and mature males were castrated at the age of about 1 week. All procedures during this study were performed according to Act for the Protection of Animals for Scientific or Educational Purposes of 15 January 2015 (Official Gazette 2015, No. 266), applicable in the Republic of Poland. Based on this act, due to the fact that tissues were taken from commercially slaughtered animals, approval from the Bioethical Committee for the present study was not required.
Immediately after collection, 2-cm (approx.) colon fragments were fixed in a solution of 4% buffered paraformaldehyde (pH 7.4). After one hour, the solution was changed into a phosphate buffer (0.1 M, pH 7.4). For the next three days, tissues were stored at 4 °C with a daily exchange of the buffer. Afterward, the colon fragments were put into 18% phosphate-buffered sucrose and stored for three weeks. The tissues were then frozen at −22 °C and cut perpendicular to the colonic lumen into 14-µm-thick sections using a freezing microtome (Microm, HM 525, Walldorf, Germany).
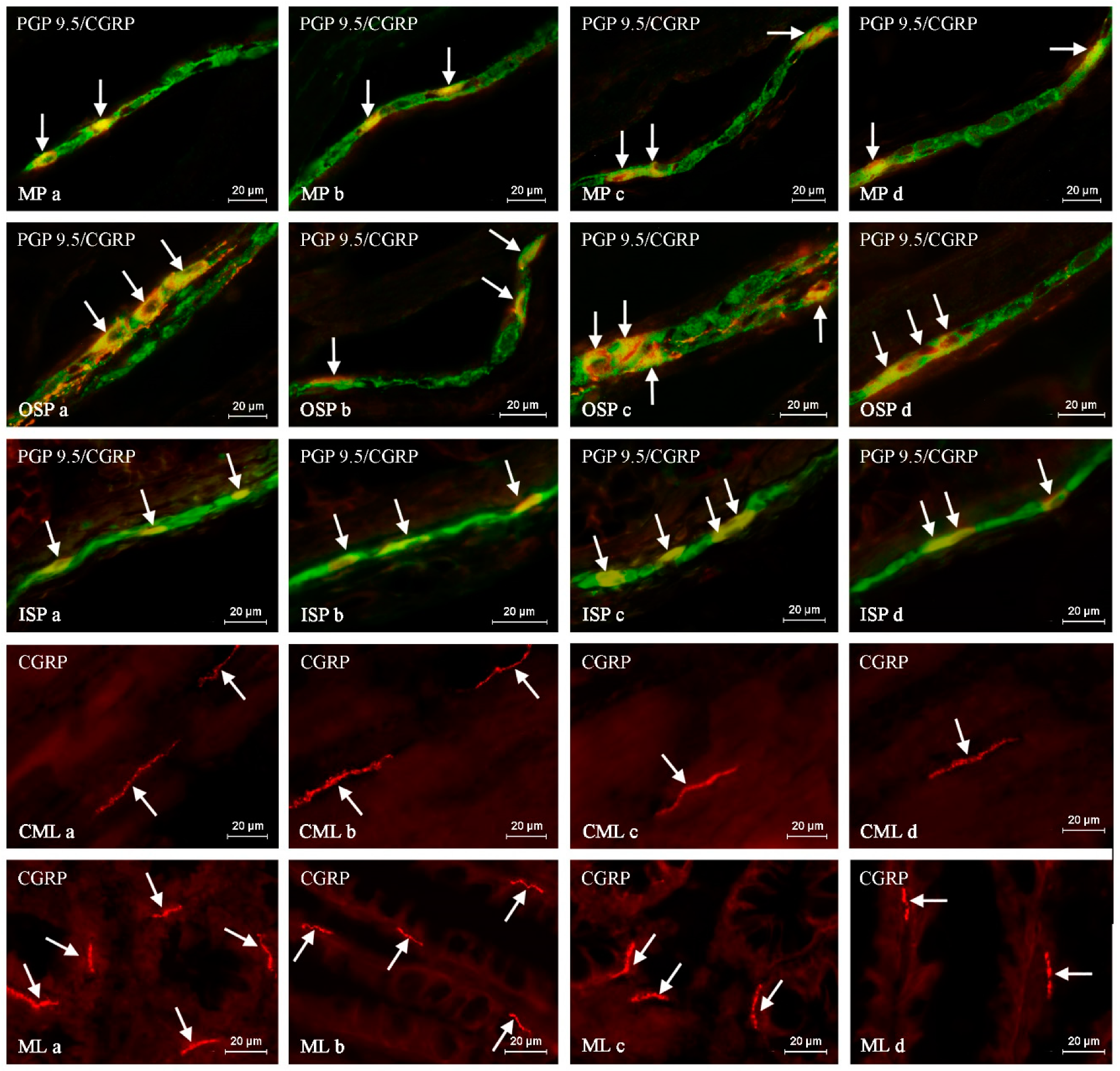
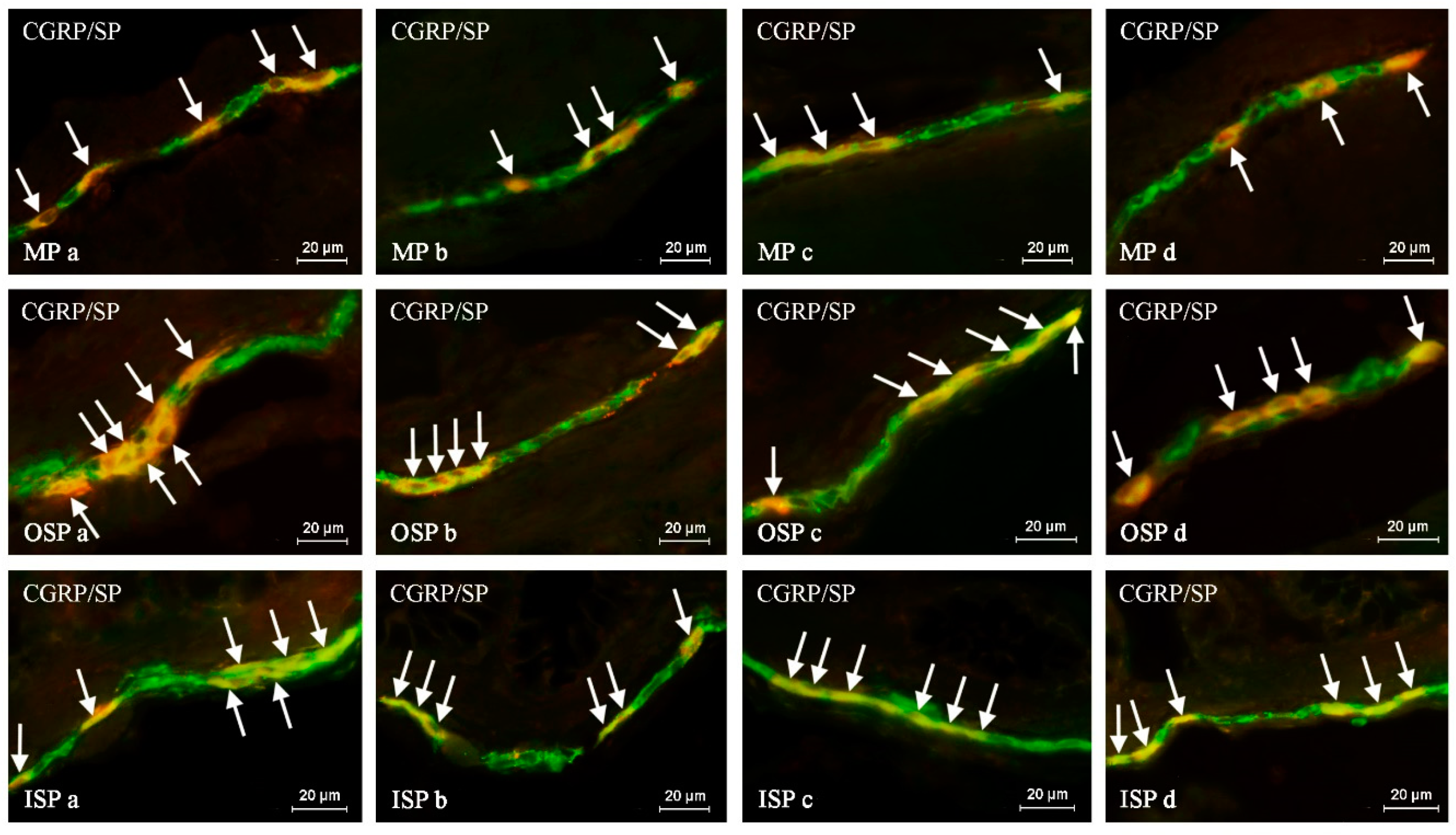
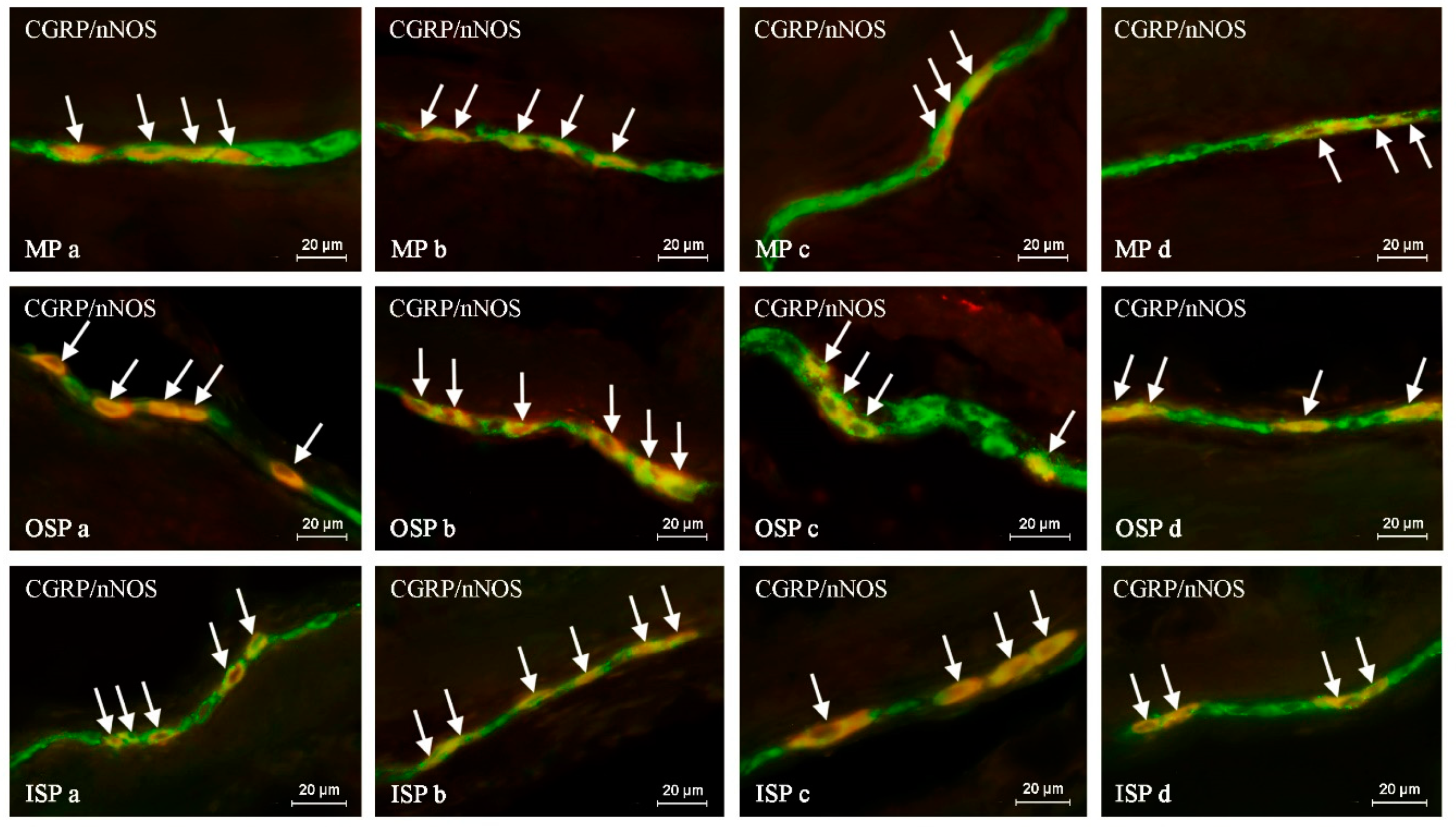
Such prepared descending colon fragments were subjected to the routine double-labeling immunofluorescence method, as described previously by Majewski et al. 2002 [
59].
In brief, this technique was carried out as follows. At room temperature (rt), frozen tissue fragments were dried for 45 minutes and incubated for one hour with a blocking solution (10% goat serum, 0.1% bovine serum albumin (BSA), 0.01% NaN3, Triton X-100, and thimerosal in PBS). Next, the colon fragments were incubated with a mixture of two primary antibodies raised in different species. Depending on the labeling, the primary antisera were directed towards: protein gene product 9.5 (PGP 9.5, used as a pan-neuronal marker), calcitonin gene-related peptide (CGRP), substance P (SP), neuronal form of nitric oxide synthase (nNOS, used as a marker of nitrergic neurons), galanin (GAL), cocaine- and amphetamine-regulated transcript (CART) and vesicular acetylcholine transporter (VAChT, used as a marker of cholinergic neurons). During the incubation process, the slices covered with the mixture of antibodies were left overnight at rt in a humid chamber. The next day, incubation was performed with a mixture of species-specific secondary antisera (conjugated to Alexa Fluor 488 or 546; 1 h, rt), which enabled the visualization of the primary antibodies connected with suitable antigens. Triple rinsing of the samples (15 min. each) after each stage of the described method was made up using PBS (pH 7.4). The specifications and working dilutions of primary and secondary antisera used during the present investigation are presented in
Table 7.
During the present study, routine tests of antibody specificity (including pre-absorption, omission and replacement of primary antibodies by non-immune sera) were performed. These tests completely eliminated specific stainings. It should be pointed out that CGRP occurs in two forms expressed from two genes CALCA and CALCB. Although both antibodies used in the present study are directed against the same form of CGRP (α CGRP encoded by CALCA), they were obtained from different species. Therefore, a comparative study between two anti-CGRP antibodies used during the investigation was performed. This study confirmed that both antibodies against CGRP bind to the same colonic nervous structures in the same pattern and frequency.
The labeled samples were observed under an Olympus BX51 microscope equipped with epi-fluorescence and appropriate filters. Only neurons with clearly-visible nucleus were included in these studies.
The percentage of CGRP-like immunoreactive (LI) neurons was evaluated in relation to nerve cells labeled with PGP-9.5, considered as 100%. At least 500 PGP-9.5-positive neurons were counted and examined for the presence of CGRP in each animal studied, within the MP, OSP and ISP. Moreover, to determine the degree of co-localization of CGRP with other neuronal factors studied, at least 150 CGRP-positive cell bodies in particular types of enteric ganglia were examined for immunoreactivity to each of the other substances and, in this part of the study, cells immunoreactive to CGRP were considered 100%. The obtained results were pooled and presented as mean ± SEM. To prevent the double counting of neurons, the sections included in the study were located at least 150 µm apart.
The density of intra-ganglionic CGRP-LI nerve fibers was evaluated using an arbitrary scale. In this case, the absence of the studied fibers was marked with (−), single fibers with (+), a rare network of fibers with (++), a dense meshwork with (+++) and a very dense meshwork with (++++). In turn, the evaluation of CGRP-positive nerves in the muscular and mucosal layers of the descending colon was based on the counting all CGRP-positive fibers per observation field (0.55 mm2). Nerves were counted in four fragments of the descending colon located at least 150 µm apart per animal (in five observation fields per section) and the obtained data were pooled and presented as a mean. In turn, the denotation of the neurochemical characterization of CGRP-LI nerves in the colonic muscular and mucosal layers was based on the counting of at least 100 nerves immunoreactive to CGRP and by evaluating each of them for immunoreactivity to each of the other studied neuronal factors. The obtained data were also pooled and presented as the mean percentage ± SEM (CGRP-positive nerves were considered to represent 100%). To prevent the double counting of the nerve fibers, in all the above-mentioned methods, the evaluated sections of the colon were located at least 200 μm apart.
A statistical analysis was performed with Student’s t-test (Statistica 12, StatSoft, Inc., Cracow, Poland) and the differences were considered statistically significant at p < 0.05. To evaluate the sex-dependent differences in the ENS, the results obtained from males and females in each age group were compared. Moreover, the results were compared in young and mature animals of the same sex to investigate the effect of the development and maturation of the GI tract on the enteric nervous structures immunoreactive to CGRP.





{kind=link}
{kind=link}
{kind=link}