Mapping Calcium Dynamics in the Heart of Zebrafish Embryos with Ratiometric Genetically Encoded Calcium Indicators

 , , , and
, , , and
Abstract
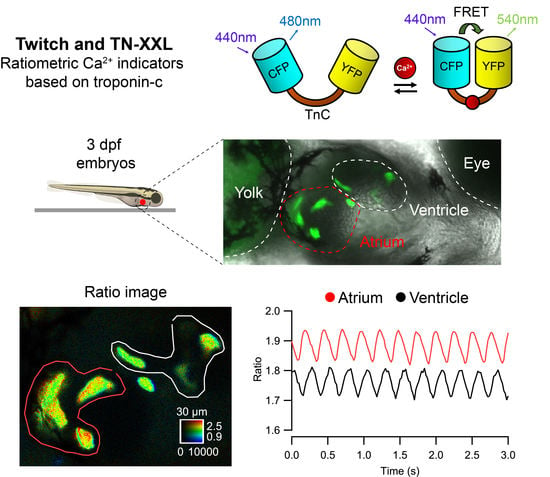
:
1. Introduction
2. Results
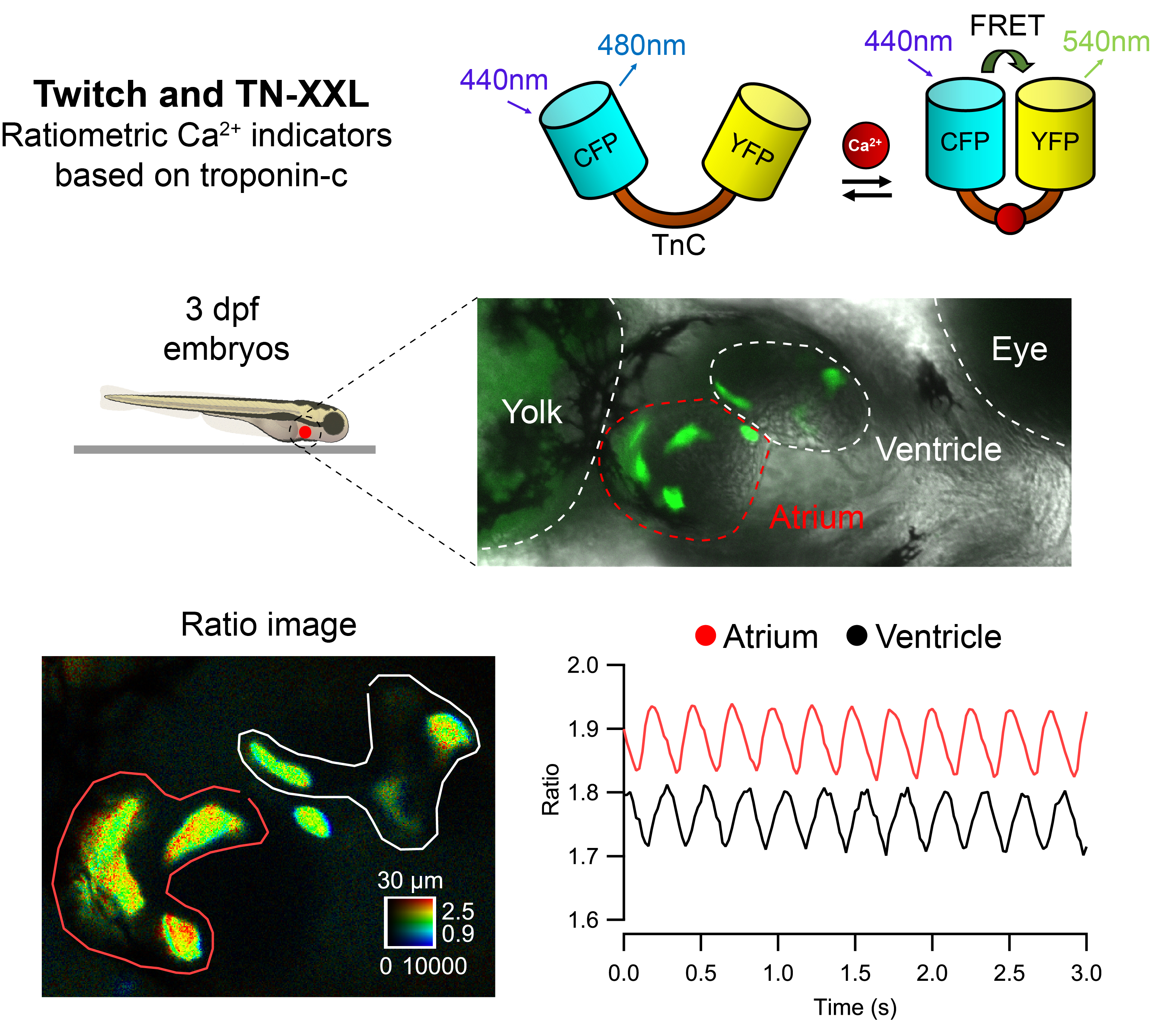
2.1. Transient Expression of TN-XXL and Twitch Ca2+ Biosensors in the Heart of Zebrafish Embryos
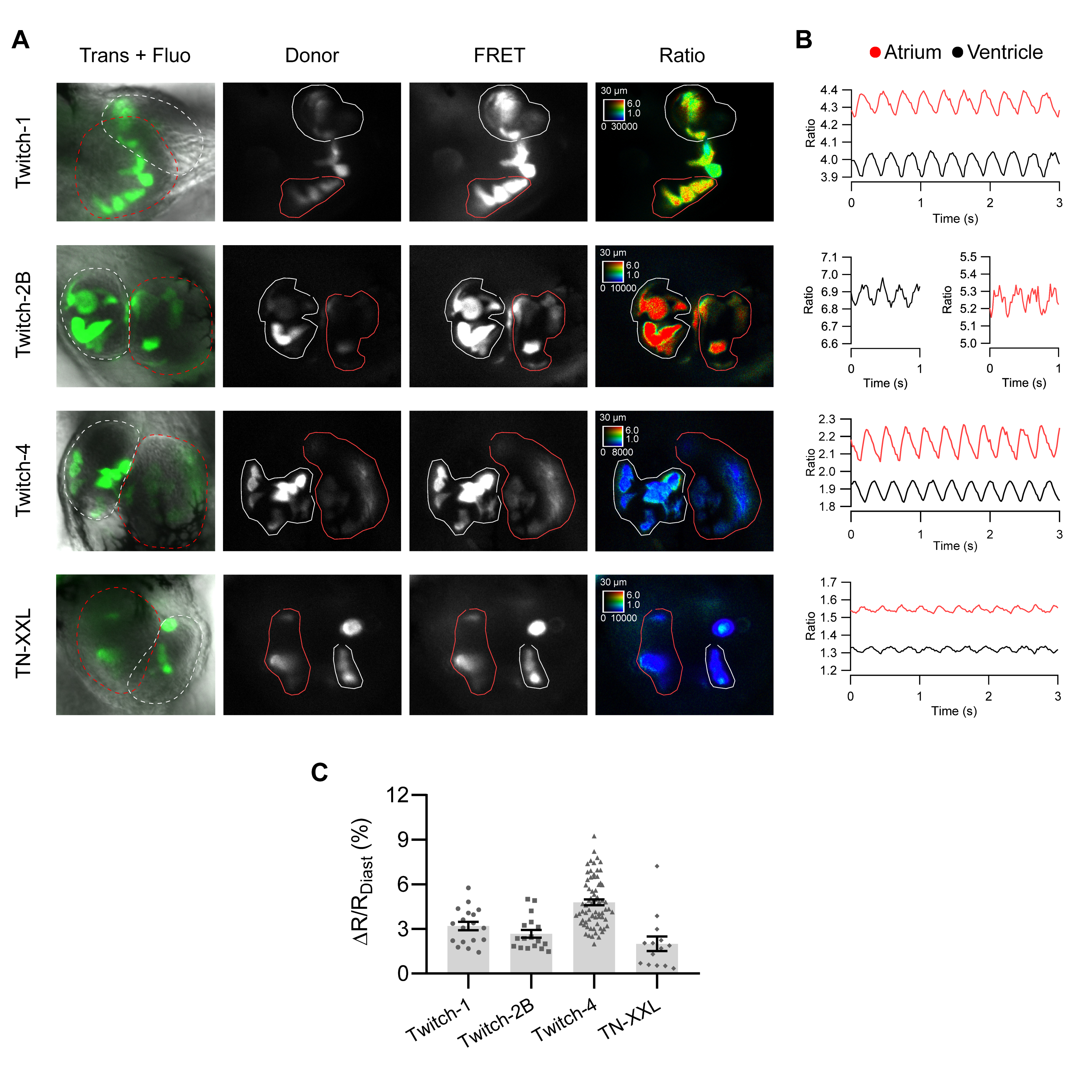
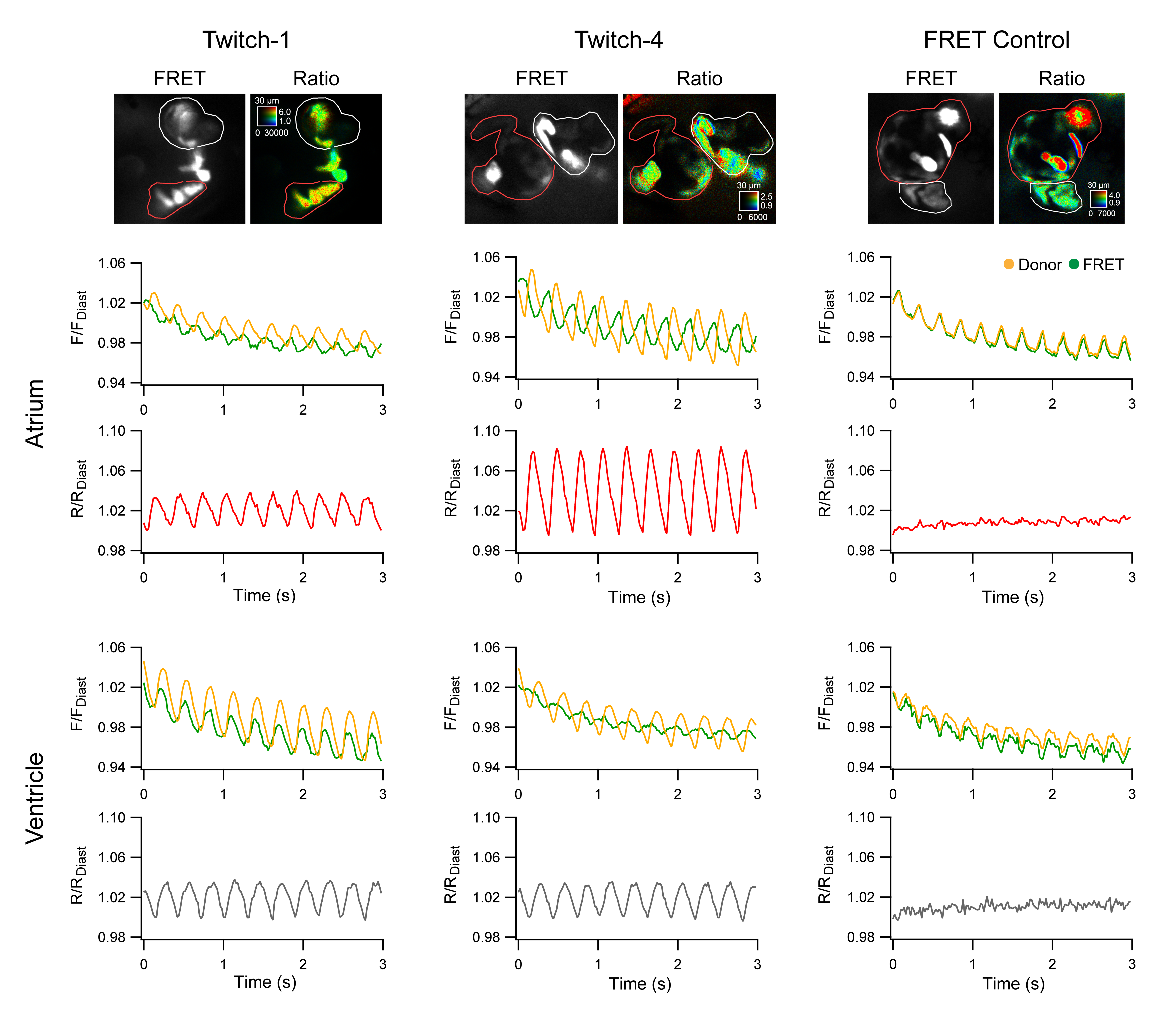
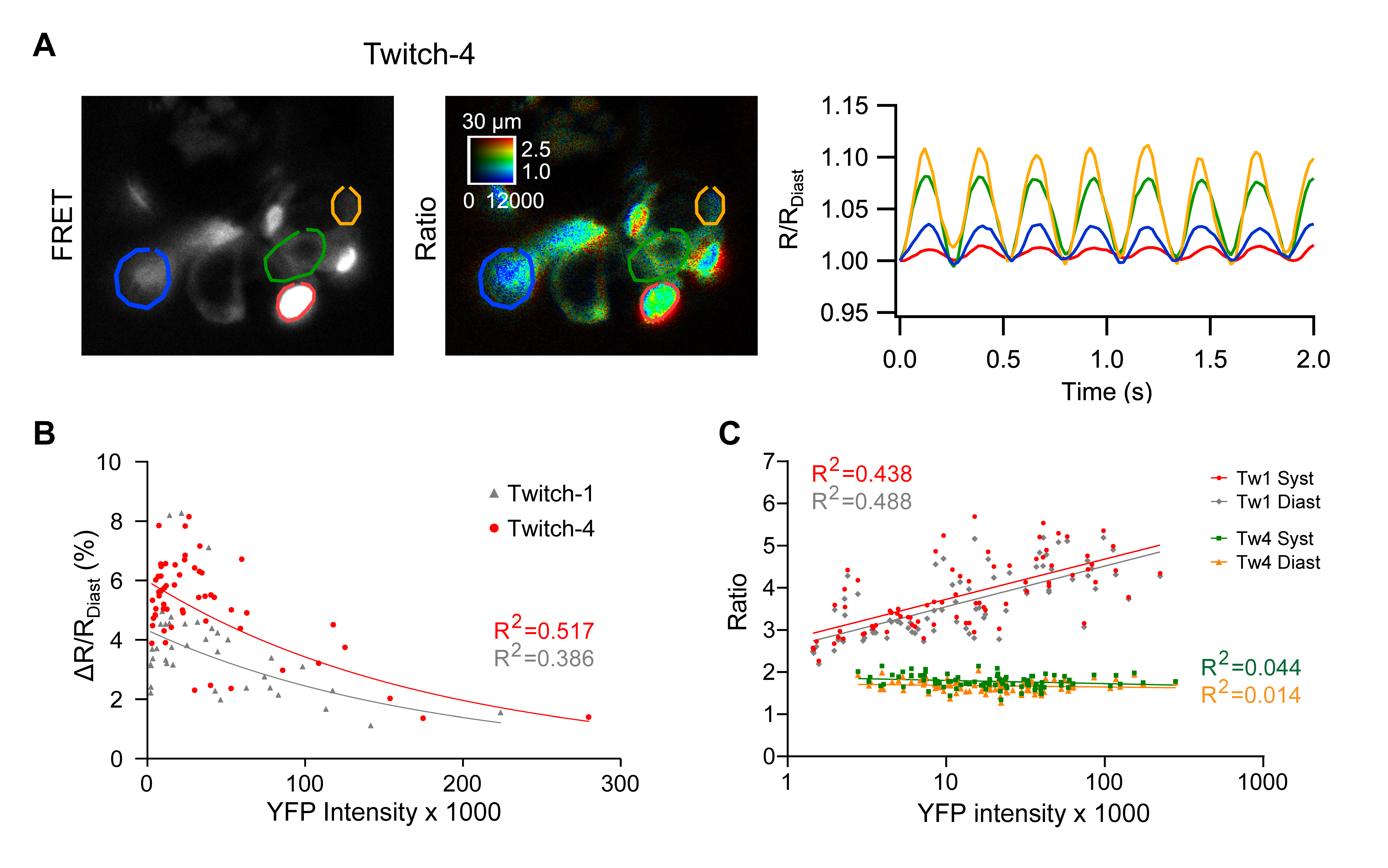
2.2. Ratiometric Imaging Reports Ca2+ Changes and Corrects for Motion Artifacts
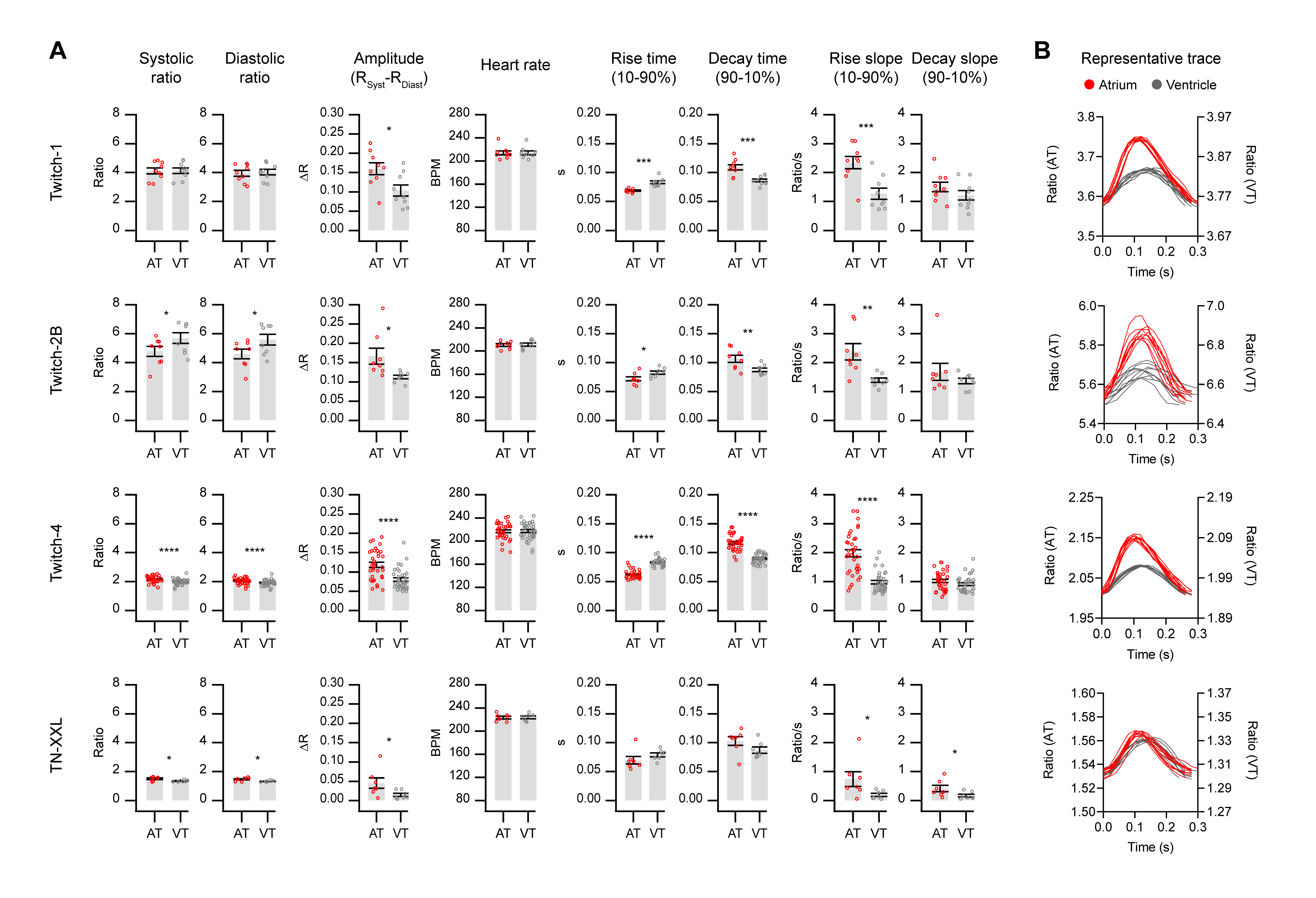
2.3. Basal Cardiac Ca2+ Kinetics Obtained with Each Biosensor
2.4. Influence of GECI Overexpression on Ca2+ Transients and on Functional Parameters of the Heart
2.5. Effect of an L-Type Ca2+ Channel Blocker on Ca2+ Transients in the Zebrafish Heart
2.6. Effect of a β-Adrenergic Antagonist on Ca2+ Transients in the Zebrafish Heart
3. Discussion
4. Materials and Methods
4.1. Constructs, Plasmids, and mRNA Synthesis
4.2. Collection and Maintenance of Zebrafish Embryos
4.3. Microinjection of Eggs
4.4. Mounting of Embryos for Microscopy
4.5. Confocal Microscopy
4.6. Ratiometric Fluorescence Imaging of TN-XXL, Twitch-1, Twitch-2b and Twitch-4
4.7. Image Processing and Data Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| bpm | beats per minute |
| cpCitrine174 | citrine fluorescent protein circularly permuted at amino acid 174 |
| DMSO | dimethyl sulfoxide |
| dpf | days post-fertilization |
| ECFP | enhanced cyan fluorescent protein |
| FP | fluorescent protein |
| FRET | Fluorescence/Förster resonance energy transfer |
| GECI | genetically-encoded Ca2+ indicator |
| GFP | green fluorescent protein |
| hpf | hours post-fertilization |
| HR | heart rate |
| LTCC | L-type Ca2+ channel |
| NCX | Na-Ca exchanger |
| PAB | para-amino blebbistatin |
| ROI | region of interest |
| SNR | signal-to-noise ratio |
| TTCC | T-type Ca2+ channel |
References
- Brunello, L.; Slabaugh, J.L.; Radwanski, P.B.; Ho, H.T.; Belevych, A.E.; Lou, Q.; Chen, H.; Napolitano, C.; Lodola, F.; Priori, S.G.; et al. Decreased RyR2 refractoriness determines myocardial synchronization of aberrant Ca2+ release in a genetic model of arrhythmia. Proc. Natl. Acad. Sci. USA 2013, 110, 10312–10317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landstrom, A.P.; Dobrev, D.; Wehrens, X.H.T. Calcium signaling and cardiac arrhythmias. Circ. Res. 2017, 120, 1969–1993. [Google Scholar] [CrossRef] [PubMed]
- Nemec, J.; Kim, J.J.; Salama, G. The link between abnormal calcium handling and electrical instability in acquired long QT syndrome—Does calcium precipitate arrhythmic storms? Prog. Biophys. Mol. Biol. 2016, 120, 210–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macrae, C.A. Cardiac Arrhythmia: In vivo screening in the zebrafish to overcome complexity in drug discovery. Expert Opin. Drug Discov. 2010, 5, 619–632. [Google Scholar] [CrossRef] [Green Version]
- Alday, A.; Alonso, H.; Gallego, M.; Urrutia, J.; Letamendia, A.; Callol, C.; Casis, O. Ionic channels underlying the ventricular action potential in zebrafish embryo. Pharmacol. Res. 2014, 84, 26–31. [Google Scholar] [CrossRef]
- Arnaout, R.; Ferrer, T.; Huisken, J.; Spitzer, K.; Stainier, D.Y.; Tristani-Firouzi, M.; Chi, N.C. Zebrafish model for human long QT syndrome. Proc. Natl. Acad. Sci. USA 2007, 104, 11316–11321. [Google Scholar] [CrossRef] [Green Version]
- Haverinen, J.; Hassinen, M.; Dash, S.N.; Vornanen, M. Expression of calcium channel transcripts in the zebrafish heart: Dominance of T-type channels. J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, P.; Ireland, J.; Grunow, B. Fish, the better model in human heart research? Zebrafish Heart aggregates as a 3D spontaneously cardiomyogenic in vitro model system. Prog. Biophys. Mol. Biol. 2018, 138, 132–141. [Google Scholar] [CrossRef]
- Milan, D.J.; Jones, I.L.; Ellinor, P.T.; MacRae, C.A. In vivo recording of adult zebrafish electrocardiogram and assessment of drug-induced QT prolongation. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H269. [Google Scholar] [CrossRef]
- Nemtsas, P.; Wettwer, E.; Christ, T.; Weidinger, G.; Ravens, U. Adult zebrafish heart as a model for human heart? An electrophysiological study. J. Mol. Cell. Cardiol. 2010, 48, 161–171. [Google Scholar] [CrossRef]
- Van Opbergen, C.J.M.; van der Voorn, S.M.; Vos, M.A.; de Boer, T.P.; van Veen, T.A.B. Cardiac Ca(2+) signalling in zebrafish: Translation of findings to man. Prog. Biophys. Mol. Biol. 2018, 138, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R.; Samsa, L.A.; Qian, L.; Liu, J. Advances in the Study of Heart Development and Disease Using Zebrafish. J. Cardiovasc. Dev. Dis. 2016, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Burns, C.G.; Milan, D.J.; Grande, E.J.; Rottbauer, W.; MacRae, C.A.; Fishman, M.C. High-throughput assay for small molecules that modulate zebrafish embryonic heart rate. Nat. Chem. Biol. 2005, 1, 263–264. [Google Scholar] [CrossRef] [PubMed]
- Cornet, C.; Calzolari, S.; Minana-Prieto, R.; Dyballa, S.; van Doornmalen, E.; Rutjes, H.; Savy, T.; D’Amico, D.; Terriente, J. ZeGlobalTox: An Innovative Approach to Address Organ Drug Toxicity Using Zebrafish. Int. J. Mol. Sci. 2017, 18, 864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letamendia, A.; Quevedo, C.; Ibarbia, I.; Virto, J.M.; Holgado, O.; Diez, M.; Izpisua Belmonte, J.C.; Callol-Massot, C. Development and validation of an automated high-throughput system for zebrafish in vivo screenings. PLoS ONE 2012, 7, e36690. [Google Scholar] [CrossRef]
- Bovo, E.; Dvornikov, A.V.; Mazurek, S.R.; de Tombe, P.P.; Zima, A.V. Mechanisms of Ca(2)+ handling in zebrafish ventricular myocytes. Pflug. Arch. Eur. J. Physiol. 2013, 465, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.C.; Llach, A.; Sheng, X.Y.; Hove-Madsen, L.; Tibbits, G.F. Calcium handling in zebrafish ventricular myocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R56–R66. [Google Scholar] [CrossRef] [Green Version]
- Akerboom, J.; Chen, T.W.; Wardill, T.J.; Tian, L.; Marvin, J.S.; Mutlu, S.; Calderon, N.C.; Esposti, F.; Borghuis, B.G.; Sun, X.R.; et al. Optimization of a GCaMP calcium indicator for neural activity imaging. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 13819–13840. [Google Scholar] [CrossRef]
- Chen, T.W.; Wardill, T.J.; Sun, Y.; Pulver, S.R.; Renninger, S.L.; Baohan, A.; Schreiter, E.R.; Kerr, R.A.; Orger, M.B.; Jayaraman, V.; et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 2013, 499, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Nakai, J.; Ohkura, M.; Imoto, K. A high signal-to-noise Ca2+ probe composed of a single green fluorescent protein. Nat. Biotechnol. 2001, 19, 137–141. [Google Scholar] [CrossRef]
- Tian, L.; Hires, S.A.; Mao, T.; Huber, D.; Chiappe, M.E.; Chalasani, S.H.; Petreanu, L.; Akerboom, J.; McKinney, S.A.; Schreiter, E.R.; et al. Imaging neural activity in worms, flies and mice with improved GCaMP calcium indicators. Nat. Methods 2009, 6, 875–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyawaki, A.; Llopis, J.; Heim, R.; McCaffery, J.M.; Adams, J.A.; Ikura, M.; Tsien, R.Y. Fluorescent indicators for Ca2+ based on green fluorescent proteins and calmodulin. Nature 1997, 388, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Rose, T.; Goltstein, P.M.; Portugues, R.; Griesbeck, O. Putting a finishing touch on GECIs. Front. Mol. Neurosci. 2014, 7, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thestrup, T.; Litzlbauer, J.; Bartholomaus, I.; Mues, M.; Russo, L.; Dana, H.; Kovalchuk, Y.; Liang, Y.; Kalamakis, G.; Laukat, Y.; et al. Optimized ratiometric calcium sensors for functional in vivo imaging of neurons and T lymphocytes. Nat. Methods 2014, 11, 175–182. [Google Scholar] [CrossRef]
- Van Opbergen, C.J.M.; Koopman, C.D.; Kok, B.J.M.; Knopfel, T.; Renninger, S.L.; Orger, M.B.; Vos, M.A.; van Veen, T.A.B.; Bakkers, J.; de Boer, T.P. Optogenetic sensors in the zebrafish heart: A novel in vivo electrophysiological tool to study cardiac arrhythmogenesis. Theranostics 2018, 8, 4750–4764. [Google Scholar] [CrossRef]
- Weber, M.; Scherf, N.; Meyer, A.M.; Panakova, D.; Kohl, P.; Huisken, J. Cell-accurate optical mapping across the entire developing heart. eLife 2017, 6. [Google Scholar] [CrossRef]
- Koopman, C.D.; Zimmermann, W.H.; Knopfel, T.; de Boer, T.P. Cardiac optogenetics: Using light to monitor cardiac physiology. Basic Res. Cardiol. 2017, 112, 56. [Google Scholar] [CrossRef]
- Tsien, R.Y.; Harootunian, A.T. Practical design criteria for a dynamic ratio imaging system. Cell Calcium 1990, 11, 93–109. [Google Scholar] [CrossRef]
- Tsien, R.Y. Monitoring cell calcium. In Calcium as a Cellular Regulator; Klee, E.C.C., Ed.; Oxford University Press: New York, NY, USA, 1999; pp. 28–54. [Google Scholar]
- McMahon, S.M.; Jackson, M.B. An Inconvenient Truth: Calcium Sensors Are Calcium Buffers. Trends Neurosci. 2018, 41, 880–884. [Google Scholar] [CrossRef]
- Neher, E.; Augustine, G.J. Calcium gradients and buffers in bovine chromaffin cells. J. Physiol. 1992, 450, 273–301. [Google Scholar] [CrossRef]
- Eisner, D.A.; Caldwell, J.L.; Kistamas, K.; Trafford, A.W. Calcium and Excitation-Contraction Coupling in the Heart. Circ. Res. 2017, 121, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Mank, M.; Santos, A.F.; Direnberger, S.; Mrsic-Flogel, T.D.; Hofer, S.B.; Stein, V.; Hendel, T.; Reiff, D.F.; Levelt, C.; Borst, A.; et al. A genetically encoded calcium indicator for chronic in vivo two-photon imaging. Nat. Methods 2008, 5, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Direnberger, S.; Mues, M.; Micale, V.; Wotjak, C.T.; Dietzel, S.; Schubert, M.; Scharr, A.; Hassan, S.; Wahl-Schott, C.; Biel, M.; et al. Biocompatibility of a genetically encoded calcium indicator in a transgenic mouse model. Nat. Commun. 2012, 3, 1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heim, N.; Griesbeck, O. Genetically encoded indicators of cellular calcium dynamics based on troponin C and green fluorescent protein. J. Biol. Chem. 2004, 279, 14280–14286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, K.; Takeda, H.; Kawakami, N.; Kobayashi, M.; Matsuda, N.; Mishina, M. A transposon-mediated gene trap approach identifies developmentally regulated genes in zebrafish. Dev. Cell 2004, 7, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Urasaki, A.; Morvan, G.; Kawakami, K. Functional dissection of the Tol2 transposable element identified the minimal cis-sequence and a highly repetitive sequence in the subterminal region essential for transposition. Genetics 2006, 174, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.J.; Tu, C.T.; Hsiao, C.D.; Hsieh, F.J.; Tsai, H.J. Germ-line transmission of a myocardium-specific GFP transgene reveals critical regulatory elements in the cardiac myosin light chain 2 promoter of zebrafish. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2003, 228, 30–40. [Google Scholar] [CrossRef]
- Polito, M.; Vincent, P.; Guiot, E. Biosensor imaging in brain slice preparations. Methods Mol. Biol. 2014, 1071, 175–194. [Google Scholar] [CrossRef]
- Varkuti, B.H.; Kepiro, M.; Horvath, I.A.; Vegner, L.; Rati, S.; Zsigmond, A.; Hegyi, G.; Lenkei, Z.; Varga, M.; Malnasi-Csizmadia, A. A highly soluble, non-phototoxic, non-fluorescent blebbistatin derivative. Sci. Rep. 2016, 6, 26141. [Google Scholar] [CrossRef] [Green Version]
- Bakkers, J. Zebrafish as a model to study cardiac development and human cardiac disease. Cardiovasc. Res. 2011, 91, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Domingo, B.; Sabariegos, R.; Picazo, F.; Llopis, J. Imaging FRET standards by steady-state fluorescence and lifetime methods. Microsc. Res. Tech. 2007, 70, 1010–1021. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.H.; Kralj, J.M.; Douglass, A.D.; Engert, F.; Cohen, A.E. Simultaneous mapping of membrane voltage and calcium in zebrafish heart in vivo reveals chamber-specific developmental transitions in ionic currents. Front. Physiol. 2014, 5, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwerte, T.; Prem, C.; Mairosl, A.; Pelster, B. Development of the sympatho-vagal balance in the cardiovascular system in zebrafish (Danio rerio) characterized by power spectrum and classical signal analysis. J. Exp. Biol. 2006, 209, 1093–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, S.L.; Yang, X.; Debiais-Thibaud, M.; Schwerte, T.; Pelster, B.; Ekker, M.; Tiberi, M.; Perry, S.F. In vivo and in vitro assessment of cardiac beta-adrenergic receptors in larval zebrafish (Danio rerio). J. Exp. Biol. 2011, 214, 1445–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicente, M.; Salgado-Almario, J.; Soriano, J.; Burgos, M.; Domingo, B.; Llopis, J. Visualization of Mitochondrial Ca2+ Signals in Skeletal Muscle of Zebrafish Embryos with Bioluminescent Indicators. Int. J. Mol. Sci. 2019, 20, 5409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsutsui, H.; Higashijima, S.; Miyawaki, A.; Okamura, Y. Visualizing voltage dynamics in zebrafish heart. J. Physiol. 2010, 588, 2017–2021. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Haendchen, R.V.; Wyatt, H.L.; Maurer, G.; Zwehl, W.; Bear, M.; Meerbaum, S.; Corday, E. Quantitation of regional cardiac function by two-dimensional echocardiography. I. Patterns of contraction in the normal left ventricle. Circulation 1983, 67, 1234–1245. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biosensor | FRET Pair | Troponin C | Linkers | ΔR/R In Vitro (%) | ΔR/R Ex Vivo (%) | Kd (µM) | Decay Time (s) | Hill Slope |
|---|---|---|---|---|---|---|---|---|
| Twitch-1 | ECFP cpCit174 | Opsanus tau swim bladder and white muscle | P, P | 400 | 0.25 | 0.8 | 1.18 | |
| Twitch-2B | mCerulean3 cpVenusCD | Opsanus tau swim bladder and white muscle | VADA, PIYP | 800 | 26.5 † | 0.2 | 2.8 | 1.31 |
| Twitch-4 | ECFP cpCit174 | Opsanus tau swim bladder and white muscle | DA, PIY | 600 | 2.8 | 0.5 | 1.04 | |
| TN-XXL * | ECFP cpCit174 | Chicken skeletal muscle | RML, EL | 260 | 10 ‡ | 0.8 | 0.88 | 1.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salgado-Almario, J.; Vicente, M.; Vincent, P.; Domingo, B.; Llopis, J. Mapping Calcium Dynamics in the Heart of Zebrafish Embryos with Ratiometric Genetically Encoded Calcium Indicators. Int. J. Mol. Sci. 2020, 21, 6610. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186610
Salgado-Almario J, Vicente M, Vincent P, Domingo B, Llopis J. Mapping Calcium Dynamics in the Heart of Zebrafish Embryos with Ratiometric Genetically Encoded Calcium Indicators. International Journal of Molecular Sciences. 2020; 21(18):6610. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186610
Chicago/Turabian StyleSalgado-Almario, Jussep, Manuel Vicente, Pierre Vincent, Beatriz Domingo, and Juan Llopis. 2020. "Mapping Calcium Dynamics in the Heart of Zebrafish Embryos with Ratiometric Genetically Encoded Calcium Indicators" International Journal of Molecular Sciences 21, no. 18: 6610. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186610