Flavonoids from Chionanthus retusus (Oleaceae) Flowers and Their Protective Effects against Glutamate-Induced Cell Toxicity in HT22 Cells

 ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Contents of Total Phenols and Total Flavonoids in C. retusus Flowers
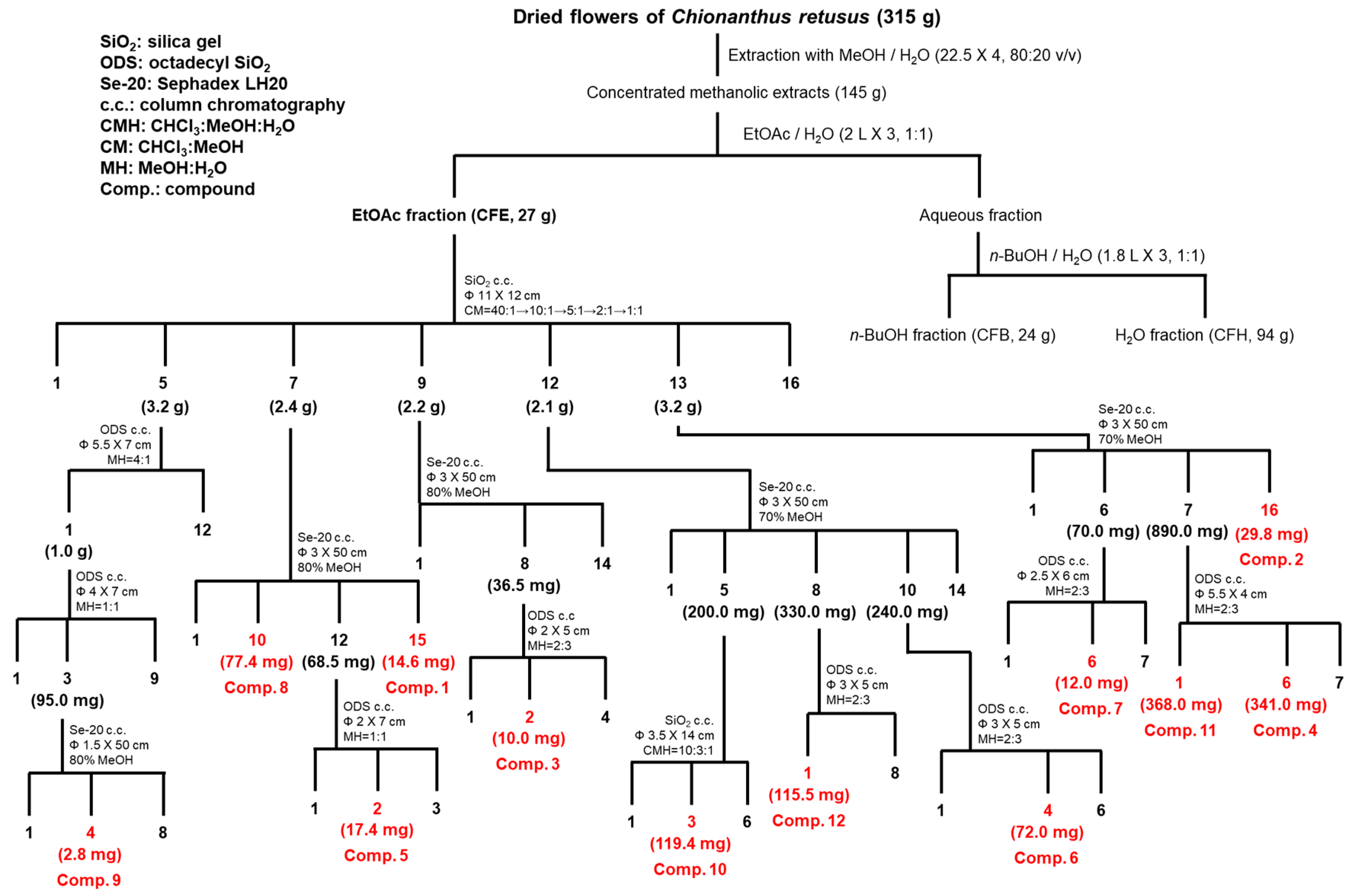
2.2. Isolation and Identification of Flavonoids from C. retusus Flowers
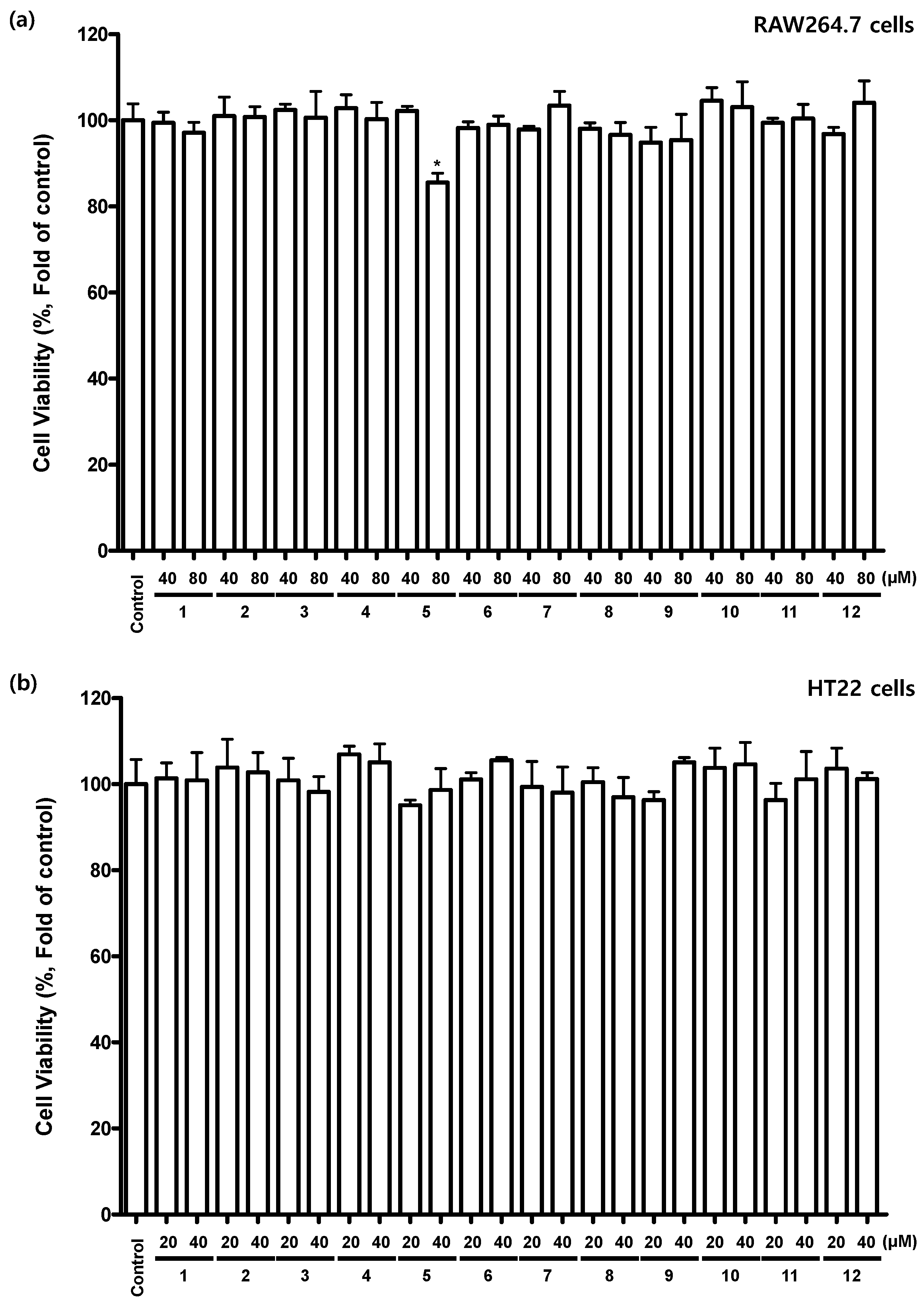
2.3. Inhibition Effects of Flavonoids 1–12 on NO Production in Lipopolysaccharide (LPS)-Induced RAW 264.7 Cells
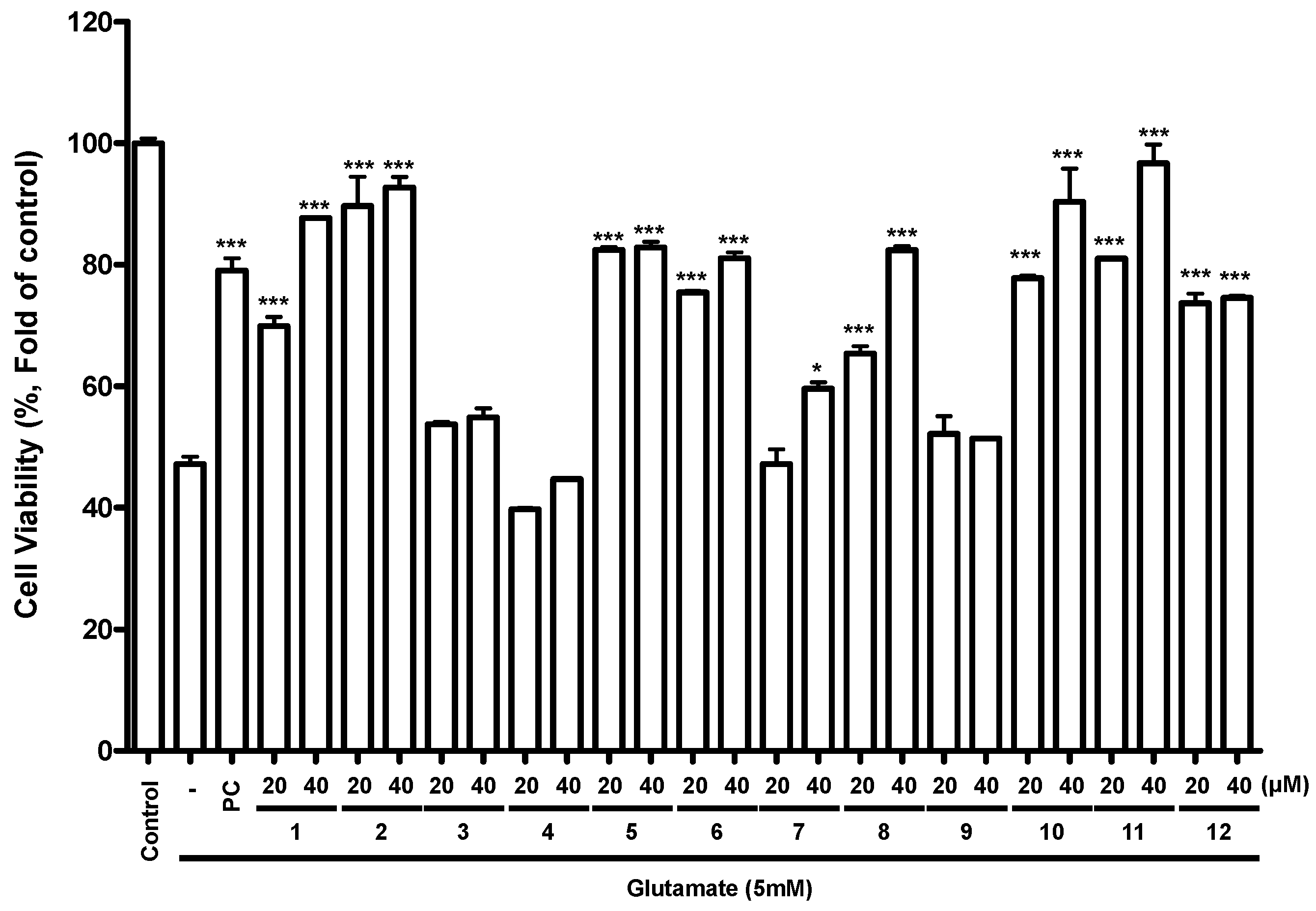
2.4. Effects of Flavonoids 1–12 on Glutamate-Induced Cell Toxicity in Mouse Hippocampal HT22 Cells
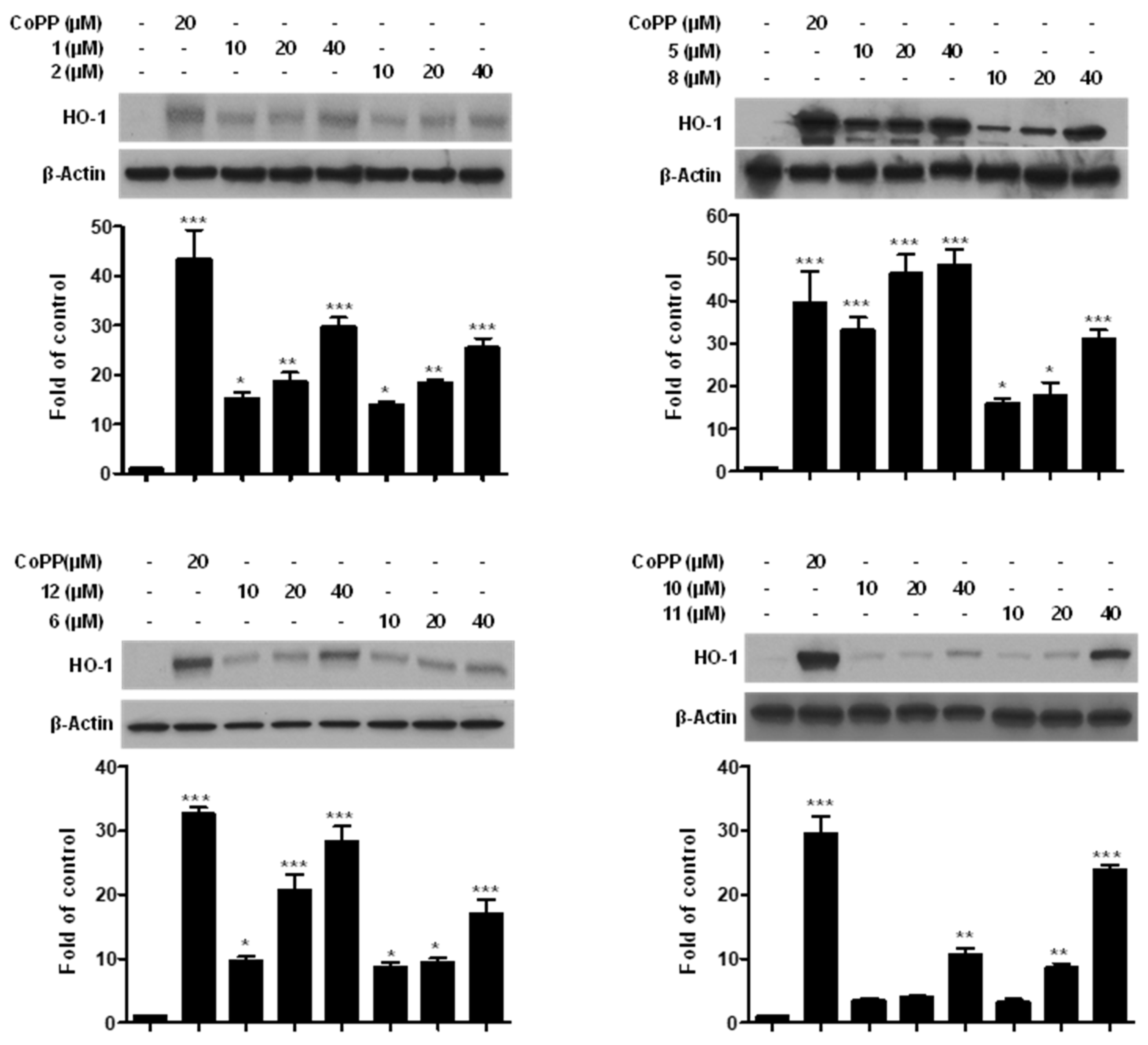
2.5. Effects of Compounds 1, 2, 5, 6, 8, and 10–12 on HO-1 Expression in Mouse Hippocampal HT22 Cells
2.6. Effects of Compounds 1, 2, 5, 6, 8, and 10–12 on Cell Viability through HO Signaling Pathway
3. Materials and Methods
3.1. Plant Materials
3.2. General Experimental Procedures
3.3. Isolation Procedure of Flavonoids (1–12) from C. retusus Flowers
3.4. Cell Culture and MTT Assay
3.5. Macrophage RAW 264.7 Culture, Viability Assay, and NO Measurement
3.6. Determination of Total Phenols and Flavonoids Contents in C. retusus Flower
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| c.c. | column chromatography |
| CoPP | cobalt protoporphyrin |
| Fr | fraction |
| HO | heme oxygenase |
| IR | infrared |
| SnPP | tin protoporphyrin IX |
| SiO2 | silica gel |
| ODS | octadecyl SiO2 |
| PC | positive control |
| TLC | thin layer chromatography |
| UV | ultraviolet |
| Ve/Vt | elution volume/total volume |
References
- Han, A.R.; Park, S.A.; Ahn, B.E. Reduced stress and improved physical functional ability in elderly with mental health problems following a horticultural therapy program. Complement. Ther. Med. 2018, 38, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.F.; Braidy, N.; Habtemariam, S.; Orhan, I.E.; Daglia, M.; Manayi, A.; Gortzi, O.; Nabavi, S.M. Neuroprotective effects of chrysin: From chemistry to medicine. Neurochem. Int. 2015, 90, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Levi, M.S.; Brimble, M.A. A review of neuroprotective agents. Curr. Med. Chem. 2004, 11, 2383–2397. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.; Lee, M.H.; Lee, S.H.; Cho, E.J. Alpha-linolenic acid regulates amyloid precursor protein processing by mitogen-activated protein kinase pathway and neuronal apoptosis in amyloid beta-induced SH-SY5Y neuronal cells. Appl. Biol. Chem. 2018, 61, 61–71. [Google Scholar] [CrossRef]
- Vauzour, D.; Vafeiadou, K.; Rodriguez-Mateos, A.; Rendeiro, C.; Spencer, J.P.E. The neuroprotective potential of flavonoids: A multiplicity of effects. Genes Nutr. 2008, 3, 91. [Google Scholar] [CrossRef] [PubMed]
- Dajas, F.; Rivera-Megret, F.; Blasina, F.; Arredondo, F.; Abin-Carriquiry, J.A.; Costa, G.; Echeverry, C.; Lafon, L.; Heizen, H.; Ferreira, M.; et al. Neuroprotection by flavonoids. Braz. J. Med. Biol. Res. 2003, 36, 1613–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spagnuolo, C.; Moccia, S.; Russo, G.L. Anti-inflammatory effects of flavonoids in neurodegenerative disorders. Eur. J. Med. Chem. 2018, 153, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Dok-Go, H.; Lee, K.H.; Kim, H.J.; Lee, E.H.; Lee, J.Y.; Song, Y.S.; Lee, Y.H.; Jin, C.B.; Lee, Y.S.; Cho, J.S. Neuroprotective effects of antioxidative flavonoids, quercetin, (+)-dihydroquercetin and quercetin 3-methyl ether, isolated from Opuntia ficus-indica var. saboten. Brain Res. 2003, 965, 130–136. [Google Scholar] [CrossRef]
- Hwang, S.L.; Shih, P.H.; Yen, G.C. Neuroprotective effects of citrus flavonoids. J. Agric. Food Chem. 2012, 60, 877–885. [Google Scholar] [CrossRef]
- Kang, H.M.; Kim, J.H.; Lee, M.Y.; Son, K.H.; Yang, D.C.; Baek, N.I.; Kwon, B.M. Relationship between flavonoid structure and inhibition of farnesyl protein transferase. Nat. Prod. Res. 2004, 18, 349–356. [Google Scholar] [CrossRef]
- Yang, H.J.; Shin, Y.J. Antioxidant compounds and activities of edible roses (Rosa hybrida spp.) from different cultivars grown in Korea. Appl. Biol. Chem. 2017, 60, 129–136. [Google Scholar] [CrossRef]
- Lee, J.M.; Rodriguez, J.P.; Quilantang, N.G.; Lee, M.H.; Cho, E.J.; Jacinto, S.D.; Lee, S.H. Determination of flavonoids from Perilla frutescens var. japonica seeds and their inhibitory effect on aldose reductase. Appl. Biol. Chem. 2017, 60, 155–162. [Google Scholar] [CrossRef]
- Rodriguez, J.P.; Lee, J.M.; Park, J.Y.; Kang, K.S.; Hahm, D.H.; Lee, S.C.; Lee, S.H. HPLC-UV analysis of sample preparation influence on flavonoid yield from Cirsium japonicum var. maackii. Appl. Biol. Chem. 2017, 60, 519–525. [Google Scholar] [CrossRef]
- Dötterl, S.; Vereecken, N.J. The chemical ecology and evolution of bee-flower interactions: A review and perspectives. Can. J. Zool. 2010, 88, 668–697. [Google Scholar] [CrossRef]
- Raguso, R.A. Wake up and smell the roses: The ecology and evolution of floral scent. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 549–569. [Google Scholar] [CrossRef]
- Wright, G.A.; Schiestl, F.P. The evolution of floral scent: The influence of olfactory learning by insect pollinators on the honest signalling of floral rewards. Funct. Ecol. 2009, 23, 841–851. [Google Scholar] [CrossRef]
- Sun, X.M.; Li, X.F.; Deng, R.X.; Liu, Y.Q.; Hou, X.W.; Xing, Y.P.; Liu, P. Extraction technology and antioxidant activity of total flavonoids from the flower of Chionanthus retusa. Food Sci. 2015, 36, 266–271, 278. [Google Scholar]
- Lee, Y.N.; Jeong, C.H.; Shim, K.H. Isolation of antioxidant and antibrowning substance from Chionanthus retusa leaves. J. Korean Soc. Food Sci. Nutr. 2004, 33, 1419–1425. [Google Scholar]
- Deng, R.X.; Zhang, C.F.; Liu, P.; Duan, W.L.; Yin, W.P. Separation and identification of flavonoids from Chinese Fringetree Flowers (Chionanthus retusa Lindl et Paxt). Food Sci. 2014, 35, 74–78. [Google Scholar]
- Kwak, J.H.; Kang, M.W.; Roh, J.H.; Choi, S.U.; Zee, O.P. Cytotoxic phenolic compounds from Chionanthus retusus. Arch. Pharm. Res. 2009, 32, 1681–1687. [Google Scholar] [CrossRef]
- Baek, Y.S.; Song, N.Y.; Nam, T.G.; Kim, D.O.; Kang, H.C.; Kwon, O.K.; Baek, N.I. Flavonoids from Fragaria ananassa calyx and their antioxidant capacities. Appl. Biol. Chem. 2015, 58, 787–793. [Google Scholar] [CrossRef]
- Han, J.T.; Bang, M.H.; Chun, O.K.; Kim, D.O.; Lee, C.Y.; Baek, N.I. Flavonol glycosides from the aerial parts of Aceriphyllum rossii and their antioxidant activities. Arch. Pharm. Res. 2004, 27, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Braca, A.; Tommasi, N.D.; Bari, L.D.; Pizza, C.; Politi, M.; Morelli, I. Antioxidant principles from Bauhinia tarapotensis. J. Nat. Prod. 2001, 64, 892–895. [Google Scholar] [CrossRef] [PubMed]
- Weber, B.; Herrmann, M.; Hartmann, B.; Joppe, H.; Schmidt, C.O.; Bertram, H.J. HPLC/MS and HPLC/NMR as hyphenated techniques for accelerated characterization of the main constituents in Chamomile (Chamomilla recutita [L.] Rauschert). Eur. Food Res. Technol. 2008, 226, 755–760. [Google Scholar] [CrossRef]
- Shrestha, S.; Lee, D.Y.; Park, J.H.; Cho, J.G.; Lee, D.S.; Li, B.; Kim, Y.C.; Jeon, Y.J.; Yeon, S.W.; Baek, N.I. Flavonoids from the fruits of Nepalese sumac (Rhus parviflora) attenuate glutamate-induced neurotoxicity in HT22 cells. Food Sci. Biotechnol. 2013, 22, 895–902. [Google Scholar] [CrossRef]
- Hiep, N.T.; Kwon, J.Y.; Kim, D.W.; Hong, S.G.; Guo, Y.Q.; Hwang, B.Y.; Kim, N.H.; Mar, W.C.; Lee, D. Neuroprotective constituents from the fruits of Maclura tricuspidata. Tetrahedron 2017, 73, 2747–2759. [Google Scholar] [CrossRef]
- Latté, K.P.; Ferreira, D.; Venkatraman, M.S.; Kolodziej, H. O-Galloyl-C-glycosylflavones from Pelargonium reniforme. Phytochemistry 2002, 59, 419–424. [Google Scholar] [CrossRef]
- Simonian, N.A.; Coyle, J.T. Oxidative stress in neurodegenerative diseases. Annu. Rev. Pharmacol. Toxicol. 1996, 36, 83–106. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Jeong, G.S. Butein provides neuroprotective and anti-neuroinflammatory effects through Nrf2/ARE-dependent haem oxygenase 1 expression by activating the PI3K/Akt pathway. Br. J. Pharmacol. 2016, 173, 2894–2909. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.S.; Lee, J.N.; Kwon, D.Y.; Kim, M.S. Ondamtanggamibang protects neurons from oxidative stress with induction of heme oxygenase-1. J. Ethnopharmacol. 2006, 108, 294–298. [Google Scholar] [CrossRef]
- Choi, B.M.; Kim, H.J.; Oh, G.S.; Pae, H.O.; Oh, H.C.; Jeong, S.J.; Kwon, T.O.; Kim, Y.M.; Chung, H.T. 1,2,3,4,6-Penta-O-galloyl-beta-d-glucose protects rat neuronal cells (Neuro 2A) from hydrogen peroxide-mediated cell death via the induction of heme oxygenase-1. Neurosci. Lett. 2002, 328, 185–189. [Google Scholar] [CrossRef]
- Nhan, N.T.; Song, H.S.; Oh, E.J.; Lee, Y.G.; Ko, J.H.; Kwon, J.E.; Kang, S.C.; Lee, D.Y.; Baek, N.I. Phenylpropanoids from Lilium Asiatic hybrid flowers and their anti-inflammatory activities. Appl. Biol. Chem. 2017, 60, 527–533. [Google Scholar]
- Lee, Y.G.; Lee, D.G.; Gwag, J.E.; Kim, M.S.; Kim, M.J.; Kim, H.G.; Ko, J.H.; Yeo, H.J.; Kang, S.H.; Baek, N.I. A 1,1′-biuracil from Epidermidibacterium keratini EPI-7 shows anti-aging effects on human dermal fibroblasts. Appl. Biol. Chem. 2019, 62, 14. [Google Scholar] [CrossRef]
- Lee, Y.G.; Rodriguez, I.; Nam, Y.H.; Gwag, J.E.; Woo, S.H.; Kim, H.G.; Ko, J.H.; Hong, B.N.; Kang, T.H.; Baek, N.I. Recovery effect of lignans and fermented extracts from Forsythia koreana flowers on pancreatic islets damaged by alloxan in zebrafish (Danio rerio). Appl. Biol. Chem. 2019, 62, 7. [Google Scholar] [CrossRef]
- Lee, Y.G.; Seo, K.H.; Lee, D.S.; Gwag, J.E.; Kim, H.G.; Ko, J.H.; Park, S.H.; Lee, D.Y.; Baek, N.I. Phenylethanoid glycoside from Forsythia koreana (Oleaceae) flowers shows a neuroprotective effect. Braz. J. Bot. 2018, 41, 523–528. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Lee, Y.G.; Lee, J.H.; Lee, N.Y.; Kim, N.K.; Jung, D.W.; Wang, W.; Kim, Y.S.; Kim, H.G.; Nguyen, N.T.; Park, H.S.; et al. Evaluation for the flowers of compositae plants as whitening cosmetics functionality. J. Appl. Biol. Chem. 2017, 60, 5–11. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Extract | EtOAc fr. | n-BuOH fr. | H2O fr. |
|---|---|---|---|---|
| Total phenols (mg GA/g DW) | 125.4 ± 3.3 | 245.6 ± 5.2 | 130.1 ± 2.5 | 53.1 ± 1.8 |
| Total flavonoids (mg CA/g DW) | 119.1 ± 2.7 | 172.1 ± 2.1 | 98.2 ± 0.9 | 18.2 ± 1.2 |
| No. | IC50 (μM) | No. | IC50 (μM) | No. | IC50 (μM) |
|---|---|---|---|---|---|
| 1 | 37.93 ± 0.03 | 5 | 5.99 ± 0.02 | 9 | >100 |
| 2 | 21.25 ± 0.03 | 6 | 30.60 ± 0.05 | 10 | 71.56 ± 0.08 |
| 3 | >100 | 7 | >100 | 11 | 57.18 ± 0.03 |
| 4 | >100 | 8 | 78.53 ± 0.03 | 12 | 60.86 ± 0.01 |
| No. | 1 (a) | 2 (a) | 3 (a) | 4 (a) | 5 (b) | 6 (b) | 7 (b) | 8 (a) | 9 (a) | 10 (a) | 11 (a) | 12 (a) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | - | - | - | - | - | - | - | 5.09, d, 12.0 | 5.05, d, 11.6 | 5.03, d, 12.0 | 5.09, d, 11.6 | 5.14, dd, 12.4, 2.8 |
| 3 | - | - | - | - | 6.91, s | 6.90, s | 6.80, s | 4.71, d, 12.0 | 4.58, d, 11.6 | 4.59, d, 12.0 | 4.71, d, 11.6 | 3.00, dd, 16.4, 12.4 |
| 2.63, dd, 16.4, 2.8 | ||||||||||||
| 6 | 6.17, brs | 6.18, brs | 6.20, brs | 6.20, brs | 6.75, d, 1.6 | 6.75, brs | 6.68, brs | 6.05, d, 1.2 | 5.87, d, 2.0 | 6.15, d, 1.6 | 6.05, d, 1.2 | 5.09, brs |
| 8 | 6.38, brs | 6.38, brs | 6.36, brs | 6.37, brs | 6.76, d, 1.6 | 6.77, brs | 6.78, brs | 6.13, d, 1.2 | 5.89, d, 2.0 | 6.21, d, 1.6 | 6.09, d, 1.2 | 6.00, brs |
| 2′ | 7.73, d, 1.8 | 8.10, d, 8.4 | 8.06, d, 8.0 | 8.07, d, 8.0 | 7.83, brs | 7.85, d, 1.2 | 7.87, d, 7.6 | 7.20, d, 1.2 | 7.29, d, 8.4 | 7.38, d, 8.0 | 7.18, d, 1.2 | 6.73, brs |
| 3′ | - | 6.91, d, 8.4 | 6.90, d, 8.0 | 6.90, d, 8.0 | - | - | 7.19, d, 7.6 | - | 6.80, d, 8.4 | 6.85, d, 8.0 | - | - |
| 5′ | 6.88, d, 8.4 | 6.91, d, 8.4 | 6.90, d, 8.0 | 6.90, d, 8.0 | 7.39, d, 8.0 | 7.36, d, 8.0 | 7.19, d, 7.6 | 7.01, d, 7.6 | 6.80, d, 8.4 | 6.85, d, 8.0 | 7.01, d, 7.6 | 6.61, d, 8.0 |
| 6′ | 7.63, dd, 8.4, 1.8 | 8.10, d, 8.4 | 8.06, d, 8.0 | 8.07, d, 8.0 | 7.61, brd, 8.4 | 7.63, dd, 8.0, 1.2 | 7.87, d, 7.6 | 7.08, dd, 7.6, 1.2 | 7.29, d, 8.4 | 7.38, d, 8.0 | 7.05, dd, 7.6, 1.2 | 6.62, brd, 8.0 |
| glc-1 | - | - | 5.28, d, 7.6 | 5.12, d, 7.6 | - | 5.24, d, 8.0 | 5.09, d, 7.6 | - | - | 5.05, d, 7.6 | 5.20, d, 8.0 | 5.09, d, 7.6 |
| glc-2 | - | - | 3.51, O | 3.27–3.79, O | - | 3.59, O | 3.25–3.81, O | - | - | 3.57, O | 3.50, O | 3.47, dd, 7.6, 7.2 |
| glc-3 | - | - | 3.28, O | 3.27–3.79, O | - | 3.35, O | 3.25–3.81, O | - | - | 3.33, O | 3.49, O | 3.39, dd, 7.2, 7.2 |
| glc-4 | - | - | 3.38, dd, 7.6, 8.0 | 3.27–3.79, O | - | 3.45, dd, 7.6, 8.0 | 3.25–3.81, O | - | - | 3.63, O | 3.37, O | 3.34, O |
| glc-5 | - | - | 3.50, O | 3.27–3.79, O | - | 3.48, O | 3.25–3.81, O | - | - | 3.55, O | 3.37, O | 3.33, O |
| glc-6 | - | - | 3.74, dd, 12.4, 6.0 | 3.27–3.79, O | - | 3.68, dd, 11.6, 5.4 | 3.25–3.81, O | - | - | 3.75, dd, 12.0, 4.8 | 3.86, dd, 11.6, 4.4 | 3.85, dd, 11.6, 5.2 |
| 3.60, dd, 12.4, 2.4 | 3.27–3.79, O | 3.55, dd, 11.6, 2.0 | 3.25–3.81, O | 3.65, dd, 12.0, 1.6 | 3.64, dd, 11.6, 1.2 | 3.67, dd, 11.6, 1.8 | ||||||
| rha-1 | - | - | - | 4.50, brs | - | - | 4.49, brs | - | - | - | - | - |
| rha-2 | - | - | - | 3.27–3.79, O | - | - | 3.25–3.81, O | - | - | - | - | - |
| rha-3 | - | - | - | 3.27–3.79, O | - | - | 3.25–3.81, O | - | - | - | - | - |
| rha-4 | - | - | - | 3.27–3.79, O | - | - | 3.25–3.81, O | - | - | - | - | - |
| rha-5 | - | - | - | 3.27–3.79, O | - | - | 3.25–3.81, O | - | - | - | - | - |
| rha-6 | - | - | - | 1.10, d, 6.0 | - | - | 1.10, d, 6.0 | - | - | - | - | - |
| No. | 1 (a) | 2 (a) | 3 (a) | 4 (a) | 5 (b) | 6 (b) | 7 (b) | 8 (a) | 9 (a) | 10 (a) | 11 (a) | 12 (a) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 158.7 | 158.0 | 158.1 | 158.1 | 163.3 | 163.2 | 164.2 | 83.8 | 82.9 | 84.9 | 83.8 | 80.9 |
| 3 | 135.8 | 137.2 | 135.2 | 135.3 | 104.5 | 104.1 | 103.8 | 72.4 | 71.5 | 73.9 | 72.4 | 44.3 |
| 4 | 179.4 | 177.3 | 179.3 | 161.1 | 182.0 | 181.6 | 182.4 | 196.9 | 197.9 | 201.1 | 196.9 | 198.7 |
| 5 | 163.2 | 162.5 | 162.6 | 162.6 | 162.5 | 162.2 | 163.0 | 164.0 | 166.9 | 165.0 | 164.0 | 165.0 |
| 6 | 100.0 | 99.3 | 99.5 | 99.8 | 99.7 | 99.6 | 98.9 | 96.2 | 96.0 | 98.4 | 96.2 | 98.0 |
| 7 | 166.1 | 165.5 | 165.9 | 165.6 | 166.4 | 166.0 | 166.5 | 167.4 | 163.3 | 167.2 | 167.4 | 166.9 |
| 8 | 94.9 | 94.5 | 94.8 | 94.7 | 94.5 | 94.4 | 94.9 | 95.1 | 95.0 | 96.8 | 95.1 | 97.0 |
| 9 | 159.2 | 158.2 | 159.0 | 159.1 | 157.9 | 157.9 | 158.1 | 163.2 | 162.6 | 166.5 | 163.2 | 164.6 |
| 10 | 105.7 | 104.5 | 105.8 | 105.4 | 104.0 | 103.8 | 116.5 | 100.5 | 100.4 | 103.0 | 100.5 | 103.8 |
| 1′ | 123.4 | 123.7 | 122.5 | 122.4 | 127.0 | 126.9 | 121.9 | 128.4 | 127.6 | 129.3 | 128.4 | 121.6 |
| 2′ | 116.1 | 130.7 | 132.1 | 132.1 | 114.6 | 114.3 | 128.9 | 114.7 | 129.5 | 130.5 | 114.8 | 116.1 |
| 3′ | 150.2 | 116.3 | 116.1 | 115.9 | 150.5 | 150.5 | 104.9 | 145.7 | 114.9 | 116.1 | 145.7 | 147.0 |
| 4′ | 145.8 | 160.5 | 161.5 | 161.1 | 149.4 | 149.3 | 162.7 | 144.9 | 157.8 | 160.1 | 144.8 | 144.5 |
| 5′ | 117.8 | 116.3 | 116.1 | 115.9 | 117.3 | 117.5 | 104.9 | 114.8 | 114.9 | 116.1 | 115.0 | 117.2 |
| 6′ | 123.2 | 130.7 | 132.1 | 132.1 | 119.8 | 119.7 | 128.9 | 119.7 | 129.5 | 130.5 | 119.8 | 119.0 |
| glc-1 | - | - | 104.0 | 104.5 | - | 104.1 | 104.5 | - | - | 101.2 | 103.9 | 101.1 |
| glc-2 | - | - | 75.5 | 75.5 | - | 75.7 | 75.8 | - | - | 75.9 | 75.7 | 74.7 |
| glc-3 | - | - | 78.1 | 77.9 | - | 78.3 | 77.8 | - | - | 78.2 | 78.1 | 77.9 |
| glc-4 | - | - | 71.1 | 73.7 | - | 71.1 | 74.0 | - | - | 71.8 | 71.3 | 70.9 |
| glc-5 | - | - | 77.8 | 76.9 | - | 77.8 | 76.8 | - | - | 78.0 | 78.0 | 78.1 |
| glc-6 | - | - | 62.6 | 68.4 | - | 62.3 | 68.7 | - | - | 62.4 | 62.4 | 62.4 |
| rha-1 | - | - | - | 102.1 | - | - | 102.1 | - | - | - | - | - |
| rha-2 | - | - | - | 71.8 | - | - | 72.1 | - | - | - | - | - |
| rha-3 | - | - | - | 72.1 | - | - | 72.3 | - | - | - | - | - |
| rha-4 | - | - | - | 71.2 | - | - | 71.5 | - | - | - | - | - |
| rha-5 | - | - | - | 69.5 | - | - | 70.1 | - | - | - | - | - |
| rha-6 | - | - | - | 17.9 | - | - | 18.0 | - | - | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-G.; Lee, H.; Jung, J.-W.; Seo, K.-H.; Lee, D.Y.; Kim, H.-G.; Ko, J.-H.; Lee, D.-S.; Baek, N.-I. Flavonoids from Chionanthus retusus (Oleaceae) Flowers and Their Protective Effects against Glutamate-Induced Cell Toxicity in HT22 Cells. Int. J. Mol. Sci. 2019, 20, 3517. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143517
Lee Y-G, Lee H, Jung J-W, Seo K-H, Lee DY, Kim H-G, Ko J-H, Lee D-S, Baek N-I. Flavonoids from Chionanthus retusus (Oleaceae) Flowers and Their Protective Effects against Glutamate-Induced Cell Toxicity in HT22 Cells. International Journal of Molecular Sciences. 2019; 20(14):3517. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143517
Chicago/Turabian StyleLee, Yeong-Geun, Hwan Lee, Jae-Woo Jung, Kyeong-Hwa Seo, Dae Young Lee, Hyoung-Geun Kim, Jung-Hwan Ko, Dong-Sung Lee, and Nam-In Baek. 2019. "Flavonoids from Chionanthus retusus (Oleaceae) Flowers and Their Protective Effects against Glutamate-Induced Cell Toxicity in HT22 Cells" International Journal of Molecular Sciences 20, no. 14: 3517. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143517