Impact of Natural Occurring ERAP1 Single Nucleotide Polymorphisms within miRNA-Binding Sites on HCMV Infection


Abstract
:1. Introduction
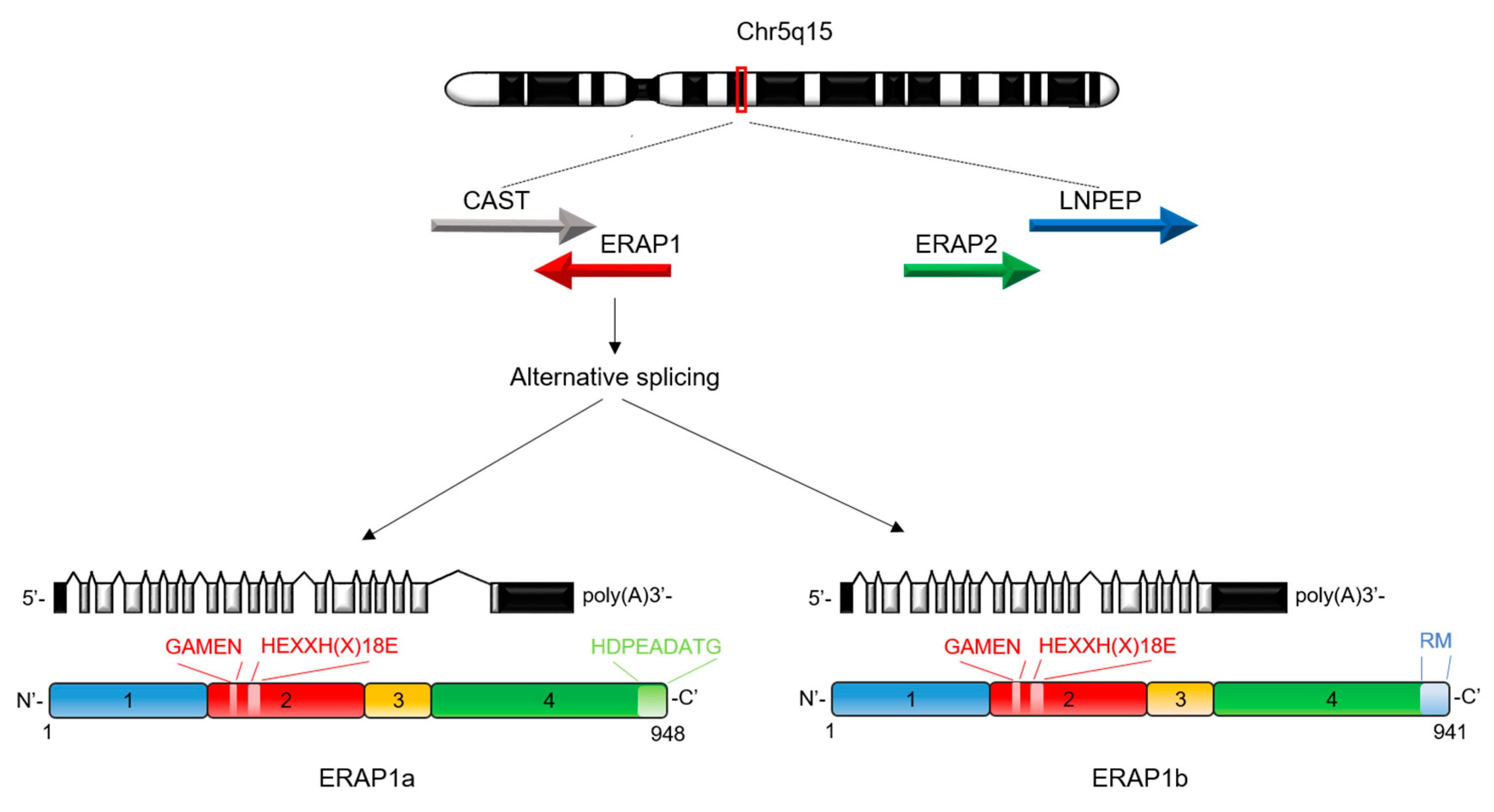
2. Biological Functions of ERAP1
3. Functional Consequences of ERAP1 Polymorphisms in Human Diseases
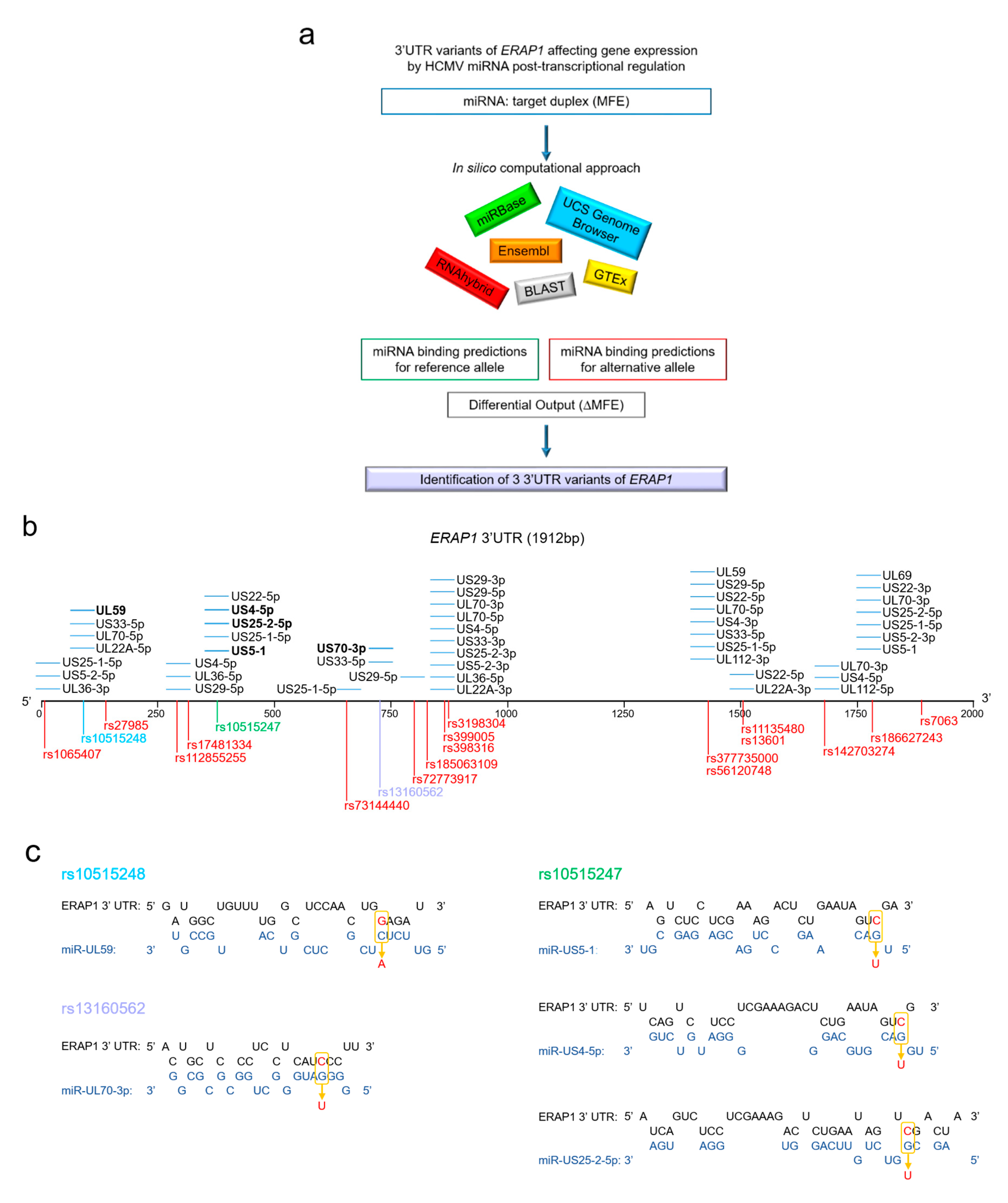
4. In Silico Prediction of miRNA-SNPs Affecting HCMV miRNA Binding Sites
5. Conclusions
Funding
Conflicts of Interest
Abbreviations
| HCMV | human cytomegalovirus |
| miRNA | microRNA |
| UL | unique long region |
| US | unique short region |
| ERAP1 | endoplasmic reticulum aminopeptidase 1 |
| SNP | single nucleotide polymorphisms |
| MHC | major histocompatibility complex |
| UTR | untranslated region |
| HLA | human leukocyte antigen |
| MAF | minor allele frequency |
| MFE | minimum free energy |
| eQTL | expression quantitative trait loci |
References
- Stempel, M.; Chan, B.; Juranic Lisnic, V.; Krmpotic, A.; Hartung, J.; Paludan, S.R.; Fullbrunn, N.; Lemmermann, N.A.; Brinkmann, M.M. The herpesviral antagonist m152 reveals differential activation of STING-dependent IRF and NF-kappaB signaling and STING’s dual role during MCMV infection. EMBO J. 2019, 38, e100983. [Google Scholar] [CrossRef] [PubMed]
- Stern-Ginossar, N.; Weisburd, B.; Michalski, A.; Le, V.T.; Hein, M.Y.; Huang, S.X.; Ma, M.; Shen, B.; Qian, S.B.; Hengel, H.; et al. Decoding human cytomegalovirus. Science 2012, 338, 1088–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varani, S.; Landini, M.P. Cytomegalovirus-induced immunopathology and its clinical consequences. Herpesviridae 2011, 2, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noriega, V.; Redmann, V.; Gardner, T.; Tortorella, D. Diverse immune evasion strategies by human cytomegalovirus. Immunol. Res. 2012, 54, 140–151. [Google Scholar] [CrossRef]
- Fruci, D.; Rota, R.; Gallo, A. The Role of HCMV and HIV-1 MicroRNAs: Processing, and Mechanisms of Action during Viral Infection. Front. Microbiol. 2017, 8, 689. [Google Scholar] [CrossRef] [Green Version]
- Van de Weijer, M.L.; Luteijn, R.D.; Wiertz, E.J. Viral immune evasion: Lessons in MHC class I antigen presentation. Semin. Immunol. 2015, 27, 125–137. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J.; Bartel, D.P.; Linsley, P.S.; Johnson, J.M. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 2005, 433, 769–773. [Google Scholar] [CrossRef]
- Grey, F.; Tirabassi, R.; Meyers, H.; Wu, G.; McWeeney, S.; Hook, L.; Nelson, J.A. A viral microRNA down-regulates multiple cell cycle genes through mRNA 5′UTRs. PLoS Pathog. 2010, 6, e1000967. [Google Scholar] [CrossRef] [Green Version]
- Grey, F.; Meyers, H.; White, E.A.; Spector, D.H.; Nelson, J. A human cytomegalovirus-encoded microRNA regulates expression of multiple viral genes involved in replication. PLoS Pathog. 2007, 3, e163. [Google Scholar] [CrossRef]
- Hook, L.; Hancock, M.; Landais, I.; Grabski, R.; Britt, W.; Nelson, J.A. Cytomegalovirus microRNAs. Curr. Opin. Virol. 2014, 7, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Hook, L.M.; Grey, F.; Grabski, R.; Tirabassi, R.; Doyle, T.; Hancock, M.; Landais, I.; Jeng, S.; McWeeney, S.; Britt, W.; et al. Cytomegalovirus miRNAs target secretory pathway genes to facilitate formation of the virion assembly compartment and reduce cytokine secretion. Cell Host Microbe. 2014, 15, 363–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Seo, D.; Kim, D.; Hong, Y.; Chang, H.; Baek, D.; Kim, V.N.; Lee, S.; Ahn, K. Temporal Landscape of MicroRNA-Mediated Host-Virus Crosstalk during Productive Human Cytomegalovirus Infection. Cell Host Microbe. 2015, 17, 838–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Kalejta, R.F.; Kerry, J.; Semmes, O.J.; O’Connor, C.M.; Khan, Z.; Garcia, B.A.; Shenk, T.; Murphy, E. BclAF1 restriction factor is neutralized by proteasomal degradation and microRNA repression during human cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2012, 109, 9575–9580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirabassi, R.; Hook, L.; Landais, I.; Grey, F.; Meyers, H.; Hewitt, H.; Nelson, J. Human cytomegalovirus US7 is regulated synergistically by two virally encoded microRNAs and by two distinct mechanisms. J. Virol. 2011, 85, 11938–11944. [Google Scholar] [CrossRef] [Green Version]
- Meshesha, M.K.; Bentwich, Z.; Solomon, S.A.; Avni, Y.S. In vivo expression of human cytomegalovirus (HCMV) microRNAs during latency. Gene 2016, 575, 101–107. [Google Scholar] [CrossRef]
- Zhang, L.; Yu, J.; Liu, Z. MicroRNAs expressed by human cytomegalovirus. Virol. J. 2020, 17, 34. [Google Scholar] [CrossRef]
- Piedade, D.; Azevedo-Pereira, J.M. The Role of microRNAs in the Pathogenesis of Herpesvirus Infection. Viruses 2016, 8, 156. [Google Scholar] [CrossRef] [Green Version]
- Pfeffer, S.; Sewer, A.; Lagos-Quintana, M.; Sheridan, R.; Sander, C.; Grasser, F.A.; van Dyk, L.F.; Ho, C.K.; Shuman, S.; Chien, M.; et al. Identification of microRNAs of the herpesvirus family. Nat. Methods 2005, 2, 269–276. [Google Scholar] [CrossRef]
- Brandetti, E.; Veneziani, I.; Melaiu, O.; Pezzolo, A.; Castellano, A.; Boldrini, R.; Ferretti, E.; Fruci, D.; Moretta, L.; Pistoia, V.; et al. MYCN is an immunosuppressive oncogene dampening the expression of ligands for NK-cell-activating receptors in human high-risk neuroblastoma. Oncoimmunology 2017, 6, e1316439. [Google Scholar] [CrossRef] [Green Version]
- Stern-Ginossar, N.; Elefant, N.; Zimmermann, A.; Wolf, D.G.; Saleh, N.; Biton, M.; Horwitz, E.; Prokocimer, Z.; Prichard, M.; Hahn, G.; et al. Host immune system gene targeting by a viral miRNA. Science 2007, 317, 376–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landais, I.; Pelton, C.; Streblow, D.; DeFilippis, V.; McWeeney, S.; Nelson, J.A. Human Cytomegalovirus miR-UL112-3p Targets TLR2 and Modulates the TLR2/IRAK1/NFkappaB Signaling Pathway. PLoS Pathog. 2015, 11, e1004881. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lee, S.; Kim, S.; Kim, D.; Ahn, J.H.; Ahn, K. Human cytomegalovirus clinical strain-specific microRNA miR-UL148D targets the human chemokine RANTES during infection. PLoS Pathog. 2012, 8, e1002577. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.; Poole, E.; Krishna, B.; Sellart, I.; Wills, M.R.; Murphy, E.; Sinclair, J. The Expression of Human Cytomegalovirus MicroRNA MiR-UL148D during Latent Infection in Primary Myeloid Cells Inhibits Activin A-triggered Secretion of IL-6. Sci. Rep. 2016, 6, 31205. [Google Scholar] [CrossRef]
- Lisboa, L.F.; Egli, A.; O’Shea, D.; Asberg, A.; Hartmann, A.; Rollag, H.; Pang, X.L.; Tyrrell, D.L.; Kumar, D.; Humar, A. Hcmv-miR-UL22A-5p: A Biomarker in Transplantation With Broad Impact on Host Gene Expression and Potential Immunological Implications. Am. J. Transplant. 2015, 15, 1893–1902. [Google Scholar] [CrossRef]
- Hancock, M.H.; Crawford, L.B.; Pham, A.H.; Mitchell, J.; Struthers, H.M.; Yurochko, A.D.; Caposio, P.; Nelson, J.A. Human Cytomegalovirus miRNAs Regulate TGF-beta to Mediate Myelosuppression while Maintaining Viral Latency in CD34(+) Hematopoietic Progenitor Cells. Cell Host Microbe 2020, 27, 104–114.e4. [Google Scholar] [CrossRef]
- Ding, M.; Wang, X.; Wang, C.; Liu, X.; Zen, K.; Wang, W.; Zhang, C.Y.; Zhang, C. Distinct expression profile of HCMV encoded miRNAs in plasma from oral lichen planus patients. J. Transl. Med. 2017, 15, 133. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Lee, S.; Shin, J.; Kim, Y.; Evnouchidou, I.; Kim, D.; Kim, Y.K.; Kim, Y.E.; Ahn, J.H.; Riddell, S.R.; et al. Human cytomegalovirus microRNA miR-US4-1 inhibits CD8(+) T cell responses by targeting the aminopeptidase ERAP1. Nat. Immunol. 2011, 12, 984–991. [Google Scholar] [CrossRef]
- Romania, P.; Cifaldi, L.; Pignoloni, B.; Starc, N.; D’Alicandro, V.; Melaiu, O.; Li Pira, G.; Giorda, E.; Carrozzo, R.; Bergvall, M.; et al. Identification of a Genetic Variation in ERAP1 Aminopeptidase that Prevents Human Cytomegalovirus miR-UL112-5p-Mediated Immunoevasion. Cell Rep. 2017, 20, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Lopez de Castro, J.A. How ERAP1 and ERAP2 Shape the Peptidomes of Disease-Associated MHC-I Proteins. Front. Immunol. 2018, 9, 2463. [Google Scholar] [CrossRef]
- Compagnone, M.; Cifaldi, L.; Fruci, D. Regulation of ERAP1 and ERAP2 genes and their disfunction in human cancer. Hum. Immunol. 2019, 80, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Andres, A.M.; Dennis, M.Y.; Kretzschmar, W.W.; Cannons, J.L.; Lee-Lin, S.Q.; Hurle, B.; Program, N.C.S.; Schwartzberg, P.L.; Williamson, S.H.; Bustamante, C.D.; et al. Balancing selection maintains a form of ERAP2 that undergoes nonsense-mediated decay and affects antigen presentation. PLoS Genet. 2010, 6, e1001157. [Google Scholar] [CrossRef] [Green Version]
- Hattori, A.; Matsumoto, K.; Mizutani, S.; Tsujimoto, M. Genomic organization of the human adipocyte-derived leucine aminopeptidase gene and its relationship to the placental leucine aminopeptidase/oxytocinase gene. J. Biochem. 2001, 130, 235–241. [Google Scholar] [CrossRef]
- Hammer, G.E.; Gonzalez, F.; James, E.; Nolla, H.; Shastri, N. In the absence of aminopeptidase ERAAP, MHC class I molecules present many unstable and highly immunogenic peptides. Nat. Immunol. 2007, 8, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Hammer, G.E.; Gonzalez, F.; Champsaur, M.; Cado, D.; Shastri, N. The aminopeptidase ERAAP shapes the peptide repertoire displayed by major histocompatibility complex class I molecules. Nat. Immunol. 2006, 7, 103–112. [Google Scholar] [CrossRef] [PubMed]
- York, I.A.; Chang, S.C.; Saric, T.; Keys, J.A.; Favreau, J.M.; Goldberg, A.L.; Rock, K.L. The ER aminopeptidase ERAP1 enhances or limits antigen presentation by trimming epitopes to 8-9 residues. Nat. Immunol. 2002, 3, 1177–1184. [Google Scholar] [CrossRef]
- Fruci, D.; Niedermann, G.; Butler, R.H.; van Endert, P.M. Efficient MHC class I-independent amino-terminal trimming of epitope precursor peptides in the endoplasmic reticulum. Immunity 2001, 15, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Reeves, E.; Edwards, C.J.; Elliott, T.; James, E. Naturally occurring ERAP1 haplotypes encode functionally distinct alleles with fine substrate specificity. J. Immunol. 2013, 191, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Kanaseki, T.; Blanchard, N.; Hammer, G.E.; Gonzalez, F.; Shastri, N. ERAAP synergizes with MHC class I molecules to make the final cut in the antigenic peptide precursors in the endoplasmic reticulum. Immunity 2006, 25, 795–806. [Google Scholar] [CrossRef] [Green Version]
- Kochan, G.; Krojer, T.; Harvey, D.; Fischer, R.; Chen, L.; Vollmar, M.; von Delft, F.; Kavanagh, K.L.; Brown, M.A.; Bowness, P.; et al. Crystal structures of the endoplasmic reticulum aminopeptidase-1 (ERAP1) reveal the molecular basis for N-terminal peptide trimming. Proc. Natl. Acad. Sci. USA 2011, 108, 7745–7750. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.; Chang, S.C.; Evnouchidou, I.; York, I.A.; Zikos, C.; Rock, K.L.; Goldberg, A.L.; Stratikos, E.; Stern, L.J. Structural basis for antigenic peptide precursor processing by the endoplasmic reticulum aminopeptidase ERAP1. Nat. Struct. Mol. Biol. 2011, 18, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Hawari, F.; Alsaaty, S.; Lawrence, M.; Combs, C.A.; Geng, W.; Rouhani, F.N.; Miskinis, D.; Levine, S.J. Identification of ARTS-1 as a novel TNFR1-binding protein that promotes TNFR1 ectodomain shedding. J. Clin. Investig. 2002, 110, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Rouhani, F.N.; Hawari, F.; Levine, S.J. Shedding of the type II IL-1 decoy receptor requires a multifunctional aminopeptidase, aminopeptidase regulator of TNF receptor type 1 shedding. J. Immunol. 2003, 171, 6814–6819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, S.; Takahashi, R.U.; Yamashita, R.; Toyoda, A.; Horii, T.; Kimura, M.; Fujiyama, A.; Nakai, K.; Tajima, S.; Matoba, R.; et al. Genome-wide analysis of DNA methylation and expression of microRNAs in breast cancer cells. Int. J. Mol. Sci. 2012, 13, 8259–8272. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Ogawa, K.; Nakamura, T.J.; Hattori, A.; Tsujimoto, M. Substrate-dependent nitric oxide synthesis by secreted endoplasmic reticulum aminopeptidase 1 in macrophages. J. Biochem. 2015, 157, 439–449. [Google Scholar] [CrossRef]
- Aldhamen, Y.A.; Seregin, S.S.; Rastall, D.P.; Aylsworth, C.F.; Pepelyayeva, Y.; Busuito, C.J.; Godbehere-Roosa, S.; Kim, S.; Amalfitano, A. Endoplasmic reticulum aminopeptidase-1 functions regulate key aspects of the innate immune response. PLoS ONE 2013, 8, e69539. [Google Scholar] [CrossRef]
- Cifaldi, L.; Romania, P.; Lorenzi, S.; Locatelli, F.; Fruci, D. Role of endoplasmic reticulum aminopeptidases in health and disease: From infection to cancer. Int. J. Mol. Sci. 2012, 13, 8338–8352. [Google Scholar] [CrossRef] [Green Version]
- Bufalieri, F.; Infante, P.; Bernardi, F.; Caimano, M.; Romania, P.; Moretti, M.; Lospinoso Severini, L.; Talbot, J.; Melaiu, O.; Tanori, M.; et al. ERAP1 promotes Hedgehog-dependent tumorigenesis by controlling USP47-mediated degradation of betaTrCP. Nat. Commun. 2019, 10, 3304. [Google Scholar] [CrossRef]
- Forloni, M.; Albini, S.; Limongi, M.Z.; Cifaldi, L.; Boldrini, R.; Nicotra, M.R.; Giannini, G.; Natali, P.G.; Giacomini, P.; Fruci, D. NF-kappaB, and not MYCN, regulates MHC class I and endoplasmic reticulum aminopeptidases in human neuroblastoma cells. Cancer Res. 2010, 70, 916–924. [Google Scholar] [CrossRef] [Green Version]
- Lorenzi, S.; Forloni, M.; Cifaldi, L.; Antonucci, C.; Citti, A.; Boldrini, R.; Pezzullo, M.; Castellano, A.; Russo, V.; van der Bruggen, P.; et al. IRF1 and NF-kB restore MHC class I-restricted tumor antigen processing and presentation to cytotoxic T cells in aggressive neuroblastoma. PLoS ONE 2012, 7, e46928. [Google Scholar] [CrossRef]
- Erre, G.L.; Piga, M.; Carru, C.; Angius, A.; Carcangiu, L.; Piras, M.; Sotgia, S.; Zinellu, A.; Mathieu, A.; Passiu, G.; et al. Global microRNA profiling of peripheral blood mononuclear cells in patients with Behcet’s disease. Clin. Exp. Rheumatol. 2015, 33, S72–S79. [Google Scholar] [PubMed]
- Puccetti, A.; Pelosi, A.; Fiore, P.F.; Patuzzo, G.; Lunardi, C.; Dolcino, M. MicroRNA Expression Profiling in Behcet’s Disease. J. Immunol. Res. 2018, 2018, 2405150. [Google Scholar] [CrossRef] [PubMed]
- Thomaidou, S.; Kracht, M.J.L.; van der Slik, A.; Laban, S.; de Koning, E.J.; Carlotti, F.; Hoeben, R.C.; Roep, B.O.; Zaldumbide, A. beta-Cell Stress Shapes CTL Immune Recognition of Preproinsulin Signal Peptide by Posttranscriptional Regulation of Endoplasmic Reticulum Aminopeptidase 1. Diabetes 2020, 69, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Judson, R.L.; Greve, T.S.; Parchem, R.J.; Blelloch, R. MicroRNA-based discovery of barriers to dedifferentiation of fibroblasts to pluripotent stem cells. Nat. Struct. Mol. Biol. 2013, 20, 1227–1235. [Google Scholar] [CrossRef] [Green Version]
- Fierabracci, A.; Milillo, A.; Locatelli, F.; Fruci, D. The putative role of endoplasmic reticulum aminopeptidases in autoimmunity: Insights from genomic-wide association studies. Autoimmun. Rev. 2012, 12, 281–288. [Google Scholar] [CrossRef]
- Babaie, F.; Hosseinzadeh, R.; Ebrazeh, M.; Seyfizadeh, N.; Aslani, S.; Salimi, S.; Hemmatzadeh, M.; Azizi, G.; Jadidi-Niaragh, F.; Mohammadi, H. The roles of ERAP1 and ERAP2 in autoimmunity and cancer immunity: New insights and perspective. Mol. Immunol. 2020, 121, 7–19. [Google Scholar] [CrossRef]
- Reeves, E.; James, E. The role of polymorphic ERAP1 in autoinflammatory disease. Biosci. Rep. 2018, 38, BSR20171503. [Google Scholar] [CrossRef]
- Reeves, E.; Elliott, T.; James, E.; Edwards, C.J. ERAP1 in the pathogenesis of ankylosing spondylitis. Immunol. Res. 2014, 60, 257–269. [Google Scholar] [CrossRef]
- Saulle, I.; Vicentini, C.; Clerici, M.; Biasin, M. An Overview on ERAP Roles in Infectious Diseases. Cells 2020, 9, 720. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Liu, N.; Zhou, Z.; Shi, L. Influence of ERAP1 and ERAP2 gene polymorphisms on disease susceptibility in different populations. Hum. Immunol. 2019, 80, 325–334. [Google Scholar] [CrossRef]
- Saunders, M.A.; Liang, H.; Li, W.H. Human polymorphism at microRNAs and microRNA target sites. Proc. Natl. Acad. Sci. USA 2007, 104, 3300–3305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manikandan, M.; Munirajan, A.K. Single nucleotide polymorphisms in microRNA binding sites of oncogenes: Implications in cancer and pharmacogenomics. OMICS 2014, 18, 142–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landi, D.; Barale, R.; Gemignani, F.; Landi, S. Prediction of the biological effect of polymorphisms within microRNA binding sites. Methods Mol. Biol. 2011, 676, 197–210. [Google Scholar]
- Sabina, S.; Vecoli, C.; Borghini, A.; Guarino, R.; Andreassi, M.G. Analysis of miRNAs Targeting 3′UTR of H2AFX Gene: A General in Silico Approach. Microrna 2015, 4, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Zee, R.Y.L.; Rivera, A.; Inostroza, Y.; Ridker, P.M.; Chasman, D.I.; Romero, J.R. Gene Variation of Endoplasmic Reticulum Aminopeptidases 1 and 2, and Risk of Blood Pressure Progression and Incident Hypertension among 17,255 Initially Healthy Women. Int. J. Genom. 2018, 2018, 2308585. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhu, J.; Zhang, W.; Chen, Y.; Zhang, K.; Popescu, L.M.; Ma, X.; Lau, W.B.; Rong, R.; Yu, X.; et al. Signature microRNA expression profile of essential hypertension and its novel link to human cytomegalovirus infection. Circulation 2011, 124, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Hattori, A.; Kitatani, K.; Matsumoto, H.; Miyazawa, S.; Rogi, T.; Tsuruoka, N.; Mizutani, S.; Natori, Y.; Tsujimoto, M. Characterization of recombinant human adipocyte-derived leucine aminopeptidase expressed in Chinese hamster ovary cells. J. Biochem. 2000, 128, 755–762. [Google Scholar] [CrossRef]
- Suppiah, V.; Moldovan, M.; Ahlenstiel, G.; Berg, T.; Weltman, M.; Abate, M.L.; Bassendine, M.; Spengler, U.; Dore, G.J.; Powell, E.; et al. IL28B is associated with response to chronic hepatitis C interferon-alpha and ribavirin therapy. Nat. Genet. 2009, 41, 1100–1104. [Google Scholar] [CrossRef]
- Bierut, L.J.; Agrawal, A.; Bucholz, K.K.; Doheny, K.F.; Laurie, C.; Pugh, E.; Fisher, S.; Fox, L.; Howells, W.; Bertelsen, S.; et al. A genome-wide association study of alcohol dependence. Proc. Natl. Acad. Sci. USA 2010, 107, 5082–5087. [Google Scholar] [CrossRef] [Green Version]
- Biernacka, J.M.; Geske, J.R.; Schneekloth, T.D.; Frye, M.A.; Cunningham, J.M.; Choi, D.S.; Tapp, C.L.; Lewis, B.R.; Drews, M.S.; Pietrzak, T.L.; et al. Replication of genome wide association studies of alcohol dependence: Support for association with variation in ADH1C. PLoS ONE 2013, 8, e58798. [Google Scholar] [CrossRef] [Green Version]
- Greenawalt, D.M.; Sieberts, S.K.; Cornelis, M.C.; Girman, C.J.; Zhong, H.; Yang, X.; Guinney, J.; Qi, L.; Hu, F.B. Integrating genetic association, genetics of gene expression, and single nucleotide polymorphism set analysis to identify susceptibility Loci for type 2 diabetes mellitus. Am. J. Epidemiol. 2012, 176, 423–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhernakova, D.V.; de Klerk, E.; Westra, H.J.; Mastrokolias, A.; Amini, S.; Ariyurek, Y.; Jansen, R.; Penninx, B.W.; Hottenga, J.J.; Willemsen, G.; et al. DeepSAGE reveals genetic variants associated with alternative polyadenylation and expression of coding and non-coding transcripts. PLoS Genet. 2013, 9, e1003594. [Google Scholar] [CrossRef]
- Sezgin, E.; An, P.; Winkler, C.A. Host Genetics of Cytomegalovirus Pathogenesis. Front. Genet. 2019, 10, 616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moens, U. Silencing viral microRNA as a novel antiviral therapy? J. Biomed. Biotechnol. 2009, 2009, 419539. [Google Scholar] [CrossRef]
- Pfeffer, S.; Zavolan, M.; Grasser, F.A.; Chien, M.; Russo, J.J.; Ju, J.; John, B.; Enright, A.J.; Marks, D.; Sander, C.; et al. Identification of virus-encoded microRNAs. Science 2004, 304, 734–736. [Google Scholar] [CrossRef]
- Mishra, R.; Kumar, A.; Ingle, H.; Kumar, H. The Interplay Between Viral-Derived miRNAs and Host Immunity During Infection. Front. Immunol. 2019, 10, 3079. [Google Scholar] [CrossRef] [Green Version]
- Tahamtan, A.; Inchley, C.S.; Marzban, M.; Tavakoli-Yaraki, M.; Teymoori-Rad, M.; Nakstad, B.; Salimi, V. The role of microRNAs in respiratory viral infection: Friend or foe? Rev. Med. Virol. 2016, 26, 389–407. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, N.; Zhou, Z.; Liang, H.; Pan, C.; Zhu, D.; Liu, F.; Zhang, C.Y.; Zhang, Y.; Zen, K. Circulating human cytomegalovirus-encoded HCMV-miR-US4-1 as an indicator for predicting the efficacy of IFNalpha treatment in chronic hepatitis B patients. Sci. Rep. 2016, 6, 23007. [Google Scholar] [CrossRef]
- Cipollini, M.; Landi, S.; Gemignani, F. MicroRNA binding site polymorphisms as biomarkers in cancer management and research. Pharmgenomics Pers. Med. 2014, 7, 173–191. [Google Scholar]
- Hu, Z.; Bruno, A.E. The Influence of 3′UTRs on MicroRNA Function Inferred from Human SNP Data. Comp. Funct. Genom. 2011, 2011, 910769. [Google Scholar] [CrossRef] [Green Version]
- Nakken, S.; Alseth, I.; Rognes, T. Computational prediction of the effects of non-synonymous single nucleotide polymorphisms in human DNA repair genes. Neuroscience 2007, 145, 1273–1279. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.R.; Marnellos, G.; Kammerer, S.; Hoyal, C.R.; Shi, M.M.; Cantor, C.R.; Braun, A. Large-scale validation of single nucleotide polymorphisms in gene regions. Genome. Res. 2004, 14, 1664–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| SNP | SNP Position | MAF * | HCMV miRNA | Seed Sequence ** | MFE # (Kcal/mol) | ΔMFE ##† (Kcal/mol) |
|---|---|---|---|---|---|---|
| rs1065407 (T/G) | 5:96776379 | 0.174 | UL22A-3p | NO | −18.1 | +4.7 |
| UL22A-5p | NO | −14.8 | +1.8 | |||
| UL70-3p | NO | −23.2 | −1 | |||
| US5-1 | NO | −21.6 | +4.7 | |||
| US5-2-3p | NO | −15 | +10.8 | |||
| rs10515248 (C/T) | 5:96776301 | 0.06 | UL69 | NO | −22.6 | 3 |
| UL112-3p | YES | −18.6 | +1.1 | |||
| US22-3p | NO | −12.4 | +3.6 | |||
| US25-2-3p | YES | −24.1 | +5.4 | |||
| US29-3p | YES | −16.6 | +5.4 | |||
| US33-3p | YES | −20.4 | +1.1 | |||
| rs27985 (A/G) | 5:96776250 | 0.006 | UL59 | YES | −14.3 | −1.8 |
| UL112-5p | NO | −12.8 | +5.5 | |||
| US5-2-3p | NO | −13.7 | −1.9 | |||
| US5-2-5p | NO | −10.1 | +5.6 | |||
| US22-3p | NO | −17.6 | −1.7 | |||
| US25-1-3p | NO | −15.7 | +10.6 | |||
| US25-2-5p | NO | −21.8 | −1.4 | |||
| rs112855255 (T/G) | 5:96776101 | 0.06 | UL36-3p | NO | −17.9 | +2.1 |
| UL69 | YES | −16.7 | −2.6 | |||
| US5-1 | NO | −15.3 | 1 | |||
| US5-2-3p | NO | −15.1 | +10.8 | |||
| US22-3p | NO | −14.7 | −1.3 | |||
| US33-5p | NO | −19.6 | −3.9 | |||
| rs17481334 (A/G) | 5:96776075 | 0.06 | UL22A-3p | NO | −11.4 | −2.1 |
| UL36-3p | NO | −11.4 | −2.1 | |||
| UL70-3p | NO | −12.7 | −1.4 | |||
| UL112-3p | NO | −13.5 | −1.2 | |||
| US5-1 | NO | −10.9 | +7.3 | |||
| US29-5p | NO | −12.9 | +2.7 | |||
| US33-5p | NO | −16.3 | −1.8 | |||
| rs10515247 (G/A) | 5:96776022 | 0.06 | UL22A-3p | NO | −12.2 | −1.8 |
| UL22A-5p | YES | −11.1 | −2.3 | |||
| UL69 | YES | −12 | +1.7 | |||
| UL70-5p | NO | −13.4 | +1.9 | |||
| US5-2-3p | NO | −16.9 | +4.9 | |||
| US22-3p | NO | −13.5 | 2 | |||
| US25-1-3p | YES | −14 | +1.2 | |||
| US29-5p | YES | −14.2 | −1.1 | |||
| rs73144440 (G/A) | 5:96775743 | 0.008 | UL22A-3p | NO | −18.6 | +2.6 |
| US4-5p | YES | −27 | +1.6 | |||
| US5-2-5p | NO | −11.6 | −1 | |||
| US22-3p | NO | −20.1 | −1.2 | |||
| US25-1-3p | NO | −22.3 | −1.8 | |||
| US25-2-5p | NO | −21.8 | −4.9 | |||
| US33-5p | NO | −23.4 | +2.7 | |||
| rs13160562 (G/A) | 5:96775667 | 0.153 | US4-5p | YES | −19.4 | −1.4 |
| US25-2-5p | NO | −21.8 | −2.9 | |||
| US29-5p | YES | −16.5 | +1.1 | |||
| rs72773917 (A/T) | 5:96775598 | 0.06 | UL59 | NO | −10.8 | +7.2 |
| US5-2-3p | NO | −17.4 | 1 | |||
| rs185063109 (T/C) | 5:96775565 | 0.005 | US5-2-3p | NO | −15.2 | +7.4 |
| US25-2-3p | YES | −11.9 | −3.3 | |||
| rs398316 (C/G) | 5:96775535 | 0.065 | UL69 | YES | −12.7 | −3.2 |
| UL70-5p | NO | −16.4 | +3.1 | |||
| UL112-3p | YES | −13.8 | −3.9 | |||
| US4-3p | YES | −14.7 | +3.9 | |||
| US5-1 | YES | −17 | +3.2 | |||
| US5-2-3p | NO | −17.2 | −1.3 | |||
| US33-5p | YES | −14.1 | +1.7 | |||
| rs399005 (T/A) | 5:96775536 | 0.064 | UL59 | YES | −16.4 | +3.8 |
| UL69 | NO | −12.7 | +1.3 | |||
| US5-2-5p | YES | −14.1 | −1 | |||
| US29-3p | NO | −18.3 | −2.8 | |||
| US33-3p | NO | −14.2 | −3.1 | |||
| US33-5p | YES | −14.1 | +1.1 | |||
| rs3198304 (A/G) | 5:96775524 | 0.153 | UL22A-5p | NO | −11 | +6.6 |
| US5-2-3p | YES | −17.2 | −2 | |||
| rs3198304 (A/G) | 5:96775524 | 0.153 | US5-2-5p | NO | −10.1 | +5.4 |
| US25-2-3p | NO | −16.4 | −2.6 | |||
| US25-2-5p | NO | −21.1 | −2.5 | |||
| rs3198304 (A/T) | 5:96775524 | 0.153 | US5-2-5p | NO | −10.1 | +5.4 |
| rs56120748 (C/G) | 5:96774918 | 0.06 | UL70-5p | NO | −16.3 | +4.6 |
| US4-3p | NO | −13.4 | +2.7 | |||
| US29-3p | NO | −16 | +8.2 | |||
| rs56120748 (C/T) | 5:96774918 | 0.06 | UL70-5p | NO | −16.3 | +5.2 |
| UL112-5p | NO | −13.1 | +4.3 | |||
| US29-3p | NO | −16 | +8.2 | |||
| rs377735000 (G/A) | 5:96774915 | 0.002 | UL70-3p | NO | −18.6 | +15.1 |
| UL70-5p | NO | −14.5 | −1.8 | |||
| US4-3p | NO | −13.4 | +9.1 | |||
| US5-2-3p | NO | −15.2 | +2.4 | |||
| US25-1-5p | NO | −15.5 | +1.4 | |||
| US29-3p | YES | −16 | +8.2 | |||
| US29-5p | NO | −15.6 | +4.7 | |||
| US33-5p | YES | −14.7 | 3 | |||
| rs377735000 (G/C) | 5:96774915 | 0.002 | UL59 | NO | −13.3 | −2.7 |
| UL70-3p | NO | −18.6 | +14.5 | |||
| US4-3p | NO | −13.4 | +9.1 | |||
| US5-2-3p | NO | −15.2 | +7.4 | |||
| US29-3p | YES | −16 | +8.2 | |||
| US29-5p | NO | −15.6 | +5.3 | |||
| rs377735000 (G/T) | 5:96774915 | 0.002 | UL70-3p | NO | −18.6 | +14.2 |
| US4-3p | NO | −13.4 | +9.1 | |||
| US5-2-3p | NO | −15.2 | +7.4 | |||
| US29-3p | YES | −16 | +8.2 | |||
| rs13601 (G/A) | 5:96774878 | 0.06 | UL70-3p | YES | −18.6 | 3 |
| US4-5p | NO | −22 | 3 | |||
| US5-2-3p | NO | −12.1 | 3 | |||
| US25-2-5p | NO | −17.5 | 3 | |||
| rs11135480 (T/G) | 5:96774878 | 0.06 | UL22A-5p | NO | −10.6 | −1.7 |
| rs186627243 (C/T) | 5:96774610 | 0.002 | UL22A-3p | NO | −14.9 | 3 |
| UL59 | NO | −15.7 | +5.4 | |||
| US5-2-5p | NO | −15.4 | 3 | |||
| US25-1-3p | NO | −15.9 | 2 | |||
| US29-5p | NO | −20.8 | +16.6 | |||
| US33-5p | NO | −16.3 | +1.3 |
| SNP | SNP Position | MAF * | HCMV miRNA | Seed Sequence ** | MFE # (Kcal/mol) |
|---|---|---|---|---|---|
| rs1065407 (T/G) | 5:96776379 | 0.174 | UL36-3p | NO | −10.8 |
| US5-2-5p | NO | −10.9 | |||
| US25-1-5p | NO | −15.8 | |||
| rs10515248 (C/T) | 5:96776301 | 0.06 | UL22A-5p | NO | −12.4 |
| UL59 | YES | −19.3 | |||
| UL70-5p | NO | −19 | |||
| rs112855255 (T/G) | 5:96776101 | 0.06 | UL36-5p | NO | −14.5 |
| US4-5p | YES | −16.9 | |||
| US29-5p | YES | −20.1 | |||
| rs10515247 (G/A) | 5:96776022 | 0.06 | US4-5p | YES | −17.7 |
| US5-1 | YES | −14.3 | |||
| US22-5p | NO | −14.3 | |||
| US25-1-5p | NO | −18.5 | |||
| US25-2-5p | YES | −21.4 | |||
| US33-5p | NO | −16.2 | |||
| rs73144440 (G/A) | 5:96775743 | 0.008 | US25-1-5p | NO | −25.2 |
| rs13160562 (G/A) | 5:96775667 | 0.153 | UL70-3p | YES | −26.4 |
| US33-5p | NO | −18.6 | |||
| rs72773917 (A/T) | 5:96775598 | 0.06 | US29-5p | NO | −15.8 |
| rs398316 (C/G) | 5:96775535 | 0.065 | UL70-3p | NO | −23.8 |
| US29-3p | NO | −18.3 | |||
| US29-5p | NO | −20.6 | |||
| US33-3p | NO | −14.2 | |||
| rs399005 (T/A) | 5:96775536 | 0.064 | UL70-3p | NO | −23.8 |
| UL70-5p | NO | −16.4 | |||
| US4-5p | NO | −25.2 | |||
| US5-2-3p | NO | −17.2 | |||
| US25-2-3p | NO | −16.4 | |||
| rs3198304 (A/G) | 5:96775524 | 0.153 | UL36-5p | NO | −14.8 |
| UL70-3p | YES | −23.8 | |||
| US4-5p | NO | −25.2 | |||
| US33-3p | NO | −14.2 | |||
| rs3198304 (A/T) | 5:96775524 | 0.153 | UL22A-3p | NO | −17.5 |
| UL36-5p | NO | −14.8 | |||
| UL70-3p | YES | −23.8 | |||
| US25-2-3p | NO | −16.4 | |||
| rs56120748 (C/G) | 5:96774918 | 0.06 | US25-1-5p | NO | −15.5 |
| US29-5p | NO | −15.6 | |||
| US33-5p | NO | −14.7 | |||
| rs56120748 (C/T) | 5:96774918 | 0.06 | UL112-3p | NO | −13.7 |
| US4-3p | NO | −13.4 | |||
| US25-1-5p | NO | −15.5 | |||
| US29-5p | NO | −15.6 | |||
| US33-5p | NO | −14.7 | |||
| rs377735000 (G/A) | 5:96774915 | 0.002 | UL59 | NO | −13.3 |
| US22-5p | YES | −15.6 | |||
| rs377735000 (G/C) | 5:96774915 | 0.002 | UL70-5p | NO | −14.5 |
| US22-5p | YES | −15.6 | |||
| US25-1-5p | NO | −15.5 | |||
| US33-5p | YES | −14.7 | |||
| rs377735000 (G/T) | 5:96774915 | 0.002 | UL59 | NO | −13.3 |
| UL70-5p | NO | −14.5 | |||
| US22-5p | YES | −15.6 | |||
| US33-5p | YES | −14.7 | |||
| rs13601 (G/A) | 5:96774892 | 0.06 | US22-5p | NO | −19.1 |
| rs11135480 (T/G) | 5:96774878 | 0.006 | UL22A-3p | NO | −19.3 |
| rs142703274 (A/T) | 5:96774718 | 0.001 | UL70-3p | NO | −21.2 |
| UL112-5p | NO | −11.7 | |||
| US4-5p | NO | −17.4 | |||
| rs186627243 (C/T) | 5:96774610 | 0.002 | UL69 | NO | −20.9 |
| UL70-3p | NO | −16.5 | |||
| US5-1 | NO | −16.6 | |||
| US5-2-3p | NO | −20 | |||
| US22-3p | NO | −15.5 | |||
| US25-1-5p | NO | −20.1 | |||
| US25-2-5p | NO | -22.2 |
| SNP | SNP Position | |ΔMFE|tot * |
|---|---|---|
| rs377735000 (G/A) | 5:96774915 | 51.1 |
| rs377735000 (G/C) | 5:96774915 | 47.2 |
| rs377735000 (G/T) | 5:96774915 | 38.9 |
| rs186627243 (C/T) | 5:96774610 | 31.3 |
| rs27985 (A/G) | 5:96776250 | 28.5 |
| rs1065407 (T/G) | 5:96776379 | 23.0 |
| rs112855255 (T/G) | 5:96776101 | 21.7 |
| rs398316 (C/G) | 5:96775535 | 20.3 |
| rs10515248 (C/T) | 5:96776301 | 19.6 |
| rs3198304 (A/G) | 5:96775524 | 19.1 |
| rs17481334 (A/G) | 5:96776075 | 18.6 |
| rs56120748 (C/T) | 5:96774918 | 17.7 |
| rs10515247 (G/A) | 5:96776022 | 16.9 |
| rs73144440 (G/A) | 5:96775743 | 15.8 |
| rs56120748 (C/G) | 5:96774918 | 15.5 |
| rs399005 (T/A) | 5:96775536 | 13.1 |
| rs13601 (G/A) | 5:96774892 | 12.0 |
| rs185063109 (T/C) | 5:96775565 | 10.7 |
| rs72773917 (A/T) | 5:96775598 | 8.2 |
| rs3198304 (A/T) | 5:96775524 | 5.4 |
| rs13160562 (G/A) | 5:96775667 | 5.4 |
| rs11135480 (T/G) | 5:96774878 | 1.8 |
| SNP | Muscle Tissue | Lung Tissue | Whole Blood | Fibroblasts |
|---|---|---|---|---|
| rs1065407 | 1.3 × 10−62 | 2.3 × 10−19 | 2.9 × 10−59 | 5.2 × 10−8 |
| rs7063 | 5.6 × 10−76 | 5.4 × 10−32 | 2.6 × 10−81 | 2.6 × 10−13 |
| rs56120748 | 2.2 × 10−20 | 6.0 × 10−29 | 3.1 × 10−10 | 3.8 × 10−19 |
| rs72773917 | 4.0 × 10−20 | 1.4 × 10−29 | 1.6 × 10−10 | 1.3 × 10−19 |
| rs13601 | 1.3 × 10−20 | 1.4 × 10−29 | 2.6 × 10−10 | 5.9 × 10−20 |
| rs11135480 | 4.0 × 10−20 | 1.4 × 10−29 | 1.6 × 10−10 | 1.3 × 10−19 |
| rs112855255 | 4.0 × 10−20 | 1.4 × 10−29 | 1.6 × 10−10 | 1.3 × 10−19 |
| rs10515247 | 4.6 × 10−20 | 1.3 × 10−29 | 3.5 × 10−10 | 1.5 × 10−19 |
| rs10515248 | 4.6 × 10−20 | 1.3 × 10−29 | 3.5 × 10−10 | 1.5 × 10−19 |
| rs13160562 | 9.8 × 10−75 | 1.4 × 10−31 | 3.9 × 10−81 | 2.9 × 10−13 |
| rs17481334 | 4.0 × 10−20 | 1.4 × 10−29 | 1.6 × 10−10 | 1.3 × 10−19 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melaiu, O.; D’Amico, S.; Tempora, P.; Lucarini, V.; Fruci, D. Impact of Natural Occurring ERAP1 Single Nucleotide Polymorphisms within miRNA-Binding Sites on HCMV Infection. Int. J. Mol. Sci. 2020, 21, 5861. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165861
Melaiu O, D’Amico S, Tempora P, Lucarini V, Fruci D. Impact of Natural Occurring ERAP1 Single Nucleotide Polymorphisms within miRNA-Binding Sites on HCMV Infection. International Journal of Molecular Sciences. 2020; 21(16):5861. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165861
Chicago/Turabian StyleMelaiu, Ombretta, Silvia D’Amico, Patrizia Tempora, Valeria Lucarini, and Doriana Fruci. 2020. "Impact of Natural Occurring ERAP1 Single Nucleotide Polymorphisms within miRNA-Binding Sites on HCMV Infection" International Journal of Molecular Sciences 21, no. 16: 5861. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165861