MEF2A Regulates the MEG3-DIO3 miRNA Mega Cluster-Targeted PP2A Signaling in Bovine Skeletal Myoblast Differentiation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
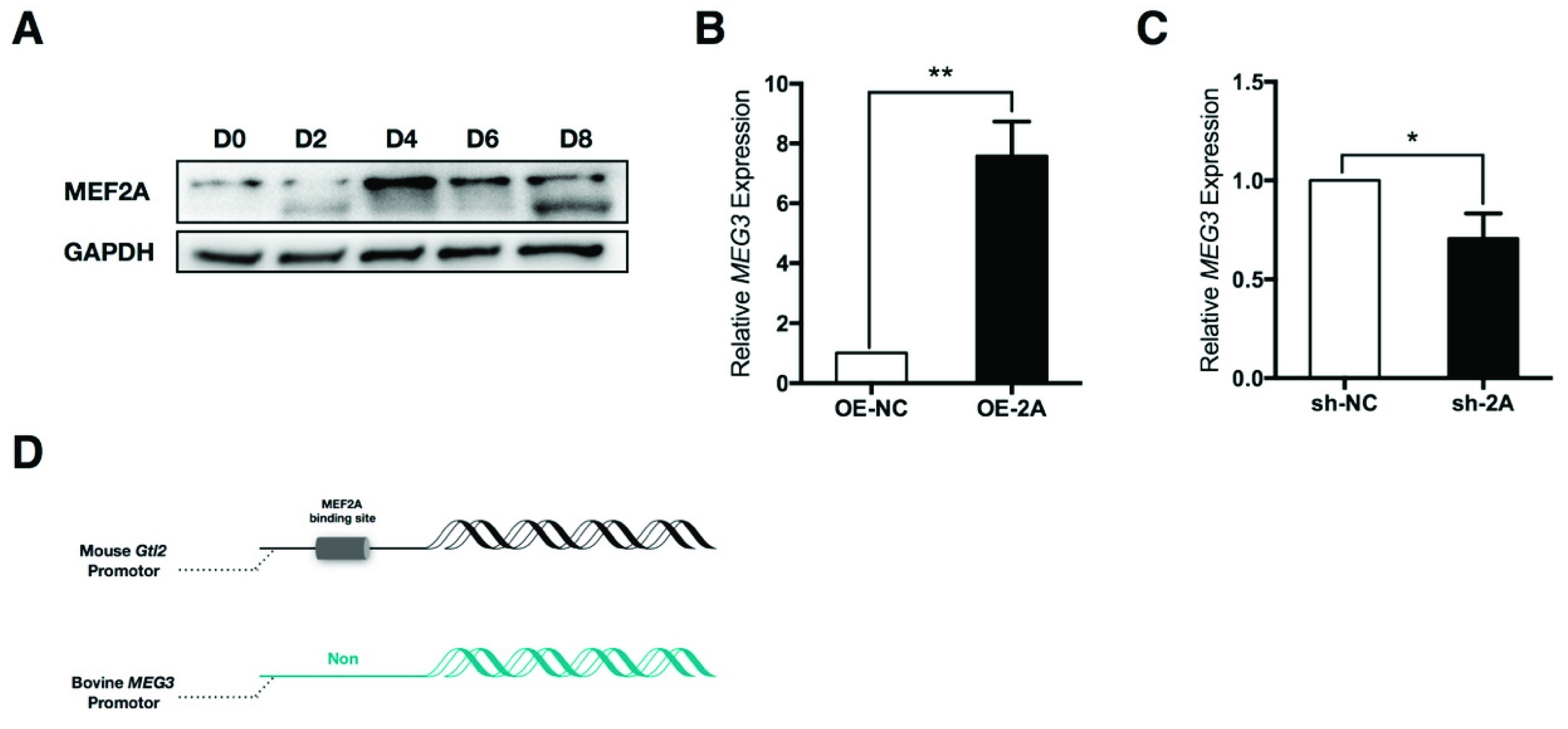
2.1. Myocyte Enhancer Factor 2A (MEF2A) Is Sufficient to Induce Maternally Expressed 3 (MEG3) Expression
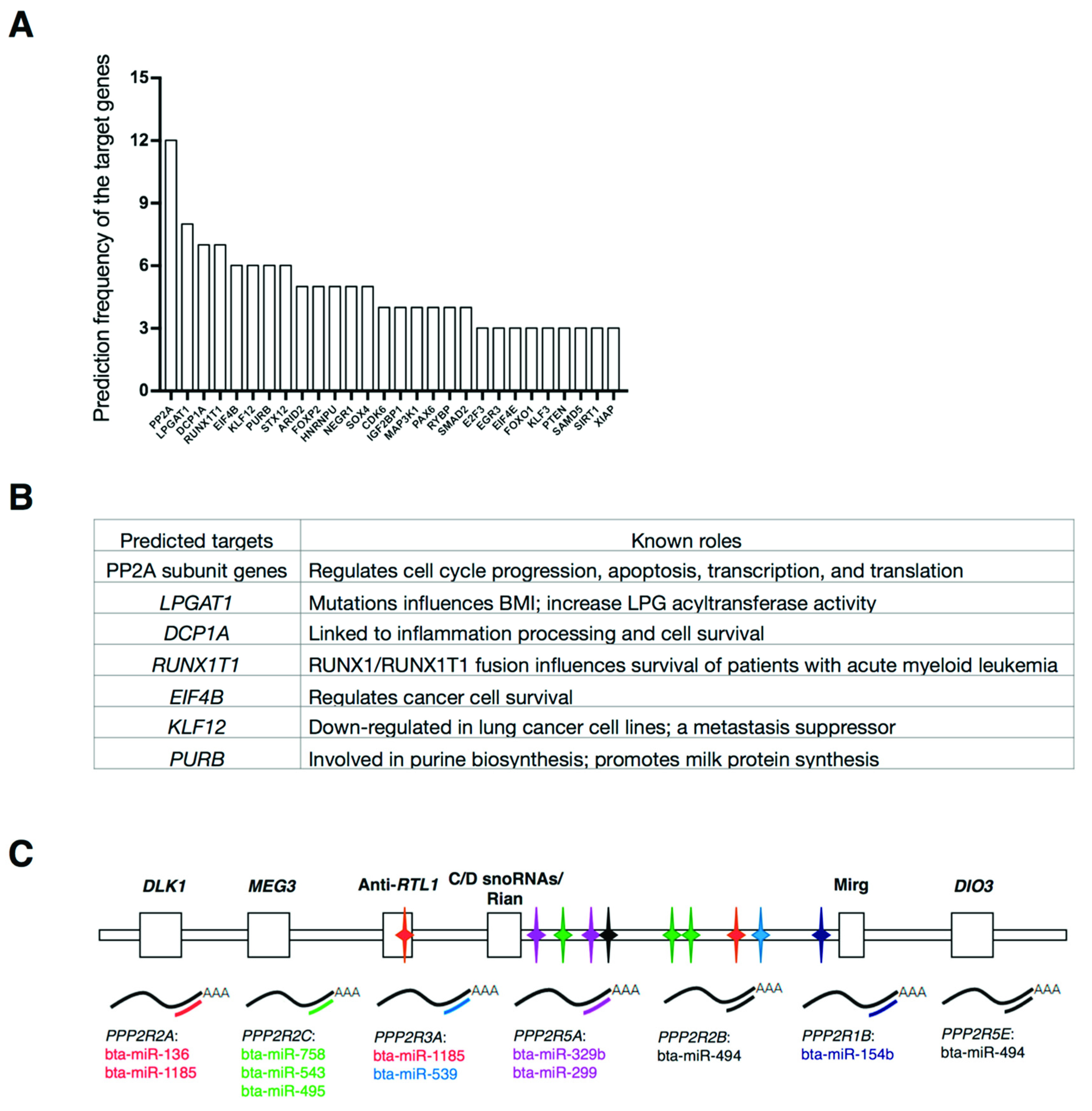
2.2. Identification of Predicted Targets of the Bovine MEG3 - Iodothyronine Deiodinase 3 (DIO3) miRNA Cluster
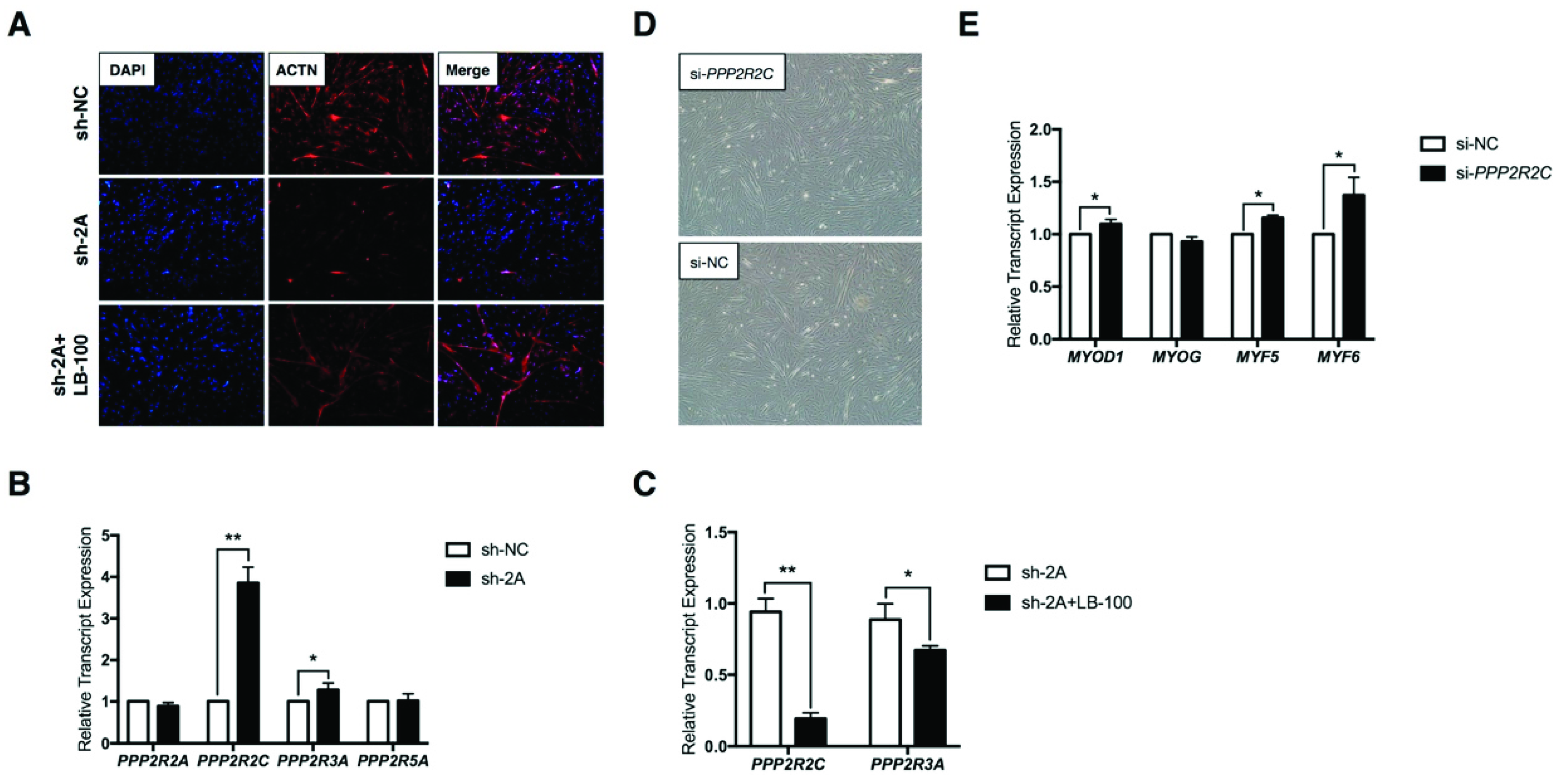
2.3. MEF2A Regulates Protein Phosphatase 2A (PP2A) Signaling During Myoblast Differentiation
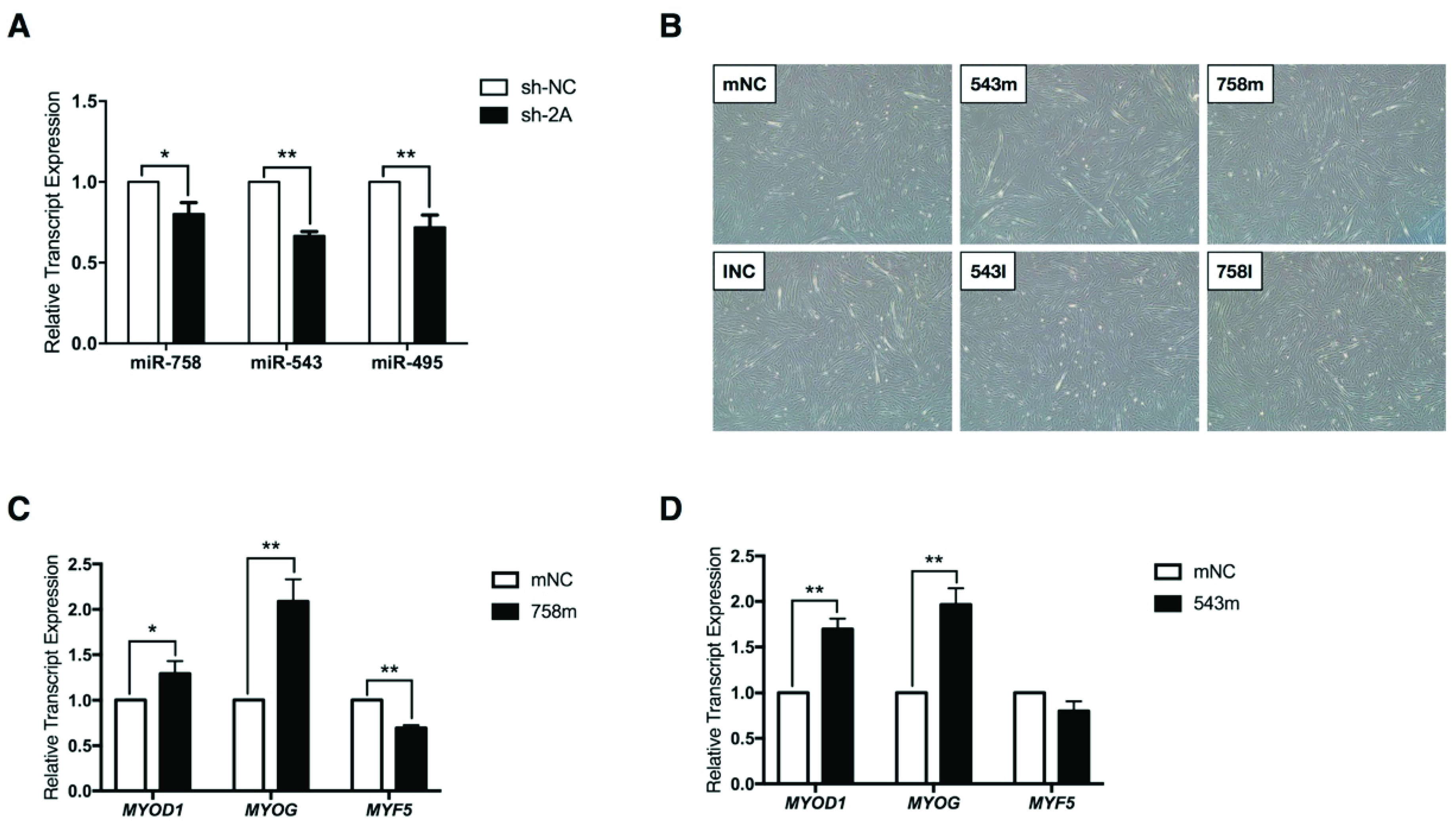
2.4. MEF2A Regulates MEG3-DIO3 miRNAs During Myoblast Differentiation
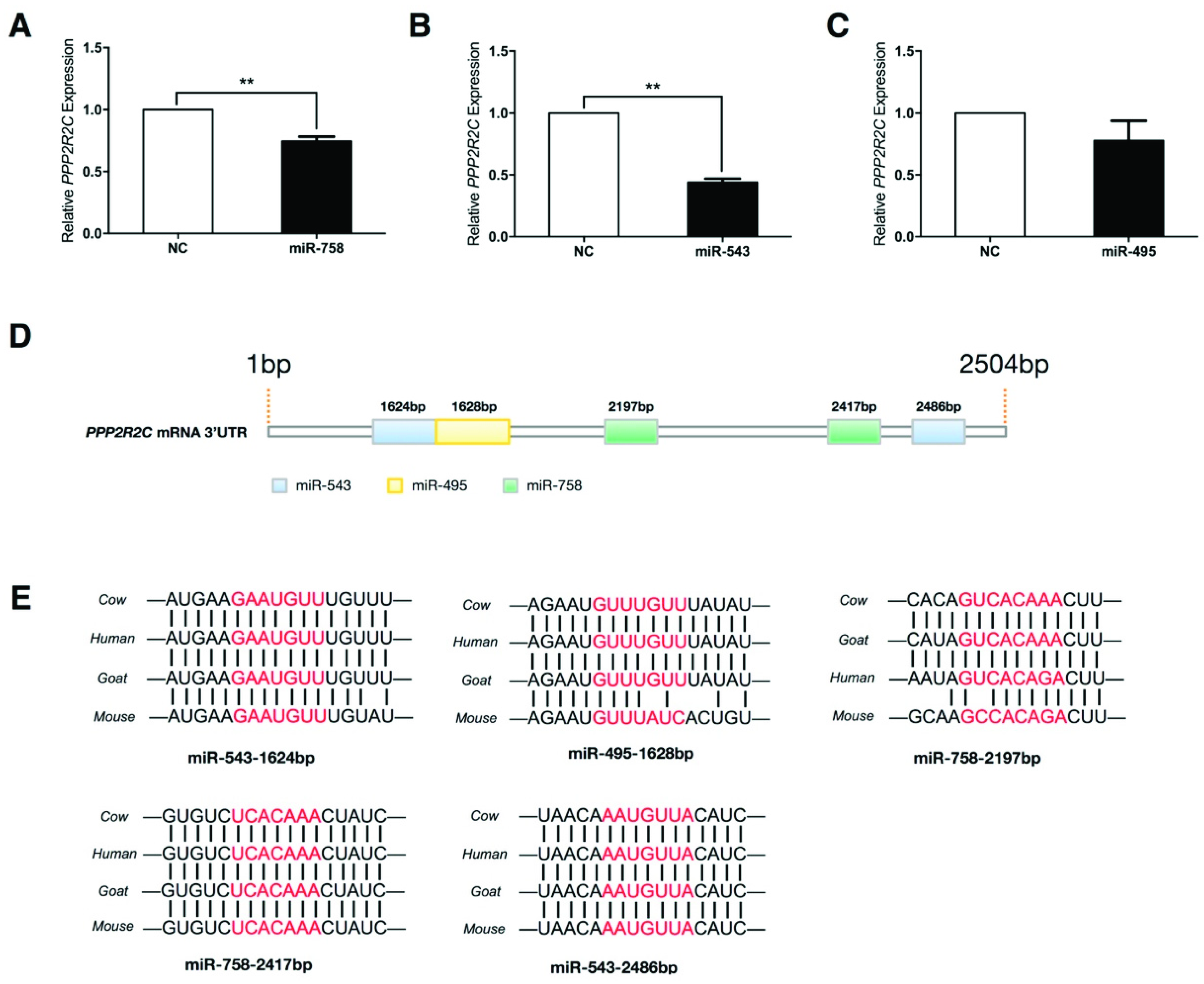
2.5. Protein Phosphatase 2 Regulatory Subunit B, Gamma (PPP2R2C) Expression Is Inhibited by miR-758 and miR-543
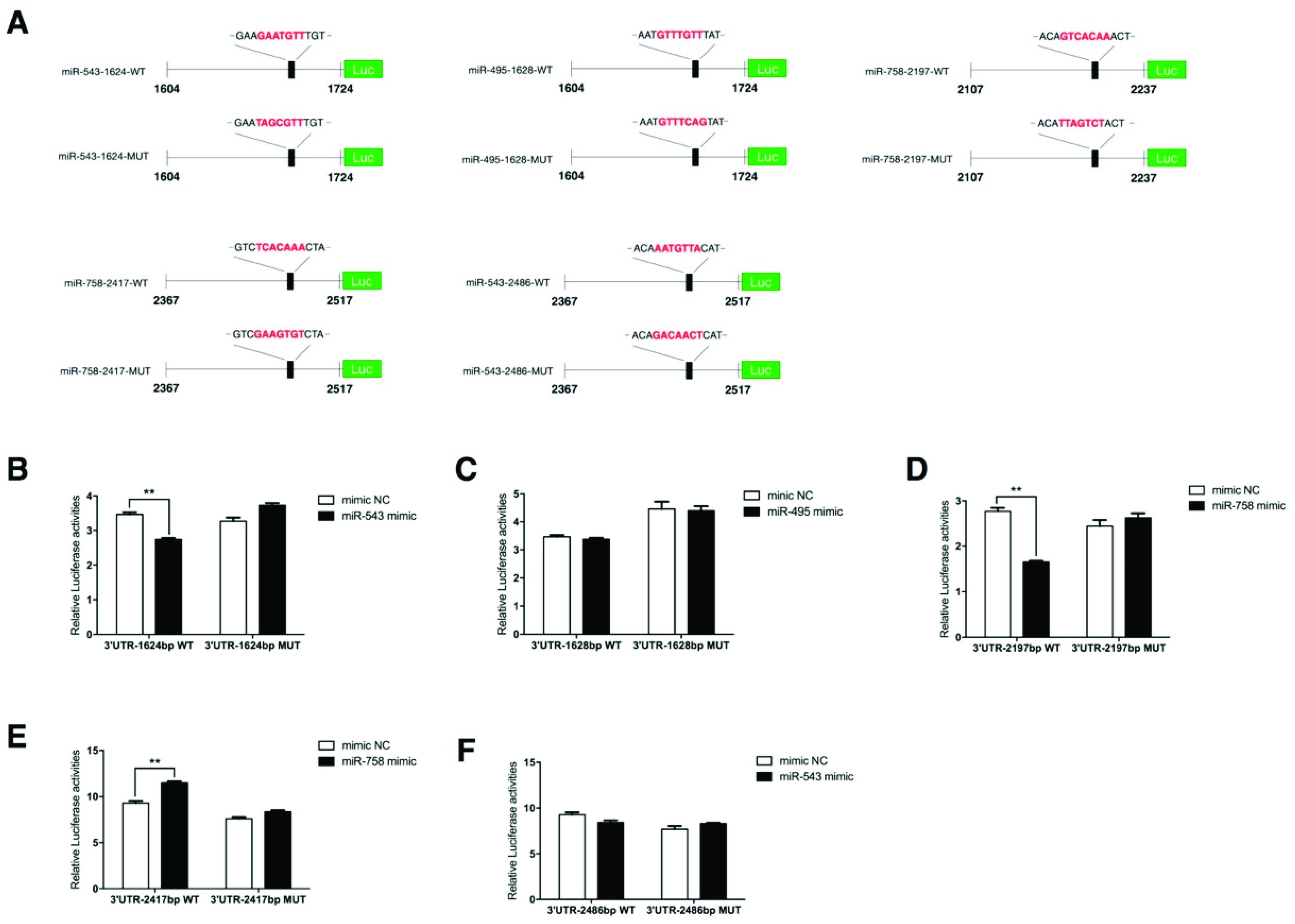
2.6. PPP2R2C Is a Direct Target of miR-758 and miR-543
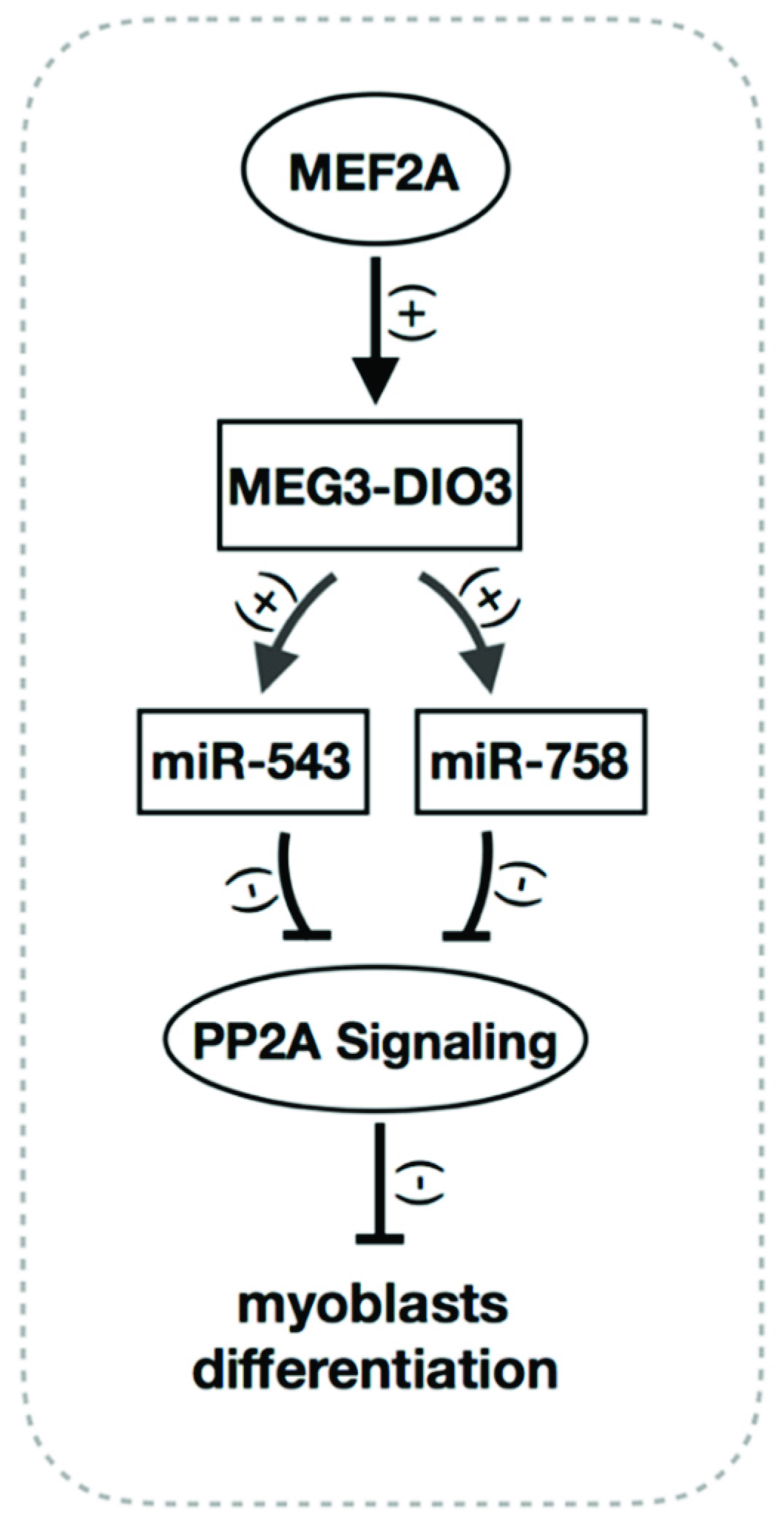
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Adenovirus, Vectors, and RNA Oligonucleotides
4.3. Transfection and Luciferase Activity Assay
4.4. Western Blot Analysis
4.5. Quantitative Real Time-PCR
4.6. Cell Culture Immunofluorescence
4.7. Computational Analysis
4.8. PP2A Inhibitor LB-100 Rescue Assay
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MEF2A | myocyte enhancer factor 2A |
| bHLH | basic helix-loop-helix |
| MAPK | mitogen-activated protein kinase |
| PP2A | protein phosphatase 2A |
| PPP2R2C | protein phosphatase 2 regulatory subunit B, gamma |
| MEG3 | maternally expressed 3 |
| DIO3 | iodothyronine deiodinase 3 |
| ACTN | actinin alpha |
| FBS | fetal bovine serum |
| OE-2A | adenovirus to overexpress MEF2A |
| OE-NC | negative control for OE-2A |
| sh-2A | adenovirus to expression MEF2A shRNA |
| sh-NC | negative control for sh-2A |
| D0 | differentiation day 0 |
| D2 | differentiation day 2 |
| D4 | differentiation day 4 |
| D6 | differentiation day 6 |
| D8 | differentiation day 8 |
| lncRNA | long-noncoding RNA |
| miRNA | microRNA |
| shRNA | short hairpin RNA |
| siRNA | short interference RNA |
| MADS | MCM1, Agamous, Deficiens, Serum Response factor |
| 758m | miR-758 mimics |
| 543m | miR-543 mimics |
| mNC | mimics negative control |
| 758I | miR-758 inhibitor |
| 543I | miR-543 inhibitor |
| INC | inhibitor negative control |
| 3′UTR | 3′ untranslated region |
| WT | wild type |
| MUT | mutant |
| RT-PCR | real time-PCR |
| ANOVA | one-way analysis of variance |
| MOI | multiplicity of infection |
| MADS | MCM1, Agamous, Deficiens, Serum Response factor |
| LPGAT1 | iysophosphatidylglycerol acyltransferase 1 |
| DCP1A | decapping mRNA 1A |
| RUNX1T1 | RUNX1 translocation partner 1 |
| EIF4B | eukaryotic translation initiation factor 4B |
| KLF12 | kruppel like factor 12 |
| PURB | purine rich element binding protein B |
| SDS-PAGE | sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| PBS | phosphate buffer solution |
| DAPI | 4′,6-diamidino-2-phenylindole |
| PVDF | polyvinylidene fluoride |
| PPP2R1B | protein phosphatase 2 scaffold subunit A, beta |
| PPP2R2A | protein phosphatase 2 regulatory subunit B, alpha |
| PPP2R2B | protein phosphatase 2 regulatory subunit B, beta |
| PPP2R5A | protein phosphatase 2 regulatory subunit B′ alpha |
| PPP2R5E | protein phosphatase 2 regulatory subunit B′ epsilon |
| PPP2R3A | protein phosphatase 2 regulatory subunit B″ alpha |
| MRFs | myogenic regulatory factors |
| MYOD1 | myogenic differentiation 1 |
| MYOG | myogenin |
| MYF5 | myogenic factor 5 |
| MYF6 | myogenic regulatory factor 4 |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
References
- Cohen, S.; Nathan, J.A.; Goldberg, A.L. Muscle wasting in disease: Molecular mechanisms and promising therapies. Nat. Rev. Drug Discov. 2015, 14, 58–74. [Google Scholar] [CrossRef] [PubMed]
- Braun, T.; Gautel, M. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis. Nat. Rev. Mol. Cell Biol. 2011, 12, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Rudnicki, M.A. Satellite cells, the engines of muscle repair. Nat. Rev. Mol. Cell Biol. 2011, 13, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, M.; Rigby, P.W. Gene Regulatory Networks and Transcriptional Mechanisms that Control Myogenesis. Dev. Cell 2014, 28, 225–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Nelson, B.R.; Bezprozvannaya, S.; Shelton, J.M.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. Requirement of MEF2A, C, and D for skeletal muscle regeneration. Proc. Natl. Acad. Sci. USA 2014, 111, 4109–4114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.L.; Naya, F.J. MicroRNAs in the Myocyte Enhancer Factor 2 (MEF2)-regulated Gtl2-Dio3 Noncoding RNA Locus Promote Cardiomyocyte Proliferation by Targeting the Transcriptional Coactivator Cited2. J. Biol. Chem. 2015, 290, 23162–23172. [Google Scholar] [CrossRef] [Green Version]
- Estrella, N.L.; Desjardins, C.A.; Nocco, S.E.; Clark, A.L.; Maksimenko, Y.; Naya, F.J. MEF2 transcription factors regulate distinct gene programs in mammalian skeletal muscle differentiation. J. Biol. Chem. 2015, 290, 1256–1268. [Google Scholar] [CrossRef] [PubMed]
- Snyder, C.M.; Rice, A.L.; Estrella, N.L.; Held, A.; Kandarian, S.C.; Naya, F.J. MEF2A regulates the Gtl2-Dio3 microRNA mega-cluster to modulate WNT signaling in skeletal muscle regeneration. Development 2012, 140, 31–42. [Google Scholar] [CrossRef]
- Wang, Y.N.; Yang, W.C.; Li, P.W.; Wang, H.B.; Zhang, Y.Y.; Zan, L.S. Myocyte enhancer factor 2A promotes proliferation and its inhibition attenuates myogenic differentiation via myozenin 2 in bovine skeletal muscle myoblast. PLoS ONE 2018, 13, e0196255. [Google Scholar] [CrossRef]
- McKinsey, T.A.; Zhang, C.L.; Olson, E.N. MEF2: A calcium-dependent regulator of cell division, differentiation and death. Trends Biochem. Sci. 2002, 27, 40–47. [Google Scholar] [CrossRef]
- Schiaffino, S.; Serrano, A.L. Calcineurin signaling and neural control of skeletal muscle fiber type and size. Trends Pharmacol. Sci. 2002, 23, 569–575. [Google Scholar] [CrossRef]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Lagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of tissue-specific microRNAs from mouse. Curr. Biol. 2002, 12, 735–739. [Google Scholar] [CrossRef]
- Chen, J.F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.L.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Williams, A.H.; Kim, Y.; McAnally, J.; Bezprozvannaya, S.; Sutherland, L.B.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. An intragenic MEF2-dependent enhancer directs muscle-specific expression of microRNAs 1 and 133. Proc. Natl. Acad. Sci. USA 2007, 104, 20844–20849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wust, S.; Drose, S.; Heidler, J.; Wittig, I.; Klockner, I.; Franko, A.; Bonke, E.; Gunther, S.; Gartner, U.; Boettger, T.; et al. Metabolic Maturation during Muscle Stem Cell Differentiation Is Achieved by miR-1/133a-Mediated Inhibition of the Dlk1-Dio3 Mega Gene Cluster. Cell Metab. 2018. [Google Scholar] [CrossRef] [PubMed]
- Seitz, H.; Royo, H.; Bortolin, M.L.; Lin, S.P.; Ferguson-Smith, A.C.; Cavaille, J. A large imprinted microRNA gene cluster at the mouse Dlk1-Gtl2 domain. Genome Res. 2004, 14, 1741–1748. [Google Scholar] [CrossRef]
- Da Rocha, S.T.; Edwards, C.A.; Ito, M.; Ogata, T.; Ferguson-Smith, A.C. Genomic imprinting at the mammalian Dlk1-Dio3 domain. Trends Genet. 2008, 24, 306–316. [Google Scholar] [CrossRef]
- Zhou, Y.; Cheunsuchon, P.; Nakayama, Y.; Lawlor, M.W.; Zhong, Y.; Rice, K.A.; Zhang, L.; Zhang, X.; Gordon, F.E.; Lidov, H.G.; et al. Activation of paternally expressed genes and perinatal death caused by deletion of the Gtl2 gene. Development 2010, 137, 2643–2652. [Google Scholar] [CrossRef]
- Perrotti, D.; Neviani, P. Protein phosphatase 2A: A target for anticancer therapy. Lancet Oncol. 2013, 14, e229–e238. [Google Scholar] [CrossRef]
- Kurimchak, A.; Grana, X. PP2A: More than a reset switch to activate pRB proteins during the cell cycle and in response to signaling cues. Cell Cycle 2015, 14, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Sangodkar, J.; Farrington, C.C.; McClinch, K.; Galsky, M.D.; Kastrinsky, D.B.; Narla, G. All roads lead to PP2A: Exploiting the therapeutic potential of this phosphatase. FEBS J. 2016, 283, 1004–1024. [Google Scholar] [CrossRef] [PubMed]
- Grech, G.; Baldacchino, S.; Saliba, C.; Grixti, M.P.; Gauci, R.; Petroni, V.; Fenech, A.G.; Scerri, C. Deregulation of the protein phosphatase 2A, PP2A in cancer: Complexity and therapeutic options. Tumor Biol. 2016, 37, 11691–11700. [Google Scholar] [CrossRef] [PubMed]
- Seshacharyulu, P.; Pandey, P.; Datta, K.; Batra, S.K. Phosphatase: PP2A structural importance, regulation and its aberrant expression in cancer. Cancer Lett. 2013, 335, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, S.; Ehler, E.; Gautel, M. From A to Z and back? Multicompartment proteins in the sarcomere. Trends Cell Biol. 2006, 16, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Wales, S.; Hashemi, S.; Blais, A.; McDermott, J.C. Global MEF2 target gene analysis in cardiac and skeletal muscle reveals novel regulation of DUSP6 by p38MAPK-MEF2 signaling. Nucleic Acids Res. 2014, 42, 11349–11362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Cao, J.; Shi, Y. Identification and characterization of a gene encoding human LPGAT1, an endoplasmic reticulum-associated lysophosphatidylglycerol acyltransferase. J. Biol. Chem. 2004, 279, 55866–55874. [Google Scholar] [CrossRef]
- Zhang, Y.; Morar, M.; Ealick, S.E. Structural biology of the purine biosynthetic pathway. Cell. Mol. Life Sci. 2008, 65, 3699–3724. [Google Scholar] [CrossRef] [Green Version]
- Traurig, M.T.; Orczewska, J.I.; Ortiz, D.J.; Bian, L.; Marinelarena, A.M.; Kobes, S.; Malhotra, A.; Hanson, R.L.; Mason, C.C.; Knowler, W.C.; et al. Evidence for a Role of LPGAT1 in Influencing BMI and Percent Body Fat in Native Americans. Obesity 2012. [Google Scholar] [CrossRef] [Green Version]
- Godin-Heymann, N.; Brabetz, S.; Murillo, M.M.; Saponaro, M.; Santos, C.R.; Lobley, A.; East, P.; Chakravarty, P.; Matthews, N.; Kelly, G.; et al. Tumour-suppression function of KLF12 through regulation of anoikis. Oncogene 2015, 35, 3324–3334. [Google Scholar] [CrossRef] [Green Version]
- Seto, E.; Yoshida-Sugitani, R.; Kobayashi, T.; Toyama-Sorimachi, N. The Assembly of EDC4 and Dcp1a into Processing Bodies Is Critical for the Translational Regulation of IL-6. PLoS ONE 2015, 10, e0123223. [Google Scholar] [CrossRef]
- Hornung, R.; Jurinovic, V.; Batcha, A.M.N.; Bamopoulos, S.A.; Rothenberg-Thurley, M.; Amler, S.; Sauerland, M.C.; Berdel, W.E.; Wörmann, B.J.; Bohlander, S.K.; et al. Mediation analysis reveals common mechanisms of RUNX1 point mutations and RUNX1/RUNX1T1 fusions influencing survival of patients with acute myeloid leukemia. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Huo, N.; Yu, M.; Li, X.; Zhou, C.; Jin, X.; Gao, X. PURB is a positive regulator of amino acid-induced milk synthesis in bovine mammary epithelial cells. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef]
- Kapadia, B.; Nanaji, N.M.; Bhalla, K.; Bhandary, B.; Lapidus, R.; Beheshti, A.; Evens, A.M.; Gartenhaus, R.B. Fatty Acid Synthase induced S6Kinase facilitates USP11-eIF4B complex formation for sustained oncogenic translation in DLBCL. Nat. Commun. 2018, 9, 829. [Google Scholar] [CrossRef] [PubMed]
- Dragomir, M.P.; Knutsen, E.; Calin, G.A. SnapShot: Unconventional miRNA Functions. Cell 2018, 174, 1038–1038.E1. [Google Scholar] [CrossRef]
- Wu, H.; Naya, F.J.; McKinsey, T.A.; Mercer, B.; Shelton, J.M.; Chin, E.R.; Simard, A.R.; Michel, R.N.; Bassel-Duby, R.; Olson, E.N.; et al. MEF2 responds to multiple calcium-regulated signals in the control of skeletal muscle fiber type. EMBO J. 2000, 19, 1963–1973. [Google Scholar] [CrossRef] [PubMed]
- Potthoff, M.J.; Olson, E.N. MEF2: A central regulator of diverse developmental programs. Development 2007, 134, 4131–4140. [Google Scholar] [CrossRef] [PubMed]
- Creemers, E.E.; Sutherland, L.B.; Oh, J.; Barbosa, A.C.; Olson, E.N. Coactivation of MEF2 by the SAP domain proteins myocardin and MASTR. Mol. Cell 2006, 23, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Virshup, D.M.; Shenolikar, S. From promiscuity to precision: Protein phosphatases get a makeover. Mol. Cell 2009, 33, 537–545. [Google Scholar] [CrossRef]
- McCright, B.; Rivers, A.M.; Audlin, S.; Virshup, D.M. The B56 family of protein phosphatase 2A (PP2A) regulatory subunits encodes differentiation-induced phosphoproteins that target PP2A to both nucleus and cytoplasm. J. Biol. Chem. 1996, 271, 22081–22089. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Mei, C.; Su, X.; Wang, H.; Yang, W.; Zan, L. MEF2A Regulates the MEG3-DIO3 miRNA Mega Cluster-Targeted PP2A Signaling in Bovine Skeletal Myoblast Differentiation. Int. J. Mol. Sci. 2019, 20, 2748. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20112748
Wang Y, Mei C, Su X, Wang H, Yang W, Zan L. MEF2A Regulates the MEG3-DIO3 miRNA Mega Cluster-Targeted PP2A Signaling in Bovine Skeletal Myoblast Differentiation. International Journal of Molecular Sciences. 2019; 20(11):2748. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20112748
Chicago/Turabian StyleWang, Yaning, Chugang Mei, Xiaotong Su, Hongbao Wang, Wucai Yang, and Linsen Zan. 2019. "MEF2A Regulates the MEG3-DIO3 miRNA Mega Cluster-Targeted PP2A Signaling in Bovine Skeletal Myoblast Differentiation" International Journal of Molecular Sciences 20, no. 11: 2748. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20112748