Natural Products from Cyanobacteria: Focus on Beneficial Activities


1
UMR 7245 MCAM, Muséum National d’Histoire Naturelle-CNRS, Paris, 12 rue Buffon, CP 39, 75231 Paris CEDEX 05, France
2
Thermes de Balaruc-les-Bains, 1 rue du Mont Saint-Clair BP 45, 34540 Balaruc-Les-Bains, France
*
Author to whom correspondence should be addressed.
Mar. Drugs 2019, 17(6), 320; https://0-doi-org.brum.beds.ac.uk/10.3390/md17060320
Submission received: 15 April 2019
/
Revised: 21 May 2019
/
Accepted: 21 May 2019
/
Published: 30 May 2019
(This article belongs to the Special Issue Compounds from Cyanobacteria II)
Abstract
:Cyanobacteria are photosynthetic microorganisms that colonize diverse environments worldwide, ranging from ocean to freshwaters, soils, and extreme environments. Their adaptation capacities and the diversity of natural products that they synthesize, support cyanobacterial success in colonization of their respective ecological niches. Although cyanobacteria are well-known for their toxin production and their relative deleterious consequences, they also produce a large variety of molecules that exhibit beneficial properties with high potential in various fields (e.g., a synthetic analog of dolastatin 10 is used against Hodgkin’s lymphoma). The present review focuses on the beneficial activities of cyanobacterial molecules described so far. Based on an analysis of 670 papers, it appears that more than 90 genera of cyanobacteria have been observed to produce compounds with potentially beneficial activities in which most of them belong to the orders Oscillatoriales, Nostocales, Chroococcales, and Synechococcales. The rest of the cyanobacterial orders (i.e., Pleurocapsales, Chroococcidiopsales, and Gloeobacterales) remain poorly explored in terms of their molecular diversity and relative bioactivity. The diverse cyanobacterial metabolites possessing beneficial bioactivities belong to 10 different chemical classes (alkaloids, depsipeptides, lipopeptides, macrolides/lactones, peptides, terpenes, polysaccharides, lipids, polyketides, and others) that exhibit 14 major kinds of bioactivity. However, no direct relationship between the chemical class and the respective bioactivity of these molecules has been demonstrated. We further selected and specifically described 47 molecule families according to their respective bioactivities and their potential uses in pharmacology, cosmetology, agriculture, or other specific fields of interest. With this up-to-date review, we attempt to present new perspectives for the rational discovery of novel cyanobacterial metabolites with beneficial bioactivity.
1. Introduction
Cyanobacteria belong to an ancient group of photosynthetic prokaryotes that present a very wide range of cellular strategies, physiological capacities, and adaptations that support their colonization of very diverse microenvironments worldwide. As a consequence, cyanobacteria occur in varied and often extreme habitats and are then able to settle in diverse biotopes (e.g., marine, terrestrial, freshwater, thermal springs) [1,2,3]. They are also well known for the production of a wide variety of bioactive natural products, including some potent toxins (e.g., microcystins, anatoxins, and saxitoxins) [2,3]. Due to the remarkable capability of cyanobacteria to proliferate and form toxic blooms that induce potential human health consequences [4], numerous studies have been conducted to develop tools for the monitoring of such blooms [5,6] or effective strategies for the mitigation of their overgrowth [7]. On the contrary, certain cyanotoxins could also constitute a promising opportunity for drug development such as certain cancer therapies [8].
Two main aspects known as the chemical diversity and the related bioactivity have to be considered when considering the application potential of natural products produced by cyanobacteria. The chemical diversity of metabolites produced by these organisms has been well described and about 15 reviews have been already published in the past 20 years, dealing with their structural and chemical diversity [9,10,11,12,13,14] or their respective biosynthetic pathways [15,16]. Beyond the notorious harmful effects of cyanotoxins, other cyanobacterial natural products show a wide range of bioactivities that could be potentially useful for diverse applications [17,18,19,20,21]. So far, among the existing reviews related to the diversity of cyanobacterial metabolites, only one has addressed the relative taxonomical positions of the different producing strains [9]. A few taxa appear to be especially prolific producers of a large set of metabolites, while others still remain to be investigated. Recent genomics approaches and genome sequencing have been important steps in the elucidation of the pathways implicated in the biosynthesis of natural products. Their wide structural diversity has been described as a consequence of the numerous biosynthetic pathways developed by cyanobacteria in order to produce these metabolites [15]. Most of the active cyanobacterial molecules are considered as being produced either through the non-ribosomal peptide (NRP) or the hybrid polyketide-NRP biosynthetic pathways [10], or by the ribosomal synthesis of pro-peptides that are post-translationally modified (RiPP). Previous genome analysis demonstrated that the diversity of the known metabolites is merely a fraction of the true metabolic potential of cyanobacteria [15]. Concerning bioactivity, cyanobacteria have long been a source of molecules with a potent nutritional property [18]. Aztec civilizations consumed cyanobacteria (Spirulina) in their routine diet [22], and Chadian populations still use them as one of their substantial food sources [23]. Besides nutritional and probiotic purposes [13,21], cyanobacteria are well-known as an important source of metabolites with technological applications in the biotechnical or pharmaceutical fields, which lead to an increase in interest in these research realms [10]. Most bioactivities described to date are the antibacterial, antifungal, anti-cancerous, immunosuppressive, anti-inflammatory, and anti-tuberculosis activities that have the potential to be used in fields such as pharmacology, cosmetology, agriculture, the food industry, or as biofuel [17]. Cyanobacteria cells represent a sustainable resource for biotechnology due to their photosynthetic, N-fixation, and autotrophic capacities [17,18,24]. Due to the current increase in their pharmaceutical value and in their application prospects for use in medicine or biotechnology, the exploration of uncovered cyanobacterial taxa constitutes a promising strategy to efficiently explore the chemical diversity of their bioactive compounds.
The present review globally and systematically describes current knowledge on the biological activities described for cyanobacterial natural products, and, thanks to the construction of a specific and freely available molecular database, regroups all information described so far concerning the chemical structures, the producing organisms, and the various bioactivities of all the different cyanobacterial metabolite families. This original material allows us to depict, from data based on exhaustive literature, which kinds of bioactive metabolite are potentially produced by the different cyanobacterial taxa. In this case, the producer organisms were considered at different taxonomic levels (family, order, and genus) and are referenced according to their original habitats (freshwater, marine, and others). The chemical diversity is described with respect to the different kinds of bioactivity and the potential links between them are questioned, according to their potential or effective molecular mechanisms of action. A specific focus on 47 cyanobacterial compounds presenting beneficial bioactivities is detailed and discussed regarding their potential in pharmaceutical, cosmetical, biotechnical, and agricultural applications, which opens new perspectives on the discovery of novel and potent bioactive cyanobacterial molecules.
2. Methods for Dataset Construction
A database was constructed using different search engines, notably PubMed and Google Scholar. The keywords used were “cyanobacteria,” “metabolite” or “natural product,” “beneficial” and “activity,” or “biological properties.” The database was first based on reviews and further completed with recent publications dealing with the isolation of new compounds from cyanobacteria.
The main entries into the database were the names of the metabolites. To avoid bias in the counting of metabolites, we stored all the data for each molecule and its variants as a “family.” In fact, there are still no molecular classification references for a natural product description. As discussed by Janssen [25], there is no standardized naming system along cyanobacterial metabolites, as in natural product discovery in general, that could induce an underestimation of the real diversity of natural products and to hide the potential link between their chemical structures, biosynthetic pathways, and evolution routes. Thus, such a valuable classification of cyanobacterial metabolites is still needed, notably in the current context of genomic and metabolomic development.
In our database, metabolites were grouped and classified based on different criterion, initially selected by different authors [13,15,25]. First, they were classified according to their biosynthetic pathways based on the genomic data reviewed by Dittmann et al. (e.g., microcystins, cryptophycins, and aeruginosins) [15]. Secondly, when biosynthetic information was not available, metabolites were classified, according to their structural homology, as proposed by Boudreau et al., [26], Janssen [25], and Chlipala et al. [13], supposing that they might be sharing at least a similar, if not the same, biosynthetic route (e.g., kulolide-family, aerucylamides, and cyanopeptolins). In most cases, metabolite variants have a few differences occurring on a few residues and have conserved the specific structure of their metabolite family. For example, the cyanopeptolin-like family, which contains, so far described, 139 variants, is comprised of a core structure of six amino acid residues and a variable side chain containing between 1 and 3 residues. The sequence of the amino acids in the core structure is usually composed with: Thr − [Leu|Arg|Tyr] − Ahp − [Ile|Phe|Thr|Leu|Val] − N-Me[Tyr|Phe] − [Val|Ile] (see Supplementary Data S1). Some amino acids are variable (in brackets) and some others are identical in the large majority of the variants, notably the 3-amino-6-hydroxy-2-piperidone (Ahp), and the threonine (Thr) that support the side chain and close the cycle with an ester bond linkage (S1) [13].
The data collected were then classified depending on the chemical class of the compound, the chemical structure, and the strain producing the metabolites with all the taxonomic information (species, genus, family, and order), in accordance with Komarek et al. (2014) [27]. In addition, we compiled the demonstrated activities for the purified compounds. Fourteen classes of activity were mostly tested through the literature: lethality (against brine shrimp, and other small invertebrates), neurotoxicity, hepatotoxicity, dermal toxicity, cytotoxicity, anti-inflammatory activity, antioxidant activity, antiviral, antibacterial, antifungal, antialgal, antiprotozoal, serine protease inhibition, and other types of enzyme inhibition.
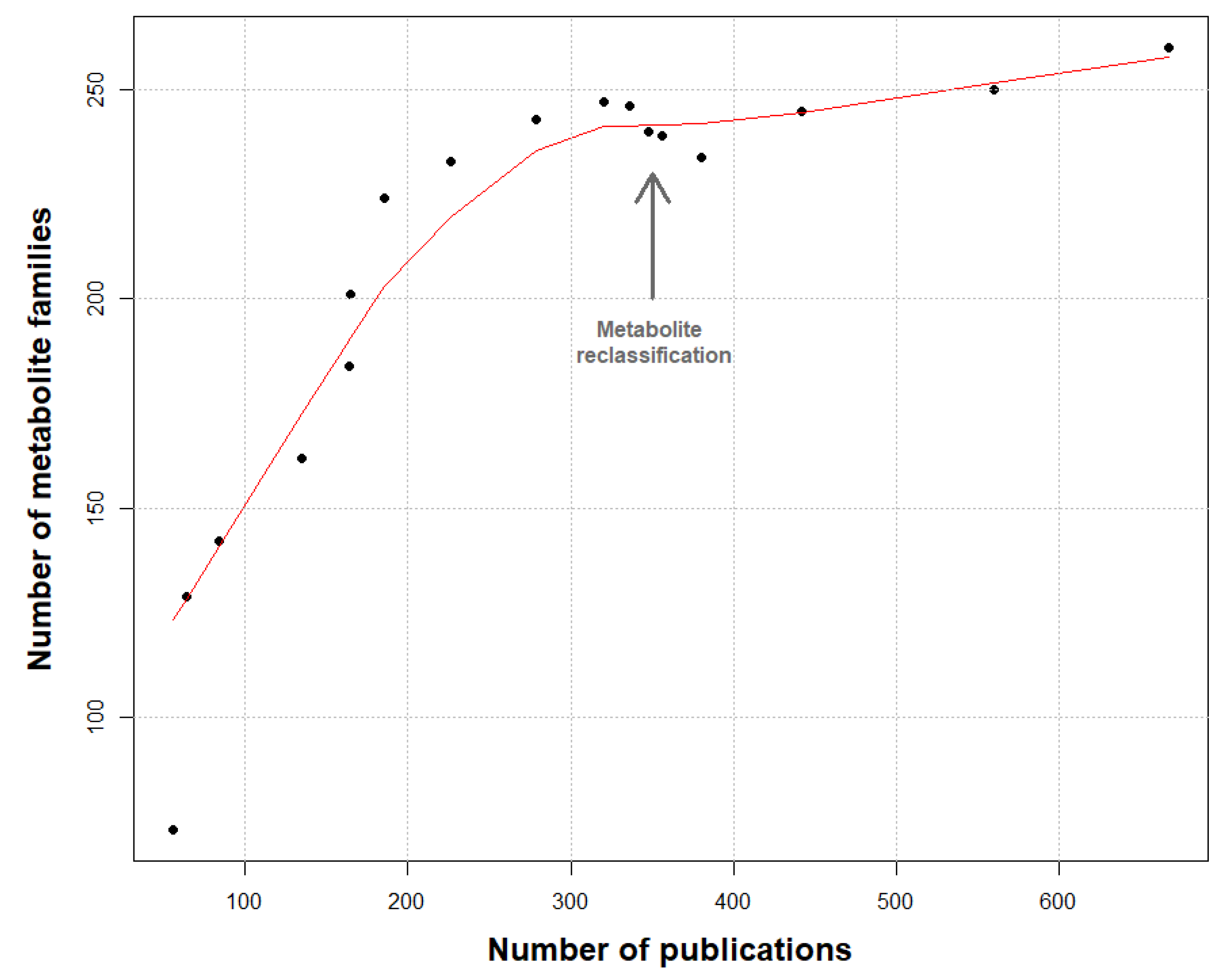
Additionally, 670 publications were analyzed, dating from the 1970s until today (April 2019). Around 1630 unique molecules have been reported so far and were grouped in 260 families of metabolites (see Supplementary Data S2). To validate the knowledge depth of our work, a rarefaction curve of the number of molecule families was constructed using the number of analyzed publications (Figure 1).
3. Taxonomy of the Producing Strains
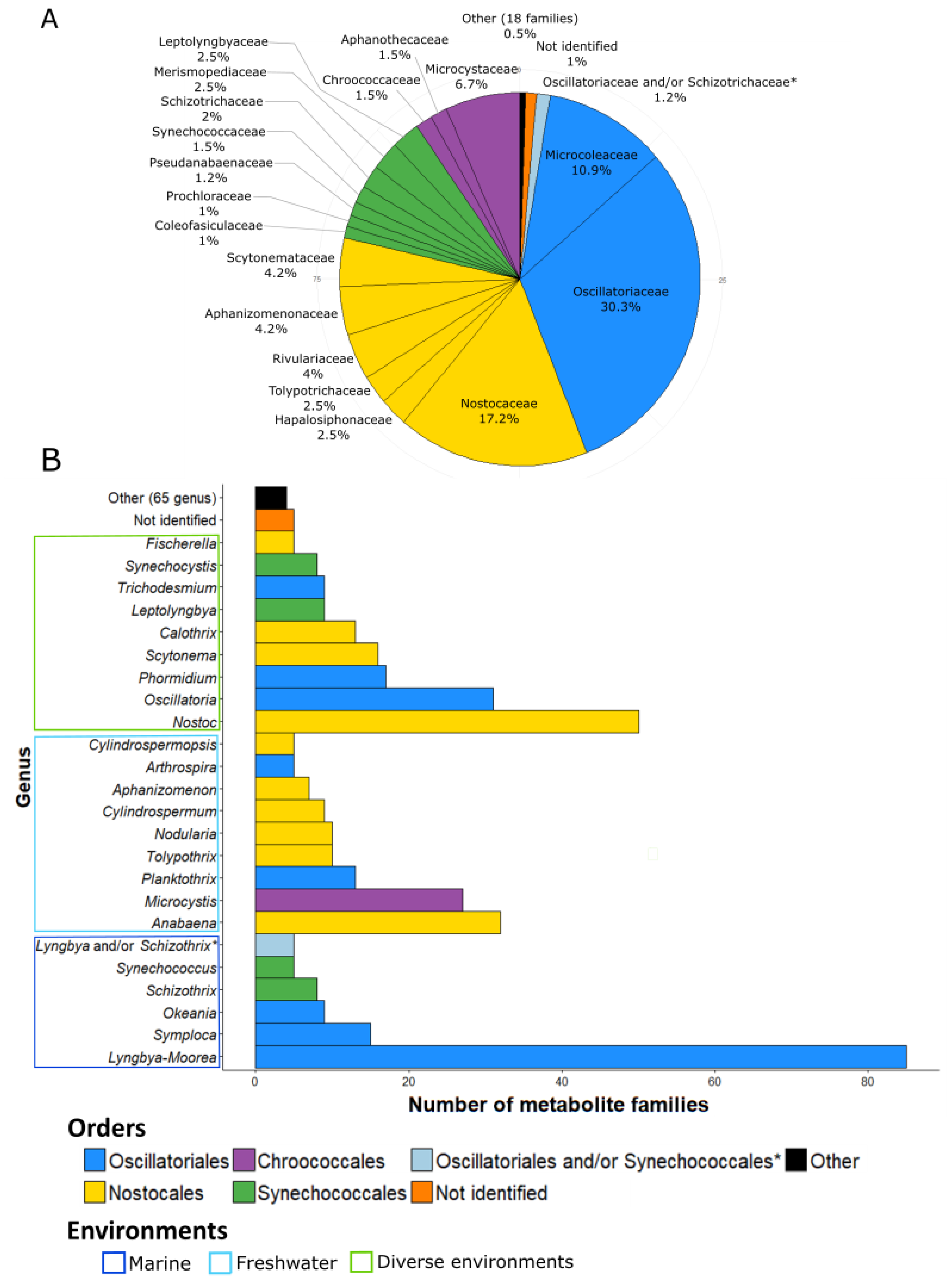
The 260 families of molecules were attributed to cyanobacteria at their different taxonomic levels (order, family, and genus) (Figure 2). Some families of compounds can be produced by different strains and, thus, occur at different taxonomical levels. For example, microcystins are produced by various strains belonging to seven different genera, five families, and three orders.
The Oscillatoriales produces the largest number of metabolite families (153 families, 46.5%). The strains belonging to the Nostocales are also considerable producers of metabolites with 98 families (29.7%). The other main producers are the strains belonging to Chroococcales and Synechococcales, which exhibit, respectively, 34 and 31 described molecule families (10.3% and 9.4%). It is interesting that, except for these four orders, the others (i.e., Pleurocapsales, Chroococcidiopsales, Gloeobacterales, and Spirulinales) remain weakly represented in the database: less than five families of metabolite have been reported so far for all of them.
Some metabolites have been isolated from cyanobacterial assemblages without accurate identification of the producer organisms. For these cases, the authors identified the genera of the two dominant cyanobacteria of the assemblage but could not accurately determine which one of them produces which molecule [28,29,30,31,32,33,34,35,36,37,38,39,40]. Tidgewell et al. (2010) [9] also identified the prevalence of marine cyanobacterial products within Oscillatoriales and Nostocales with 58% and 24% of the isolated molecules, respectively. Within Oscillatoriales, members of the genus Lyngbya, and, notably, Lyngbya majuscula produce the highest number of metabolites. This benthic genus is widely spread through the tropical marine ecosystem and has been widely studied because of its toxicity and implication in many dermatitis cases around the world [41,42]. A number of studies have been conducted on the Lyngbya genus, and a high number of new metabolites have been described. In fact, Lyngbya is, to date, the most productive genus of bioactive cyanobacterial compounds (Figure 2B). Recent studies showed that Lyngbya is polyphyletic [27,43] and using polyphasic approaches, Lyngbya has been split into four new genera: Moorea [44], Okeania [45], Limnoraphis [46], and Microseira [47]. Some marine strains previously identified morphologically as Lyngbya majuscula and Lyngbya sordida were, therefore, renamed as Moorea producens, and some strains of Lyngbya bouillonii were renamed to Moorea bouillonii on the basis of molecular and phylogenetic analyses [44]. In the same way, some freshwater strains morphologically identified as Lyngbya wollei were separate from the Lyngbya genus and described as Microseira wollei after analysis of their phylogenetic position [47].
According to this information, we decided to present the number of metabolite families produced by the Lyngbya and the Moorea genera together (reported as Lyngbya-Moorea in Figure 2B), given that the majority of families isolated from Lyngbya species were reported to be from Lyngbya majuscula (46 of 78 described from all the Lyngbya) or from Lyngbya spp. strains sampled from tropical marine environments (22 of 78), as described for the Moorea genus and were possibly misidentified with regard to this newly described genus [44].
At the family level, the main producers of known bioactive compounds belong to Oscillatoriaceae (30.3%, producing 122 families of compounds), followed by Nostocaceae and Microcoleaceae (17.2% and 10.9% for 69 and 48 molecule families, respectively) (Figure 2A). At the genus level (Figure 2B), Lyngbya-Moorea exhibits the highest number of isolated compounds (85 families of metabolites representing 20.6%), in accordance with the perceived richness of production for the Lyngbya genus due to its polyphyletic status [48]. Nostoc is the second most prolific genus of bioactive compound families with 50 isolated families so far (12.1% of the total number of families of metabolites). The other most important genera are Anabaena, Oscillatoria, and Microcystis (with 32, 31, and 27 families of molecules, respectively, representing 7.8%, 7.5%, and 6.6%) (Figure 2B).
When looking at the habitats of these cyanobacteria, a large number of compounds were isolated from marine environments (148 families of metabolite in the database, which means 53% of the families of metabolites) in comparison to the number of strains isolated from freshwater environments (77 families of metabolites, 27.6%) (Figure 2B). However, this difference might be at least partly due to the high number of compounds isolated from the marine species Lyngbya majuscula-Moorea producens (49 families of molecules, 18.8% of the families in the database) and to the existence of various research programs focused on marine species (e.g., the Panama International Cooperative Biodiversity Group, ICBG).
Overall, we observed that diversity at the genus level is important, as illustrated by the 90 different genera present in the database. Moreover, 65 different genera have been reported to produce less than four molecules (Figure 2B). We also noticed that five molecules were isolated from Lyngbya/Schizothrix assemblages and five others from unidentified strains of cyanobacteria (Figure 2B). Thus, at the genus level, the diversity of producers is large with a high number of genera studied (90 different genera). Nevertheless, these genera generally belong to the same orders (e.g., Oscillatoriales, Nostocales, Synechococcales, and Chroococcales) while some orders were not studied. For example, among the Pleurocapsales order, only four genera have been reported to produce metabolites. As a result, the covered diversity appears not to be exhaustive and can still be increased.
According to Shih et al. (2013) [49], the genomic potential of cyanobacteria to produce secondary metabolites is high with more than 70% of the studied strains presenting non-ribosomal peptide synthase (NRPS) or polyketide synthase (PKS) gene clusters in their genomes. In particular, they identified one strain belonging to the Fischerella genus (Fischerella sp. PCC 9339) that exhibits 22 NRPS/PKS clusters in its genome. On the contrary, only five compound families have been isolated from the genus Fischerella so far and are listed on the present database. Moreover, it is interesting to note that, among the 126 strains analyzed by Shih et al. (2013) [49], only 14 were formally reported to produce characterized metabolites.
In addition, the best producer genus, Lyngbya-Moorea, remains rarely studied at the genomic level: four genomes are available in the Genbank database and another three are available on the Microscope platform [50]. Considering the number of compounds isolated from the Lyngbya-Moorea genus (85 compound families), most of the links between the identified molecules and the responsible biosynthetic gene clusters remain to be characterized. We also compared our collected data with those reported by Dittman et al. (2015) [15] in order to determine when the isolated molecule families are linked with a specific and identified biosynthetic gene cluster. This review showed that less than 20% of the molecule families from the database are associated with specific identified biosynthetic gene clusters. Thus, the biosynthetic pathways of a large majority of compounds is still unknown as well as the regulation mechanisms controlling their production. Therefore, these observations highlight part of the remaining possibilities for the discovery of new molecules, gene production, and biosynthetic pathways.
4. Chemical Diversity and Bioactivity of Natural Products from Cyanobacteria
Each of the 260 families of compounds was classified by chemical classes and bioactivity (Figure 3 and Figure 4). The 260 families of compounds were classified by their chemical classes, and 10 different classes were listed: alkaloids, depsipeptides, lipopeptides, macrolides/lactones, peptides, terpenes, polysaccharides, lipids, polyketides, and others (Figure 3). Of the 260 metabolite families, 66 belong to the peptide class. Together with the depsipeptide and lipopeptide classes, they represent 133 families of compounds (51%) derived from peptides. This is not surprising, regarding the diversity of biosynthetic pathways described in cyanobacteria: NRPS (non-ribosomal peptide synthase), PKS (polyketide synthase) and RiPPs (ribosomally synthesized and post-translationally modified peptides) with the ability to produce a wide range of metabolites and notable peptides [15] (Figure 3).
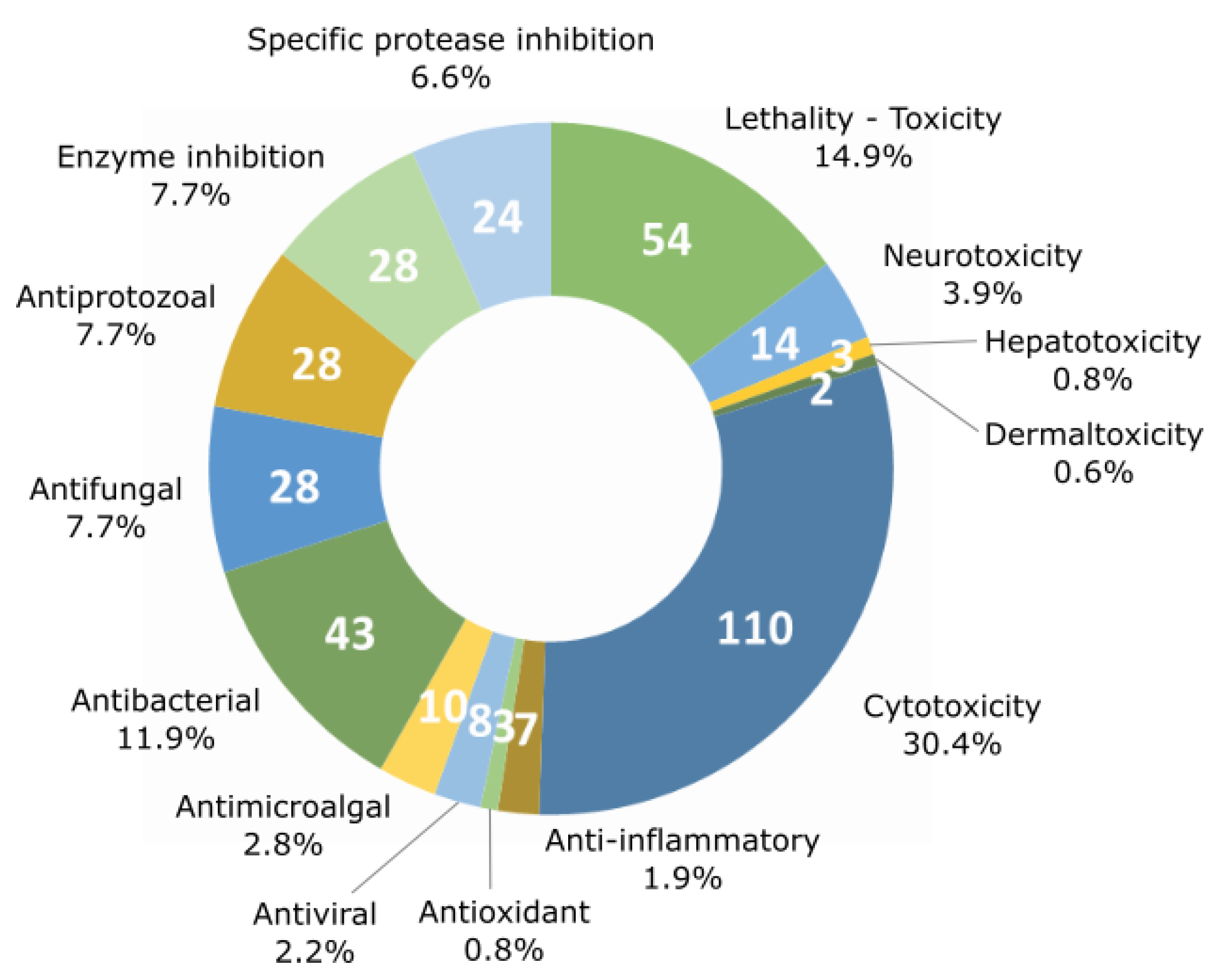
Fourteen major activities have been listed from the literature (lethality, neurotoxicity, hepatotoxicity, dermaltoxicity and cytotoxicity, anti-inflammatory, antioxidant, antiviral, anti-microalgal, antibacterial, antifungal, and antiprotozoal activities as well as protease and enzyme inhibition activities). Cytotoxic activity against various cell lines is the most frequently detected type of bioactivity with up to 110 families of the 260 listed. On the other hand, lethality and the antibacterial activities have been detected for 54 and 43 compound families, respectively (Figure 4).
The number of compounds displaying each tested activity is shown in Figure 5. The activities of molecules have been tested against different targets ranging from a specific cellular mechanism to an entire organism. For example, the inhibitory activity of proteases and other enzymes was shown to target enzymatic processes when the lethality and antimicrobial activity were tested against whole organisms. The lethality tests were generally realized against small invertebrates such as the brine shrimp crustacean Artemia salina, the gastropod mollusk Biomphalaria glabrata, and the crustacean Thamnocephalus platyurus. The present analysis confirms preceding observations (i.e., that cytotoxicity is the most commonly detected activity, followed by lethality and antibacterial activity). Some activities were detected only for a restricted number of compounds: dermaltoxicity concerned only two families of metabolites (aplysiatoxins and lyngbyatoxins) [51,52], hepatotoxicity was observed for three families (cylindrospermopsins, microcystins, and nodularins) [53,54,55], antioxidant and anti-inflammatory activities were observed for four (carotenoids, chlorophylls, mycosporine-like amino acids, and phycocyanins) [56,57,58,59], and seven metabolite families (coibacins, honaucins, aeruginosins, malyngamides, phycocyanin, scytonemin, and tolypodiol) [60,61,62,63,64,65,66], respectively. Nevertheless, there are only a few examples of these activities being tested by authors in comparison with cytotoxicity and lethality, which have been investigated far more regularly. In terms of anti-inflammatory activity, all seven tested molecules cited above were positive for this type of activity, and 53% of the studied molecule families have been tested for cytotoxic activity, while only 2.7% have been tested for anti-inflammatory activity. In parallel, some of these metabolite families can exhibit more than one activity. In fact, a total of 362 activities have been detected for the whole of the 260 metabolite families.
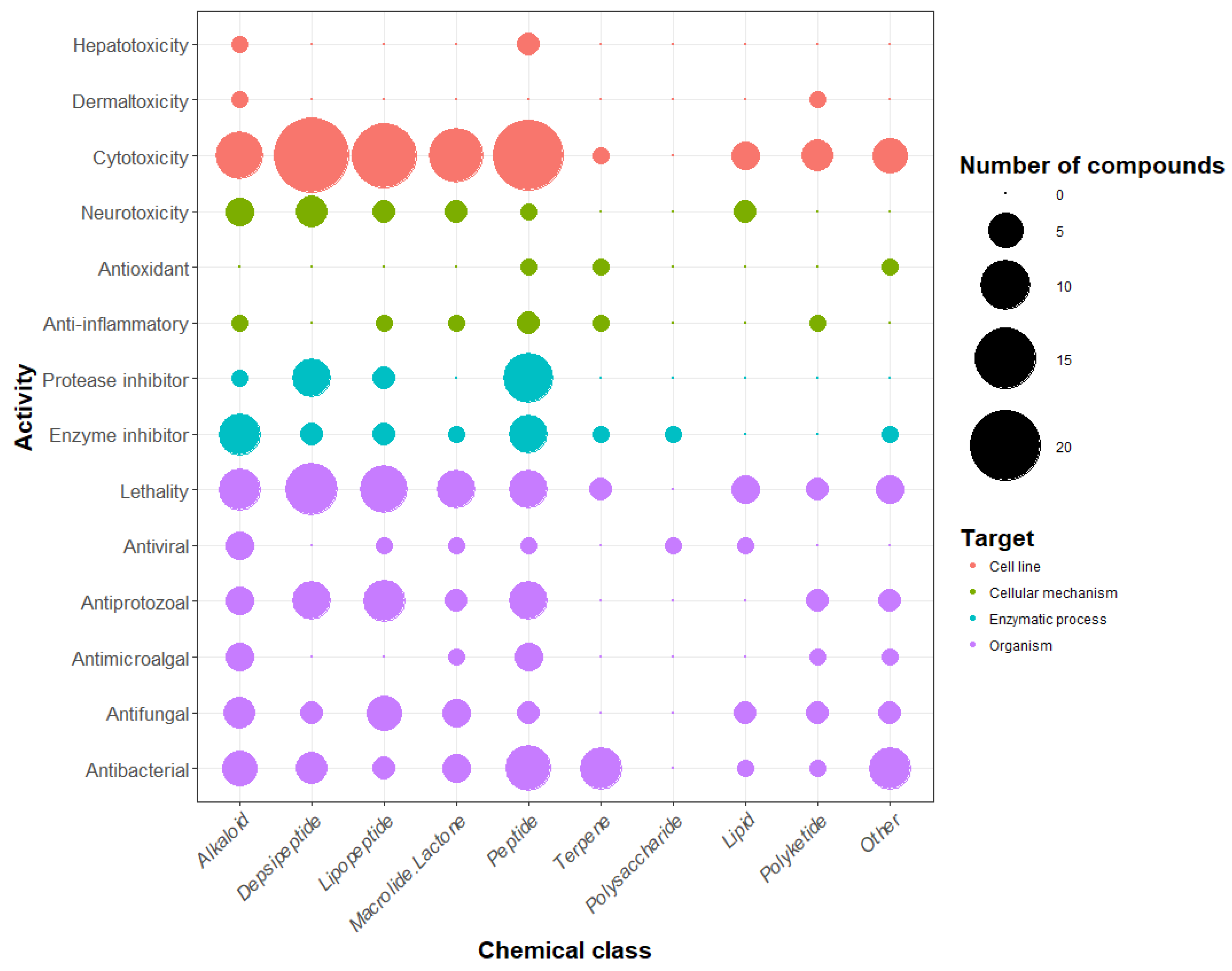
Focusing on the chemical classes, it appears that there is no specific indication that one chemical class exhibits specific activities with regard to other classes. The results from the review showed that the polysaccharide class presents only two tested activities (enzyme inhibition and antiviral activity), but only three types of polysaccharides isolated from cyanobacteria have been observed so far (calcium spirulan, cyclodextrins, and iminotetrasaccharide) [67,68,69]. Five chemical classes (the alkaloids, the depsipeptides, the lipopeptides, the macrolides, and the peptides) seem to present a remarkably large set of activities. When comparing the number of detected activities with the number of molecules belonging to each chemical class, the most bioactive molecules were shown to be the alkaloids, the lipopeptides, and the polyketides, which exhibit respectively 2.2, 1.9, and 1.8 activities per molecule on average.
These observations highlight a bias in the bioactivities searched from the isolated molecules. First, reported activities were those that researchers decided to test. Thus, the metabolite bioactivity profile could be underestimated because of the number of tests realized and remains the main limitation for the description of the potential applications of the bioactive molecules. In addition, there is still no consensus concerning the dose and dilution threshold that should be considered for each individual bioactivity test. In some cases, the concentration difference, used to determine if two distinct molecules are active, is important. For example, odoamide [70], which is a cyclic depsipeptide member of the aurilides family, and scytoscalarol [71], a sesterterpene, have both been described as being “cytotoxic.” However, their respective IC50 values appear to be very different: 26.3 nM against HeLa S3 human cervical cancer cells for odoamide and 135 µM against Vero cells for scytoscalarol, which represents a concentration difference of 500 times between their respective inhibition potentials. Furthermore, tests can be realized against several cell lines and strains presenting different sensitivity responses, which limit the comparison between results.
With 10 chemical classes and 14 types of bioactivity, the cyanobacterial metabolites are diverse and highly active. However, half of the families of metabolites listed in the database are peptides or peptide derivatives. This could be due to the importance of the peptide biosynthetic pathway (NRPS, PKS, and RiPPs) or the extraction methods used, which might favor peptide extraction. We did not observe a link between chemical classes and activities, but this observation must be considered carefully with regard to the low number of investigated molecules in some classes (i.e., polyketides, polysaccharides, and terpenes). The most frequently detected activity for cyanobacterial metabolites is cytotoxicity (42% of the metabolite families), whereas antioxidant or anti-inflammatory activities were detected for only 1.5% and 2.7% of the families. This imbalance is due to the frequency at which tests were carried out. In fact, cytotoxicity was tested for 53% of the molecules, while anti-inflammatory activity was only tested in 2.7%. This observation may reflect the research inclination to find new pharmaceutical compounds, notably cytotoxic compounds that are usable in cancer therapy, and suggests the potential for the discovery of new activities for application in other fields.
5. Beneficial Activities of Natural Products Produced by Cyanobacteria
In this review, we further considered and developed 47 examples of molecules that are considered as exhibiting potential beneficial activities for several purposes. The 260 families of compounds could have a wide range of applications, e.g., agriculture, pharmacology, cosmetology, or in the food industry. For potential applications in agriculture, cyanobacterial compounds could be useful for alternative soil fertilization methods and as chemical pesticides [18]. The potential pharmaceutical applications of cyanobacterial metabolites include the development of new antibiotics, antibacterial drugs, or antiviral drugs [21]. Metabolite families were selected because of their specific features described below in each bioactivity-related section.
5.1. Antimicrobial Activity
Antimicrobial compounds that do not present toxic effects are particularly of interest for applications in the food industry in order to clean processing equipment or for food preservation [72,73]. Cyanobacteria produce 85 families of metabolites isolated from various strains, which display potent antimicrobial activity (representing a third of the 260 molecule families listed in the database) [18]. Below, we summarize the different antimicrobial metabolites (organized by type of antimicrobial activity) that have been isolated from cyanobacteria so far and the corresponding relevant information (see Table 1, Table 2, Table 3, Table 4 and Table 5). We also detail some examples of specific molecules that exhibit interesting bioactivity profiles such as the selective profile for their activity, which present broad-spectrum action together with the absence of associated cytotoxicity.
5.1.1. Antibacterial Activity
Among the metabolite families listed, 43 molecules exhibit antibacterial activity, which represents 17% of the families. These components were, in general, tested against different types of bacteria: GRAM-negative, GRAM-positive mycobacteria, and cyanobacteria.
Among the 43 molecules, 22 are also cytotoxic and 16 present lethal activity against small invertebrates. Only three of them—eucapsitrione, kulolide-like molecules, and abietic acid—may have specific antimicrobial activity and no activity against other microorganisms.
Eucapsitrione and kulolide-like molecules (Table 1, details available in Supplementary Data S3) show antibacterial activity (against Mycobacterium tuberculosis) without inhibitory activity against the yeast Candida albicans [74,75]. Eucapsitrione is a anthraquinone derivative molecule isolated from the cyanobacterium Eucapsis sp. (UTEX 1519) [74]. This phenolic compound family is well-known in plants and some microorganisms, and has demonstrated a large range of bioactivities, including antimicrobial, antioxidant, anti-inflammatory, and potent anticancer properties [76,77,78,79]. This opens up other perspectives and applications for these anthraquinone derivatives isolated from cyanobacteria, such as eucapsitrione. However, so far, its other potential bioactivities have not been tested.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Antibacterial molecules extracted from the database and discussed in this review.
| Molecule Family | Chemical Classes | Activity | Producing Organisms | References |
|---|---|---|---|---|
| Eucapsitrione | Anthraquinone derivative | -Antibacterial -No antifungal -Cytotoxic | Eucapsis sp. UTEX 1519 | [74] |
| Kulolide-like analogs | Depsipeptide | -Antibacterial -No antifungal -Antiprotozoal -Lethal -Cytotoxic -VGSC (Voltage Gate Sodium Channel) activation | Lyngbya majuscula, Rivularia sp., Moorea producens, Okeania sp., Symploca hydnoides, Oscillatoria margaritifera | [26,75,80,81,82,83,84,85,86,87,88,89,90] |
| Abietic acids | Terpene | -Antibacterial -No lethality -No antialgal | Plectonema radiosum LEGE 06105, Nostoc sp. LEGE 06077 and LEGE 07365, Chroococcidiopsis sp. LEGE 06174, Synechocystis sp. LEGE 06079, Synechocystis salina LEGE 06099, Leptolyngbya ectocarpi LEGE 11425, Nodosilinea sp. LEGE 13457, Nodosilinea nodulosa LEGE 07084 | [91] |
| Hapalindole-like | Alkaloid | -Antibacterial -Antifungal -Antialgal -Cytotoxic -Insecticidal -Lethal activity -Reverse multidrug resistance (MDR) -VGSC modulator | Hapalosiphon fontinalis, Westiellopsis sp., Fischerella musicola, Hapalosiphon welwitschii, Westiella intricata, Fischerella ambigua, Hapalosiphon delicatulus, Hapalosiphon hibernicus, Westiellopsis prolifica, Fischerella sp., Hapalosiphon laingii | [92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117] |
More details about compound activities are available in Supplementary Data S3.
Table 2.
Antialgal molecules extracted from the database.
| Molecule Family | Chemical Classes | Activity | Producing Organisms | References |
|---|---|---|---|---|
| Cyanobacterin | Lactone derivative | -Antialgal -Anti-cyanobacterial -Growth inhibition | Scytonema hofmanni UTEX 2349, Nostoc linckia CALU 892 | [118,119,120,121] |
| Fischerellins | Polyketide | -Antialgal -Anti-cyanobacterial -Antifungal -Lethal -Growth inhibition | Fischerella musicola, Fischerella sp., Fischerella ambigua, Fischerella tesserantii | [122,123,124,125] |
| Westiellamide-like analogs | Peptide | -Antialgal -Anti-cyanobacterial -No antifungal -Lethal activity -Cytotoxic | Westiellopsis prolifica EN-3-1, Nostoc sp. 31, Stigonema dendroideum IA-45-3, Oscillatoria raoi TAU IL-76-1-2, Nostoc spongiaeforme var. tenue str. Carmeli | [126,127,128,129,130,131] |
| Ambigols | Alkaloid | -Antialgal -Antibacterial -Antifungal -Antiprotozoal -Lethal activity -Cytotoxicity -Enzyme inhibition | Fischerella ambigua 108b | [132,133] |
| Schizotrin-like analogs | Peptide | -Antialgal -Antibacterial -Antifungal -Antiprotozoal -Lethal activity -Cytotoxicity | Schizothrix sp. TAU IL-82-2, Lyngbya sp. 36.91, Phormidium sp. LEGE 05292, Tychonema sp. CCAP 1462/13 | [134,135,136,137,138,139,140,141] |
The kulolide-like family includes 44 related molecules. Kulolide, which is the first molecule of the family to be discovered, was isolated from a cephalaspidean mollusk Philinopsis speciosa [80]. Luesch and co-workers (2001) isolated the first cyanobacterial analogues of this family, naming them the pitipeptolides, and proposed a cyanobacterial origin for kulolide itself [89]. All members of the kulolide-like family share chemical similarities and can be categorized into two subgroups: those containing 2,2-dimethyl-3-hydroxy-7-octynoic acid (Dhoya) and those containing 3-hydroxy-2-methyl-7-octynoid acid (Hmoya) [26]. The same activities were not tested for all analogues, but some of them have shown antibacterial, antiprotozoal, cytotoxic, and even lethal activities (Table 1).
The third example of a family of molecule presenting a specific anti-bacterial activity is that of abietic acids (Table 1). Abietic acid is a terpene that is generally found in resin and used by conifers as a defense metabolite [91]. It demonstrates anti-cyanobacterial activity against Synechococcus nidulans, and seems to be non-toxic to Chlorella vulgaris and the brine shrimp Artemia salina (Table 1). It has been suggested that its activity and defense mechanisms could be equivalent to those of coniferous plants, i.e., trapping microorganisms or acting as allelochemical compounds. These non-toxic properties are compelling for the development of specific anti-cyanobacterial products.
The hapalindole-like group is a family of alkaloids, which contains around 80 related molecules [92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117] (Table 1). These metabolites were previously isolated from Hapalosiphon, Fischerella, Westiellopsis, and Westiella genera. They show a wide range of activity, most notably, antibacterial activity against 27 various bacterial strains, together with antifungal and antialgal activities. They are also cytotoxic and exhibit additional insecticidal activity. Some of them were even able to reverse drug resistance in cancer cell lines [97,116] (Table 1). They putatively exhibit modulatory activity on sodium channels [95], which could explain their diverse bio-activities.
5.1.2. Antialgal Activity
Antialgal activity was tested generally against microalgae, and 10 families of metabolites were shown to present such activity. Among these 10 families, four also exhibited anti-cyanobacterial activity, and it can be supposed that these molecules may be acting against general photosynthesis mechanisms. For example, cyanobacterins isolated from two strains, Scytonema hofmanni UTEX 2349 and Nostoc linckia CALU 892 [118,119], were shown to present significant antimicrobial activity directed against a large panel of microalgal and cyanobacterial strains (Table 2). These compounds also inhibit the growth of eight angiosperm plants, such as duckweed (Lemna genus), pea, corn, sorrel, black bindweed, wild oat, and green foxtail [120] (Table 2). Gleason and Case (1986) showed that this activity is due to the inhibition of the Hill reaction in photosystem II without inhibition of photosystem I [120].
Another example is the fischerellin family. These compounds were observed in four strains belonging to the Fischerella genus. They show a wide range of activities comprising growth inhibition of Lemna minor, antifungal and lethal activities, and antialgal and anti-cyanobacterial activities. Hagmann & Jüttner (1996) showed that fischerellins A is an effective inhibitor of photosystem II [123] (Table 2).
The westiellamide-like analogs family comprise 12 related cyclic peptides isolated from five strains belonging to four different genera (Table 2). The related molecules, known as the bistratamides, were previously isolated from the ascidian Lissoclinum bistratum [142], and authors hypothesized a cyanobacterial symbiont origin for these molecules [128]. This family of compounds have been shown to have anti-algal and anti-cyanobacterial activities (Table 2), but they did not show any antifungal activity against the yeast Saccharomyces cerevisiae [126,127,128,129,130,131]. Moreover, one of them, dendroamide A, has shown the ability to reverse the multi-drug resistance of a human breast carcinoma cell line (MCF-7/ADR) [126]. The MCF-7/ACR cell line overexpresses the P-glycoprotein pump, which transports drugs outside of the cell, providing higher resistance to chemical treatment. Dendroamide A is able to specifically inhibit the action of the P-glycoprotein pump, which allows the drug to penetrate and lyse the cells, so it has potential anticancer applications.
Among the antialgal compounds, two have a remarkably broad spectrum of antimicrobial activities: the ambigols and the schizotrin-like analogs families, both show antialgal, antibacterial, antifungal, and antiprotozoal activities (Table 2). Three ambigol variants were isolated from Fischerella ambigua strain 108b, while the schizotrin-like family includes 13 structurally related molecules isolated from four different strains (Table 2). In addition to these antimicrobial activities, the ambigols also present enzyme inhibition activity against cyclooxygenases and HIV-1 reverse transcriptase. The members of the schizotrin-like family, the portoamides (isolated from Phormidium sp. LEGE 05292), have also shown mitochondrial metabolism inhibition activity, which induces a further decrease in the cellular ATP content in cells exposed to portoamides [140]. This property is also promising for the development of drugs acting against tumors and cancers [143].
Via their main antialgal action (i.e., photosynthesis inhibition), the molecules have been shown to present other potential uses and could be used as alternatives to chemical herbicides based on PSII inhibition (e.g., 3-(3,4-dichlorophenyl)-1,1-dimethylurea, DCMU). These families of compounds could be used to develop new algaecides and herbicides and/or to develop new pharmaceutical drugs.
Table 3.
Antifungal molecules extracted from the database.
| Molecule Family | Chemical Classes | Activity | Producing Organisms | References |
|---|---|---|---|---|
| Hassallidins | Glycolipopeptide | -Antifungal -No antibacterial activity | Hassalia sp. B02-07, Anabaena sp. (SYKE 748A, 90y1998, 90M3, 299B, 258, SYKE763A, 0TU33S16, 0TU43S8, 1TU33S8, 1TU35S12, 1TU44S9, 1TU44S16, SYKE971/6, NIVA-CYA269/2, NIVA-CYA269/6, XPORK5C, XSPORK7B, XSPORK36B, XSPORK14D, BECID19), Anabaena cylindrica Bio33 Cylindrospermopsis raciborskii (ATC-9502 & CS-505), Aphanizomenon gracile Heaney/Camb 1986 140 1/1, Nostoc sp. (159 & 113.5), Tolypothrix sp. PCC 9009 Planktothix serta PCC 8927 | [144,145,146,147,148] |
| Lyngbyabellins | Depsipeptide | -Antifungal -No antibacterial activity -Lethal activity -Cytotoxic | Lyngbya majuscula Lyngbya sp., Lyngbya bouillonii Moorea bouillonii | [149,150,151,152,153,154,155,156] |
| Micro-guanidines | Guanidine derivative | -Antifungal -No cytotoxicity -No protease inhibition | Microcystis sp. TAU IL-306, Microcystis aeruginosa TAU IL-374 | [157,158,159] |
| Majusculamides | Lipopeptide | -Antifungal -Cytotoxic -Immunosuppressive activity -Actin filaments disrupting -Anti-settlement activity | Lyngbya majuscula, Lyngbya polychroa | [160,161,162,163,164,165,166] |
5.1.3. Antifungal Activity
Twenty-eight families of compounds showed antifungal activities. Toxicity tests were carried out against diverse fungal species, which are mostly pathogenic ones. Quite common ones include Candida albicans, Saccharomyces cerevisiae, Penicillium notatum, and Aspergillus oryzae, and less common ones include Trichophyton mentagrophytes and Ustilago violacea. Among these compounds, 11 showed several other types of antimicrobial activity in addition to antifungal activity. Only two metabolite families, hassallidins and lyngbyabellins, demonstrated specific antifungal activity without presenting any antibacterial activity. The hassallidins are cyclic glycolipopeptides isolated from three strains belonging to the Nostocales (Table 3). Four variants have been characterized so far [144,145,146,147,148], and the non-ribosomal peptide gene cluster responsible for hassallidin biosynthesis has been identified. Thus, the hassallidins cluster was detected by bioinformatics analysis of the genomes of four heterocytous cyanobacteria, Aphanizomenon gracile, Cylindrospermopsis raciborskii, Nostoc sp., and Tolypothrix sp., and hassallidins production was confirmed by LC/MS analysis (Table 3). Recently, Pancrace et al. (2017) identified the hassallidins gene cluster and characterized a new hassallidins variant from Planktothrix serta (PCC 8927), which is a nitrogen-fixing, non-heterocytous forming strain [147]. They concluded that the strain gain of the cluster occurred by horizontal transfer and, therefore, questioned the natural product distribution and diversity among cyanobacteria.
The lyngbyabellins are cyclic depsipeptides. They were isolated from Lyngbya and Moorea species (Table 3). Hectochlorin is the only member of the family that was tested for antibacterial and antifungal activity, which showed no antibacterial activity but displayed antifungal activity against Candida albicans [152]. The distinctive feature of the lyngbyabellins is that they can also disrupt actin filaments. Luesch et al. (2000) [151] and Han et al. (2005) [150] showed that cells exposed to Lyngbyabellin A and E lost their microfilament network, which caused cell cycle arrest at the cytokinesis phase. Marquez et al. (2002) [152] showed that the same process appears with cells exposed to hectochlorin. They also demonstrated that the molecule stimulates actin polymerization and then induces cell cycle disorders.
Microguanidines are guanidine derivatives isolated from two strains of Microcystis (Table 3). These molecules showed antifungal activity against Saccharomyces cerevisiae E4orf4 without cytotoxic activity. This specificity could be of interest for the development of new antifungal products [158].
Majusculamides are lipopeptides produced by Lyngbya majuscula and Lyngbya polychroa. These metabolites combine antifungal and cytotoxic activities with immunosuppressive and anti-settlement properties [160,161,162,163,164,165,166]. Simmons et al. (2009) [165] also demonstrated the ability of majusculamides to disrupt actin filaments that may explain these specific properties (Table 3).
5.1.4. Antiviral Activity
Viral diseases are one of the main health concerns around the world. According to the World Health Organization (WHO), HIV and AIDS caused around one million deaths in 2017 [167]. We noted that eight families of cyanobacterial compounds have shown antiviral activity. Antiviral activity was generally determined by testing against the human immunodeficiency virus (HIV-1 or HIV-2) or the Herpes simplex virus (HSV-1 or HSV-2). The aplysiatoxins showed activity against Chikungunya’s virus (CHIKV) [168] (Table 4), but are also very active dermatotoxins [51,169] and tumor-promoting molecules due to their capacity to activate protein kinase C (PKC), which is an enzyme that plays roles in cell proliferation, differentiation, and apoptosis [168] (Table 4). Recently, Han et al., demonstrated that two aplysiatoxin analogues showed the capability to inhibit the potassium channels [170], which opens interesting perspectives for the study and use of these molecules for drug development.
Table 4.
Antiviral molecules extracted from the database.
| Molecule Family | Chemical Classes | Activity | Producing Organisms | References |
|---|---|---|---|---|
| Aplysiatoxins | Alkaloid | -Antiviral -Dermatitis and swimmer’s itch agents -Cytotoxic | Lyngbya majuscula, Schizothrix calcicola, Oscillatoria nigro-viridis, Trichodesmium erythaeum | [51,168,170,171,172,173] |
| Cyanovirin-N | Protein | -Antiviral -No cytotoxicity -Stop fusion and transmission of HIV-1 virus | Nostoc ellipsosporum Cyanothece sp. | [174,175,176] |
| Calcium spirulan | Polysaccharide | -Antiviral -No cytotoxicity -Low anticoagulant activity | Arthrospira platensis | [67,177,178] |
Two other families of molecules have shown antiviral activity against a large panel of viruses. The first one, cyanovirin-N analogs, have been isolated from Nostoc ellipsosporum [174] and Cyanothece sp. [176] (Table 4). These molecules are proteins belonging to the lectins class because of their ability to bind glycans. Cyanovirins show inhibitory activity against HIV-1, HIV-2, simian immunodeficiency virus (SIV), feline immunodeficiency virus, HHV-6, and measles virus [174,175]. Also, they inhibit Ebola and influenza viruses [176]. Nevertheless, cyanovirins are not active against some viruses, such as human herpesvirus A (HHV-1), cytomegalovirus, and adenovirus type 5 [175]. Cyanovirins are also non-cytotoxic for non-infected cells (at concentrations required for antiviral activity) [174,175] (Table 4). In fact, cyanovirin-N binds gp120, which is a glycoprotein component of the HIV envelope. As a result, the molecule inhibits membrane fusion into target cells and stops virus transmission. Calcium spirulan has been isolated from Arthrospira platensis (anc. Spirulina platensis) and is a sulphated polysaccharide. It shows antiviral activity against a wide range of viruses including HIV-1, HSV-1, the human cytomegalovirus (HCMV), measles virus, mumps virus, and influenza virus, in addition to a low cytotoxicity against several cell lines (ID50 values between 2900 and 7900 µg/mL) (Table 4) [67,178]. Furthermore, calcium spirulan seems inactive against Poliovirus and Coxsackievirus, two non-enveloped viruses, which means that it likely has selective activity for enveloped viruses. Hayashi et al. (1996) [67] also showed that this molecule inhibits virus penetration in targeted cells. Other sulphated polysaccharides are known for their anticoagulant and antiviral activity, such as heparin or dextran sulphate [179,180]. In comparison to these molecules, calcium spirulan showed a lower anticoagulant activity and a longer half-life in blood [177], which confirms its promising potential for the development of new specific antiviral drugs.
5.1.5. Antiprotozoal Activity (Against Malaria, Leishmaniosis, Chagas Disease)
The last kind of antimicrobial properties tabulated is antiprotozoal activity. Protozoans are eukaryotic microorganisms, some of them have parasitic lifestyles and are well-known for their involvement in human diseases such as Malaria, Leishmaniosis, Chagas’ disease, and Trypanosomiasis. These diseases represent a huge problem in tropical countries where the parasite is transmitted by mosquitoes. The WHO identified more than 210 million Malaria cases in 2016 [181]. Therein, molecules with antiprotozoal activity are actively being sought in order to develop new drugs against these diseases.
Table 5.
Antiprotozoal molecules extracted from the database.
| Molecule Family | Chemical Classes | Activity | Producing Organisms | References |
|---|---|---|---|---|
| Companeramides | Depsipeptide | -Antiprotozoal -No significant cytotoxicity | Leptolyngbya sp. or «Hyalidium» | [29] |
| Hoshinolactam | Lactam | -Antiprotozoal -No cytotoxicity | Oscillatoria sp. | [182] |
| Dolastatins | Peptide | -Antiprotozoal -Lethal -Cytotoxic | Lyngbya majuscula, Symploca hydnoides, Lyngbya sp., Symploca sp. VP642, Lyngbya-Schizothrix assemblage | [30,32,75,183,184,185,186,187,188,189] |
From the review, 28 cyanobacterial metabolites showed antiprotozoal activities. Tests have been conducted against several strains of Plasmodium falciparum (causative agent of Malaria), Leishamania donovani (Leishmaniosis), Trypanosoma cruzi (Chagas’ disease), and Trypanosoma brucei (Sleeping sickness). Among the 28 concerned families of molecules, 19 showed antiprotozoal activity against drug-resistant strains, especially against chloroquine-resistant strains of Plasmodium falciparum (see Table 5). Nevertheless, most of them are less active than the antibiotics currently used. For example, companeramides are cyclic depsipeptides produced by a cyanobacterium previously identified as Leptolyngbya sp. (now Hyalidium) [29] (Table 5). Companeramides showed antimalarial activity against three strains of chloroquine-resistant Plasmodium falciparum. They also showed no significant cytotoxicity against the cell lines used in the test, which constitutes a unique property for the development of specific but non-toxic antimalarial drugs. Unfortunately, the activity of companeramides against the parasite is 100-fold lower than that of chloroquine (a commonly used drug), which reduces their potential utilization.
However, some molecules show promise as substitutes for antibiotic treatment because of their strong activity against the parasite. This is the case for hoshinolactam and dolastatins. Hoshinolactam is an aromatic molecule belonging to the lactam chemical class [182]. It was isolated from an environmental sample rich in Oscillatoria sp. and has shown antiprotozoal activity against Trypanosoma brucei (IC50 = 3.9 nM) with no cytotoxicity against MDR-5 (the host cell, IC50 > 25 µM) (Table 5). Furthermore, the IC50 of pentamidine (another commonly used drug) against Trypanosoma species is 4.7 nM. Thus, the activity of hoshinolactam is equivalent to that of the antibiotics, and hoshinolactam represents a promising alternative to pentamidine for Trypanosomiasis treatment [182].
Dolastatins are a well-studied family of peptides. The first members of this family were isolated in 1977 from the sea hare Dolabella auricularia [190]. In 1998, other molecules belonging to the dolastatins family were isolated from the cyanobacteria Lyngbya majuscula and Symploca hydnoides, which leads to the hypothesis that dolastatins isolated from the mollusc have a cyanobacterial dietary origin [191]. Dolastin 10, which is one of the dolastatin-related molecules, is the most potent antiprotozoal metabolite discovered so far from cyanobacteria. This exhibits an IC50 of 0.1 nM (the IC50 of chloroquine is, on average, 5 nM for the chloroquine-sensitive strain of P. falciparum) [184]. Dolastatins are also strongly cytotoxic molecules (Table 5). They are able to inhibit tubulin polymerization, which induces cellular cycle arrest and apoptosis [192]. Antiprotozoal and cytotoxic activities are both the result of this property. Therefore, there is no apparent specificity for this molecule to act directly against the parasite itself, where the cellular host is likely the most potent target of dolastatins. For this reason, Fennel et al. (2003) [184] concluded that dolastatins do not constitute a promising antiprotozoal drug despite their strong activity.
5.2. Potential Anticancer Activity
Currently, cancers constitute the most important non-transmittable diseases worldwide. According to the WHO, cancer was the cause of one in six deaths (9.6 million) in 2018 [193]. The annual cost of cancer in 2010 was estimated to be $1.16 trillion USD [194]. That is why numerous studies have been conducted to understand the physiology of different cancers and to find new efficient anticancer drugs. For this purpose, researchers are looking for molecules, and, notably, natural products that are able to kill cells or inhibit cell proliferation. In this section, we selected some particular cyanobacterial metabolites for which the mode of action is known as significant examples. We feature metabolites acting on the microtubule or the microfilament network, histone deacetylase and proteases inhibitors, and molecules with the ability to reverse multi-drug resistance.
5.2.1. Cytotoxic Activity
The first type of activity test was performed to determine the potential of molecules as anticancer agents due to cytotoxicity. Different cell lines derived from tumor cells, like the HeLa cell line (derived from cervical cancer), KB (HeLa derivative), LoVo (human colon tumor), H-460 (human lung cancer), and MCF-7 (human breast cancer) have been used to assess this activity. Most of the time, the molecules investigated were tested against two or more cell lines to detect a potent specificity and to evaluate their potential for drug development. According to this review, 110 families of metabolites isolated from cyanobacteria showed cytotoxicity, which represents 43% of the molecule families listed in the database.
The best example of potent anticancer molecules derived from cyanobacteria is the dolastatin family [191]. One synthetic analogue of dolastatin 10, monomethyl auristatin E, is actually used to treat Hodgkin’s lymphoma in the drug Brentuximab vedotin [191]. Luesch et al. (2001) [187] showed that dolastatin 10 and symplostatin 1 are 100-fold more efficient than vinblastine (anticancer drug extracted originally from the Madagascar periwinkle) against the same cell line due to their ability to depolymerize microtubules. Unfortunately, dolastatins also have strong cytotoxicity [187,195]. Researchers found a way to reduce this toxicity by coupling monomethyl auristatin E with a chimeric antibody against CD30 (tumor necrosis factor receptor, highly expressed in Hodgkin’s lymphoma) in order to target only tumor cells [196]. Since then, other antibody drugs linked (ADC) with monomethyl auristatin E have been developed. For example, glembatumumab vedotin is currently under clinical trial. This drug targets GPNMB (glyprotein non-metastatic melanoma protein B), which is a glycoprotein expressed in melanoma and breast tumors [197]. In addition to the dolastatins, other cyanobacterial metabolites destabilize the microtubule network. Notably, one such metabolite is tubercidin, which is a nucleoside produced by Tolypothrix byssoidea, Tolypothrix distorta, Plectonema radiosum, and Scytonema saleyeriense var. indica [198,199]. This molecule was previously isolated from the bacterium Streptomyces tubercidicus. Tubercidin has shown inhibition of cell proliferation with an IC50 of 248 nM (Table 6). Tubercidin acts against dolastatins showing a microtubule stabilizing activity comparable to taxol bioactivity [200]. Its cytotoxicity is due to its stabilizing property, which causes mitotic arrest at G2/M transition and stops growth [201].
Another mechanism of cytotoxicity noted from cyanobacterial metabolites is the destabilization of actin microfilaments. As tubulin microtubules, actin microfilaments are key cytoskeleton components of cells. Microfilaments are involved in several mechanisms: cell division (cytokinesis), cell motility, cell adhesion, exocytosis, and endocytosis [211]. Thus, molecules with actin-modulating activity are sought in order to develop anticancer drugs because of their ability to induce apoptosis [211]. Four cyanobacterial metabolite families have shown disrupting activity of the actin microfilament network: the lyngbyabellins, the majusculamides, the aurilides, and the swinholide-like molecules (Table 6).
Lyngbyabellins and majusculamides, as mentioned above, have shown antifungal activity that likely corresponds to their ability to modulate actin polymerization [150,151,152,165]. Aurilides are cyclic depsipeptides, and the first member of this family was isolated from the sea hare Dolabella auricularia [212]. Since then, seven other related molecules have been isolated from two cyanobacterial genera: Lyngbya and Okeania [70,202,203,204,205], and one from Philinopsis speciosa (cephalaspidean mollusc) [213]. Aurilides showed nanomolar cytotoxic activity associated with a moderate toxicity to Artemia salina. Two analogues, lagunamides A and B, have also shown antimalarial activity and anti-swarming activity against Pseudomonas aeruginosa [203] (Table 6). Han et al. (2006) [202] showed that aurilides induce microfilament disruption at the micromolar level. They concluded that this disrupting activity is likely related to their toxic and antimicrobial activities.
Swinholide-type molecules were macrolides, originally isolated from the sponge Theonella swinhoei [214]. In 2005, Andrianasolo et al. (2005) [206] succeeded in isolating swinholide A and two new related molecules (ankaraholides A and B) from two cyanobacteria (Symploca sp. and Geitlerinema sp., respectively), which leads to the hypothesis of a symbiotic origin of the compounds isolated from the sponge [206] (Table 6). More recently, Humisto et al. (2018) identified the swinholide biosynthetic cluster in Nostoc sp. (Table 6) [207], and Tao et al. (2018) isolated nine swinholide-related metabolites from a marine Phormidium sp. [208]. Swinholide A, isolated from the marine sponge, showed microfilament-disrupting activity by stabilizing actin dimers [215]. In addition to their cytotoxic activity, cyanobacterial swinholides also showed the same actin-disrupting activity, which is of interest for the development of related anticancer drugs [206].
Other metabolites with noticeable cytotoxicity are anabaenolysins, which are lipopeptides isolated from two strains of the Anabaena genus [210] (Table 6). Anabaenolysins showed cytotoxicity against all of the 10 cell lines tested, with LC50 values between 4 and 20 µM depending on the cell lines and the anabaenolysin variants [210]. In addition, using a trypan dye exclusion assay, these authors showed that anabaenolysins have a unique profile. Instead of excluding the dye, cells showed an influx of trypan dye, which means that anabaenolysins permeabilize cell membranes until necrotic death [210]. Anabaenolysins are able to solubilize the lipid component of the cell membrane, and likely acts with the same mechanism as the detergent digitonin. Anabaenolysins particularly target cholesterol-containing membranes and do not induce permeabilization of mitochondria membranes. As detergents, anabaenolysins also show hemolytic activity, but at lower concentrations than digitonin and surfactin [209]. In addition, Oftedal et al. (2012) showed that the permeabilization ability of anabaenolysins also allows the internalization of nodularin [209]. This property is of interest for the development of a drug administration strategy involving anabaenolysins as a synergistic compound and other bioactive molecules that cannot be passed through the membrane within the targeted cells alone.
Six cyanobacterial families of compounds showed the ability to reverse multi-drug resistance (MDR) in addition to their cytotoxic properties. These include the cryptophycins [216,217,218,219], the hapalindole-like metabolites [116], hapalosin [220], the patellamides [221,222], the tolyporphins [223,224], and the westiellamide-like [126] molecules (see Supplementary Data S3). Among them, five families displayed MDR reversal activity by acting on the P-glycoprotein pumps (except for cryptophycins and patellamides for which the MDR reversal mechanisms have still not been described). P-glycoprotein is a glycosylated transmembrane protein that transports drugs and toxins out of the cell. This protein is often overexpressed in cancer cells and leads to resistance against standard chemotherapeutics because of its lower accumulation in targeted cells [225]. Thus, metabolites with the ability to inhibit this efflux pump are of interest for developing anticancer drugs or to supplement current chemotherapeutic strategies in order to increase their efficiency on resistant cancer cells.
5.2.2. Protease Inhibitory Activity
Proteases are a widespread family of enzymes found in most, if not all, organisms. They are involved in a large number of pathways including coagulation, inflammation, digestion, hemostasis, and blood pressure regulation [226,227]. There are several types of proteases that are classified by their specific hydrolysis mechanisms. The major groups are the metalloproteinases, the serine proteases, the cysteine proteases, the threonine proteases, and the aspartic acid proteases [227]. Because of their ubiquity, these enzymes are attractive targets for the development of new drugs against diverse diseases [226]. Some proteases have also shown the potential to act against thrombotic diseases [226], hypertension [227], pulmonary diseases [228], asthma [229], pathogenic microorganisms [230,231], and even cancers [227,232]. According to our investigation, 24 families of metabolites presenting diverse protease inhibitor activities have been isolated from cyanobacteria to date. These compounds have shown inhibitory activity against a wide range of proteases, including enzymes belonging to the cathepsin family or the well-known serine proteases trypsin, chymotrypsin, and thrombin. Only three metabolite families have shown an inhibitory activity against cathepsins. Cathepsins are frequently overexpressed in cancer cells and are involved in tumorigenesis, cell invasion, and metastasis [233,234,235,236,237,238]. One of these three families, the spumigins, isolated from Nodularia spumigena and Anabaena compacta [239,240,241], is a set of linear peptides that are structurally similar to the aeruginosins (Table 7). They showed inhibitory activity against several proteases including trypsin, thrombin, plasmin, and cathepsin B [240]. All of these proteases are potentially involved in cancer cell processes, and, notably, cathepsin B has been proposed to be a promising target for anticancer drug development [234,242].
Another example of metabolites with interesting activity is the cyanopeptolin-like family. This family is the second in terms of the number of structural analogues isolated, after the microcystins (respectively 140 and 246 molecular variants described so far). Currently, more than 50 papers have reported on the isolation and activities of these metabolites. They are cyclic depsipeptides isolated from 12 different cyanobacterial genera (Table 7). Among the large number of analogs, a wide range of activity has been reported for these cyanobacterial metabolites including protease activity and other types of enzyme inhibition, cytotoxicity, lethal activity, and antimicrobial activity, which open various possibilities for developing therapies targeting cancer cells or microorganisms or those that fight some diseases like emphysema [277], pancreatitis [295], or thrombosis [296]. Nevertheless, this large number of activities can also represent a problem such as how to develop a therapeutic drug exhibiting a specific activity. It would be interesting to study some analogs in more depth or to conduct a structure–activity relationship study in order to increase the specificity of synthetic variants.
Lastly, another class of inhibitors that would be of interest for the development of new therapeutics against tumors is the proteasome inhibitors. Proteasome or ubiquitin-proteasome is a multi-enzymatic complex of eukaryotes. It is involved in protein degradation in a different way than the lysosomes [296]. Because proteasome catalysis is involved in a wide variety of essential pathways, including cell-cycle progression and the regulation of apoptosis, it is a potent target for cancer therapy. Moreover, malignant cells have been shown to be more affected by proteasome inhibitors than normal cells, which reduce the potentially deleterious side effects of these molecules [232]. Four cyanobacterial families of metabolites were described to inhibit the 20S core of proteasome: the carmaphycins [293], the cylindrocyclophanes [169,297,298,299,300,301,302,303], the nostocyclopeptides [304,305], and nostodione [306,307] (see Supplementary Data S3). Among them, the most efficient 20S proteasome inhibitors are the carmaphycins, which exhibit an IC50 of around 2.5 nM [293], whereas the other compounds present a micromolar range of action [169,305,306] (Table 7). Only two carmaphycin variants (A and B) have been isolated from Symploca sp., so far. These molecules are linear peptides with cytotoxic and antiprotozoal activities. They show the additional ability to inhibit the 20S proteasome activity in yeast and Plasmodium by interacting with the β5 subunit [293,294]. These bioactivities are interesting for the use of carmaphycins as anticancer or antimalarial therapeutics. Two studies were conducted to enhance the specificity of carmaphycins for either applications. To develop a specific antimalarial drug, LaMonte et al. (2017) synthesized synthetic analogues of carmaphycin B and identified one analog with a selectivity index of 380 for antiprotozoal activity over cytotoxic activity [294]. On the other hand, Almaliti et al. (2018) studied the potential of carmaphycins as anticancer drugs and as an antibody–drug conjugate (ADC) in order to enhance the selectivity of the molecules for cancer cells and to reduce the potential side effects [308].
Therefore, cyanobacterial metabolites with protease inhibition activities were shown to be less specific for further use, but the synthesis of synthetic analogs increased the selectivity of some of these molecules.
5.2.3. Histone Deacetylase Inhibitors
Histone deacetylases (HDACs) are enzymes involved in re-modeling the chromatin and the acetylation/deacetylation of histone and non-histone proteins. Furthermore, histone deacetylases play a key role in histone–DNA interactions and in the binding to transcription factors. HDACs have also been identified as potent regulators of gene expression [309,310]. Because cancer generally emerges from genetic mutations inducing hyperactivation of oncogenes or loss of tumor-suppressor genes, targeting mechanisms that are involved in the epigenetic regulation of genes is a promising strategy for the development of anti-tumor drugs [310].
Two molecules showing histone deacetylase inhibitory activity have been isolated from cyanobacteria so far, which are known as largazole and santacruzamate A. Both of these molecules come from Symploca sp. strains (Table 8). Largazole has shown inhibition against 12 class I HDACs in addition to inhibition of the ubiquitin-activating enzyme (E1). It has also shown cytotoxicity to several cell lines (Table 8). Largazole acts as a pro-drug—the molecule needed to be activated by hydrolysis to release its active form, the largazole thiol [309]. Santacruzamate A has also shown histone deacetylase inhibition and cytotoxicity. It shares some structural features with suberoylanilide hydroxamic acid (SAHA), which is a clinically approved HDAC inhibitor that is used to treat refractory cutaneous T-cell lymphoma [317]. Salvador-Reyes and Luesch (2015) performed an in-depth review of the activities and mechanisms of action of these two metabolites [309]. They highlighted the high potency of largazole in anticancer drug development, while the potency of santacruzamate seems to remain more limited.
5.3. Anti-Inflammatory and Antioxidant Activity
In this section, we specifically describe cyanobacterial metabolites that showed no cytotoxicity associated with their anti-inflammatory or antioxidant properties.
5.3.1. Anti-Inflammatory Activity
According to our review, seven metabolite families isolated from cyanobacteria were found to have anti-inflammatory activity (aeruginosins, coibacins, honaucins, malyngamides, phycocyanin, scytonemin, and tolypodiol).
Currently, anti-inflammatory molecules have been widely studied in order to develop new therapeutics directed against chronic inflammatory diseases, such as rheumatoid arthritis, psoriasis, chronic obstructive pulmonary disease, multiple sclerosis, and inflammatory bowel disease [318]. Anti-inflammatory compounds can also be useful against cardiovascular diseases, such as atherosclerosis [319], and neurodegenerative diseases like Parkinson’s disease [320].
Anti-inflammatory tests have been performed in vitro or in vivo in mice. For example, malyngamides have been shown to inhibit superoxide production generated by inflammation-promoting agents [332], and honaucins inhibit pro-inflammatory cytokine expression [61] in the murine macrophage cell line RAW264.7. The mouse ear edema assay has been performed in vivo by observing the resorption of ear edema in the presence of anti-inflammatory compounds, such as phycocyanin [64], scytonemin [330], and tolypodiol [66], which have shown noteworthy activities by using this assay.
Three metabolites seem to be particularly interesting according to their specific bioactivity profiles: the aeruginosins, phycocyanin, and scytonemin, which have not shown any toxicity when tested in vitro or in vivo. Aeruginosins have shown anti-inflammatory properties using the AlphaLISA assay. They are able to down-regulate the level of pro-inflammatory mediators (IL-8 and ICAM-1) in stimulated endothelial cells [62] without affecting the viability of two different cell lines [62] (Table 9). Aeruginosins have also shown serine protease inhibitory activity against trypsin, thrombin, and plasmin [321], and their corresponding biosynthetic gene cluster was first identified in Planktothrix agardhii and Nodularia spumigena (Table 9) [241,322]. Currently, no correlation between serine protease inhibition and the anti-inflammatory activity of aeruginosins were shown. However, on neutrophils, it has been shown that some serine proteases (elastase, cathepsin G, and proteinase 3) are responsible for the conversion and activation of proinflammatory chemokines (and notably, interleukine-8 (IL-8)) and are able to conserve or enhance the inflammation response [333,334,335]. In this regard, it will be compelling to further test whether aeruginosins are capable of inhibiting other serine proteases, notably elastase, cathepsin G, and proteinase 3, in order to determine whether the down-regulation of IL-8 induced by the aeruginosins is mediated through serine protease inhibition processes.
Phycocyanin is a phycobiliprotein, constituting one of the major cyanobacterial pigments, together with the chlorophylls and phycoerythrin. It is involved in light-harvesting and the energy transfer of phycobilisomes within the outer membrane of thylakoids. In addition, phycocyanin has shown a wide variety of beneficial properties including antioxidant, anti-inflammatory, neuroprotective, and hepatoprotective activities [64] (Table 9). Authors of phycocyanin studies have reviewed the main features of phycocyanin anti-inflammatory mechanisms. Phycocyanin is able to scavenge ROS, has anti-lipoperoxidative effects, and inhibits cyclooxygenases (specifically COX-2) as well as TNF-α release. All of these properties are unique from the perspective of new therapeutics development targeting neurodegenerative diseases such as Alzheimer’s disease, Parkinson’s disease, or Huntington’s disorder, or as an anti-inflammatory agent [64].
Scytonemin is an alkaloid pigment found in the sheath of some cyanobacteria and, particularly, on some organisms living in extreme environments [65]. Scytonemin synthesis is mainly induced by UV-A exposure in order to reduce heating and the oxidative stress [65]. Scytonemin is mainly involved in photoprotection by UV-absorption [65]. It has also been shown to have anti-inflammatory activity with no cytotoxicity against non-proliferating cells [65,330,331]. In addition, scytonemin has been shown to inhibit polo-like kinase 1 (PLK1), which is an enzyme involved in the phosphorylation and activation of proteins such as cdc25C, which is involved in cell cycle progression and the G2/M transition in the cell cycle (Table 9). As a consequence, scytonemin can repress cell proliferation [65,330,331]. Therefore, scytonemin could be a promising compound for use in the development of anticancer therapeutics, sunscreen agents, or anti-inflammatory drugs.
Last but not least, as mentioned above, ambigol have been shown to inhibit cyclooxygenases. Cyclooxygenases are enzymes belonging to the oxidoreductase enzymatic class. Two related isoforms, COX-1 and COX-2 [336], have been discovered so far and are involved in inflammation processes through the synthesis of prostaglandins from arachidonic acid. Some classical anti-inflammatory molecules are known to target COX. For example, aspirin, which is the most famous COX inhibitor discovered so far, is a nonsteroidal anti-inflammatory drug (NSAID) [337]. For these reasons, ambigol is a promising cyanobacterial anti-inflammatory compound. Nevertheless, further studies are still needed in order to describe its activities and potential unexpected side effects in-depth [338].
5.3.2. Antioxidant Activity
Oxidative stress is widely recognized to be implicated in neurodegenerative diseases [339,340], metabolic disorders [341], hypertension [342], liver diseases [343], and cardiovascular diseases [344]. Thus, antioxidant molecules are required to develop or supplement therapy for reducing the harmful effects of oxidative stress.
According to our review, only four compounds isolated from cyanobacteria show antioxidant properties. As mentioned above, this low number in comparison to cytotoxic or antimicrobial compounds might be due to the fact that this activity has been poorly tested in secondary metabolites and its testing has generally been limited to pigments or molecules implicated in light-harvesting or UV protection. Antioxidant activity has been characterized for the carotenoids, chlorophyll, the mycosporine-like amino acids (MAAs), and the phycobiliproteins such as phycocyanin (Table 10).
Carotenoids are orange pigments that are localized in the thylakoid membrane. They absorb light between 400 and 500 nm and are involved in photosynthesis by transferring energy to chlorophyll through a single-singlet energy transfer mechanism [346,349]. Five carotenoids are found in the majority of cyanobacteria: β-carotene, zeaxanthin, nostoxanthin, echinenone, and canthaxanthin [345]. In addition to their role in light harvesting, carotenoids act as potent photoprotectant molecules and show antioxidant activity through ROS scavenging [345,349] (Table 10).
Chlorophylls are the ubiquitous pigments of photosynthetic organisms. Chlorophyll a is the major isoform used by cyanobacteria with most absorbing light at 660 nm [345]. Chlorophylls are mainly involved in photosynthesis, but they have also shown antioxidant activity in vitro via radical scavenging and, on the contrary, singlet oxygen production under high light conditions, which mitigates their potential use as antioxidant therapeutics [345] (Table 10).
Mycosporine-like amino acids (MAAs) are pigments that are widely produced by cyanobacteria (Table 10) and other algae [58,345]. They absorb light in the UV-A and UV-B ranges with a maximum absorbance between 310 and 360 nm [58]. The primary function of MAAs is to protect cells from damage by absorbing UV and to dissipate energy without generating ROS [345,348]. In addition, MAAs show other unique properties. They have been demonstrated to have antioxidant activity through ROS scavenging, are able to protect skin from UV damage, and are involved in osmotic regulation, desiccation, and defense against oxidative and thermal stresses. They are also able to protect fibroblasts against UV-induced cell death [58,348]. Jain et al. (2017) stated that two products containing MAAs have been commercialized as sunscreen agents for cosmetics and for use in plastics, paints, and varnishes as a photo stabilizer [58].
Lastly, as mentioned above, phycocyanins are antioxidant molecules with the ability to scavenge ROS. In addition to their anti-inflammatory activity, this antioxidant property increases the potential of phycocyanins to be used for pharmaceutical applications [325].
5.4. Other Metabolites with Potential Beneficial Properties
To close this review on the beneficial activities demonstrated for cyanobacterial metabolites, we highlight a few other compounds that are of potential interest for various fields of application because of their specific features.
For instance, grassystatins-tasiamides constitute a depsipeptide group of related compounds isolated from Lyngbya and Symploca tropical species [350,351,352,353,354,355,356]. These metabolites have shown protease inhibitory activity against cathepsin D, cathepsin E, and the β-amyloid precursor protein-cleaving enzyme A (BACE1) for tasiamides B and F [350,351] (Table 11). In addition, these compounds have shown moderate or no cytotoxicity at concentrations higher than required for protease inhibitory activity [353,354,356]. Cathepsin D is an aspartic protease that is localized in the lysosome. This enzyme is considered a biomarker of some forms of metastatic breast cancer because of its related overexpression [236]. Cathepsin D has also been shown to promote proliferation and metastasis [236]. Cathepsin E, being also an aspartic protease, is mainly localized in immune system cells and, notably, in antigen-presenting cells [357]. Grassystatin A induces the reduction of antigen presentation in dendritic cells [352], which is correlated with the involvement of cathepsin E in this process and has led to the hypothesis that grassystatin could modulate the immune response. Alzheimer’s disease pathogenesis is mediated by the accumulation of amyloid β peptide (Aβ) in the brain. BACE1 is responsible for Aβ formation by cleaving the amyloid precursor protein (APP). As a result, BACE1 inhibitors could be promising targets for the development of new therapeutics against Alzheimer’s disease [351,358]. Considering these activities, we assume that members of the grassystatins-tasiamides family constitute promising components for the development of antiproliferative agents, immune response modulatory compounds, and therapeutics for Alzheimer’s disease treatment.
During the process of database construction, we noticed that five metabolite families showed a remarkable ability to bind to cannabinoid receptors (CB1 and CB2). These metabolites were grenadamide [359], the semiplenamides [360], serinolamide A [361], mooreamide A [362], and the columbamides [363]. CB1 and CB2 are cell membrane receptors that belong to the endocannabinoid system (ECS), which is an important part of the human physiological system. It is involved in a wide range of different processes, such as brain plasticity, memory, nociception, appetite regulation, the sleep–wake cycle, the regulation of emotions and stress, and addiction. This ubiquity for the regulation of various vital processes makes exogenous CB1 and CB2 ligands attractive as modulators of this system for the management of pain, diabetes, obesity, cancer, epilepsy, or Alzheimer’s disease, or to develop new anxiolytics [364,365]. Columbamides are the most potent CB1/CB2 ligands from cyanobacteria discovered so far (Table 11) [363]. They are linear acyl amides that have been isolated from Moorea bouillonii PNG05-198 using a genome mining approach [363]. To date, only the CB1-binding and CB2-binding activity of columbamides has been tested, and other investigations are required in order to look deeper into the activity profiles of these molecules, since they still remain promising compounds for therapeutic developments.
6. Conclusions
In this review, all available information concerning the beneficial activities of natural products of cyanobacteria was gathered. To write this review, a molecular database of the various families of metabolites isolated from cyanobacteria was constructed from the systematic analysis of 670 articles. The derived database represents 260 families of metabolites. It groups various types of information concerning the taxonomy of producing strains, the respective chemical classes, the origin strain habitats, and the tested/demonstrated activities for each member of the family, together with the related full references.
According to this review, from the above 300 different genera of cyanobacteria (referenced by the taxonomy published by Komarek et al. in 2014) [27], 90 have, so far, been reported to produce bioactive metabolites. Some of them have been shown to produce a high number of compounds, such as those from the genus Lyngbya-Moorea, which includes 85 families of metabolites isolated so far. However, the Lyngbya genus is a polyphyletic group and its taxonomy position is under revision. This number might be re-evaluated and distributed within distinctive new genera. The genomes of the producing strains are not available in the majority of cases, whereas Shih et al. (2013) demonstrated the large genomic potential of numerous cyanobacteria thanks to the biosynthetic pathways of metabolites highlighted by genome mining analyses [49]. Therefore, the potential for the discovery of new natural molecules and new biosynthetic pathways from cyanobacteria still remains very important and needs to be systematically explored.
Cyanobacterial metabolites belong to 10 chemical classes (including peptides, alkaloids, terpenes, and lipids), where most of the families of metabolites are peptide derivatives (above 50% of the families). Fourteen different types of activities can be distinguished for cyanobacterial metabolites (e.g., antimicrobial, lethality, cytotoxicity, and antioxidant). The large majority of the components are cytotoxic (110 families), whereas some activities have only been tested rarely, and their occurrence appears to be weakly demonstrated. Globally, no clear correlation has been observed between chemical classes and the specificity of the respective types of bioactivity. Further studies are needed in order to precisely understand the mechanisms of action of cyanobacterial metabolites, which potentially links bioactivity with structural features in order to support the new hypothesis on the biological function of the production of these components for organisms.
Lastly, 47 metabolites isolated from cyanobacteria that present remarkable interest for diverse fields of application were investigated further in the present literature review. For example, hassallidins, which show specific antifungal activity without antibacterial activity, and scytonemin, which has anti-inflammatory properties with no cytotoxicity, were detailed. These metabolites are potentially useful for the development of new concrete applications for cyanobacterial natural products and illustrate the interest in cyanobacteria as a prolific source of bioactive molecules.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/1660-3397/17/6/320/s1. Supplementary Data S1: Amino acid sequences of cyanopeptolin-like family; S2: Cyanobacterial metabolite database; S3: Detailed of the molecules described.
Author Contributions
J.D., C.B., A.R. and B.M. conceive the review. J.D, C.B. and B.M. write the draft of the manuscript. All authors review the manuscript.
Funding
This research was funded by the Thermes de Balaruc-les-Bains, Balrauc, France. The ANR through a PhD grant awarded to J. Demay, supported this work (grant n° 2017/0633).
Acknowledgments
We would like to thank the UMR 7245 MCAM, Muséum National d’Histoire Naturelle, Paris, France for laboratories facilities. The authors thank the anonymous referees for providing valuable suggestions that significantly improved the quality of the manuscript. We would like to thank MDPI (https://0-www-mdpi-com.brum.beds.ac.uk/authors/english) for English language editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Whitton, B.A.; Potts, M. Ecology of Cyanobacteria II: Their Diversity in Space and Time; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2012; pp. 1–13. [Google Scholar]
- Kurmayer, R.; Deng, L.; Entfellner, E. Role of toxic and bioactive secondary metabolites in colonization and bloom formation by filamentous cyanobacteria Planktothrix. Harmful Algae 2016, 54, 69–86. [Google Scholar] [CrossRef]
- Mazard, S.; Penesyan, A.; Ostrowski, M.; Paulsen, I.T.; Egan, S. Tiny Microbes with a Big Impact: The Role of Cyanobacteria and Their Metabolites in Shaping Our Future. Mar. Drugs 2016, 14, 97. [Google Scholar] [CrossRef] [PubMed]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Humbert, J.-F.; Törökné, A. New Tools for the Monitoring of Cyanobacteria in Freshwater Ecosystems. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 84–88. [Google Scholar]
- Salmaso, N.; Bernard, C.; Humbert, J.-F.; Akçaalan, R.; Albay, M.; Ballot, A.; Catherine, A.; Fastner, J.; Häggqvist, K.; Horecká, M.; et al. Basic Guide to Detection and Monitoring of Potentially Toxic Cyanobacteria. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 46–69. ISBN 0-7803-8560-8. [Google Scholar]
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef]
- Zanchett, G.; Oliveira-Filho, E.C. Cyanobacteria and cyanotoxins: From impacts on aquatic ecosystems and human health to anticarcinogenic effects. Toxins 2013, 5, 1896–1917. [Google Scholar] [CrossRef] [PubMed]
- Tidgewell, K.; Clark, B.R.; Gerwick, W.H. The natural products chemistry of cyanobacteria. In Comprehensive Natural Products II Chemistry and Biology; Elsevier: Oxford, UK, 2010; Volume 2, pp. 141–188. [Google Scholar]
- Ali Shah, S.A.; Akhter, N.; Auckloo, B.N.; Khan, I.; Lu, Y.; Wang, K.; Wu, B.; Guo, Y.W. Structural diversity, biological properties and applications of natural products from cyanobacteria. A review. Mar. Drugs 2017, 15, 354. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.; Zhang, J.; He, S.; Yan, X. New peptides isolated from marine cyanobacteria, an overview over the past decade. Mar. Drugs 2017, 15, 132. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, J.; He, S.; Yan, X. A review study on macrolides isolated from cyanobacteria. Mar. Drugs 2017, 15, 126. [Google Scholar] [CrossRef] [PubMed]
- Chlipala, G.E.; Mo, S.; Orjala, J. Chemodiversity in freshwater and terrestrial cyanobacteria—A source for drug discovery. Curr. Drug Targets 2011, 12, 1654–1673. [Google Scholar] [CrossRef]
- Sivonen, K.; Leikoski, N.; Fewer, D.P.; Jokela, J. Cyanobactins-ribosomal cyclic peptides produced by cyanobacteria. Appl. Microbiol. Biotechnol. 2010, 86, 1213–1225. [Google Scholar] [CrossRef]
- Dittmann, E.; Gugger, M.; Sivonen, K.; Fewer, D.P. Natural Product Biosynthetic Diversity and Comparative Genomics of the Cyanobacteria. Trends Microbiol. 2015, 23, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Pattanaik, B.; Lindberg, P. Terpenoids and Their Biosynthesis in Cyanobacteria. Life 2015, 5, 269–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Parihar, P.; Singh, M.; Bajguz, A.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Uncovering potential applications of cyanobacteria and algal metabolites in biology, agriculture and medicine: Current status and future prospects. Front. Microbiol. 2017, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.S.; Kumar, A.; Rai, A.N.; Singh, D.P. Cyanobacteria: A precious bio-resource in agriculture, ecosystem, and environmental sustainability. Front. Microbiol. 2016, 7, 529. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Fenner, A.M. Drug Discovery from Marine Microbes. Microb. Ecol. 2013, 65, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Shishido, T.K.; Humisto, A.; Jokela, J.; Liu, L.; Wahlsten, M.; Tamrakar, A.; Fewer, D.P.; Permi, P.; Andreote, A.P.D.; Fiore, M.F.; et al. Antifungal compounds from cyanobacteria. Mar. Drugs 2015, 13, 2124–2140. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, S.; Menakha, M. Pharmaceutical applications of cyanobacteria-A review. J. Acute Med. 2015, 5, 15–23. [Google Scholar] [CrossRef]
- Ciferri, O.; Tiboni, O. The Biochemistry and Industrial Potential of Spirulina. Annu. Rev. Microbiol. 1985, 39, 503–526. [Google Scholar] [CrossRef]
- Abdulqader, G.; Barsanti, L.; Tredici, M.R. Harvest of Arthrospira platensis from Lake Kossorom (Chad) and its household usage among the Kanembu. J. Appl. Phycol. 2000, 12, 493–498. [Google Scholar] [CrossRef]
- Lau, N.-S.; Matsui, M.; Abdullah, A.A.-A. Cyanobacteria: Photoautotrophic Microbial Factories for the Sustainable Synthesis of Industrial Products. BioMed Res. Int. 2015, 2015, 754934. [Google Scholar] [CrossRef]
- Janssen, E.M.-L. Cyanobacterial peptides beyond microcystins—A review on co-occurrence, toxicity, and challenges for risk assessment. Water Res. 2019, 151, 488–499. [Google Scholar] [CrossRef]
- Boudreau, P.D.; Byrum, T.; Liu, W.T.; Dorrestein, P.C.; Gerwick, W.H. Viequeamide A, a cytotoxic member of the kulolide superfamily of cyclic depsipeptides from a marine button cyanobacterium. J. Nat. Prod. 2012, 75, 1560–1570. [Google Scholar] [CrossRef]
- Komarek, J.; Kastovsky, J.; Mares, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Soares, A.R.; Engene, N.; Gunasekera, S.P.; Sneed, J.M.; Paul, V.J. Carriebowlinol, an Antimicrobial Tetrahydroquinolinol from an Assemblage of Marine Cyanobacteria Containing a Novel Taxon. J. Nat. Prod. 2014, 78, 534–538. [Google Scholar] [CrossRef]
- Vining, O.B.; Medina, R.A.; Mitchell, E.A.; Videau, P.; Li, D.; Serrill, J.D.; Kelly, J.X.; Gerwick, W.H.; Proteau, P.J.; Ishmael, J.E.; et al. Depsipeptide companeramides from a panamanian marine cyanobacterium associated with the coibamide producer. J. Nat. Prod. 2015, 78, 413–420. [Google Scholar] [CrossRef]
- Nogle, L.M.; Williamson, R.T.; Gerwick, W.H. Somamides A and B, two new depsipeptide analogues of dolastatin 13 from a Fijian cyanobacterial assemblage of Lyngbya majuscula and Schizothrix species. J. Nat. Prod. 2001, 64, 716–719. [Google Scholar] [CrossRef]
- Iwasaki, A.; Sumimoto, S.; Ohno, O.; Suda, S.; Suenaga, K. Kurahamide, a cyclic depsipeptide analog of dolastatin 13 from a marine cyanobacterial assemblage of Lyngbya sp. Bull. Chem. Soc. Jpn. 2014, 87, 609–613. [Google Scholar] [CrossRef]
- Harrigan, G.G.; Yoshida, W.Y.; Moore, R.E.; Nagle, D.G.; Park, P.U.; Biggs, J.; Paul, V.J.; Mooberry, S.L.; Corbett, T.H.; Valeriote, F.A. Isolation, structure determination, and biological activity of dolastatin 12 and Lyngbyastatin 1 from Lyngbya majuscula/Schizothrix calcicola cyanobacterial assemblages. J. Nat. Prod. 1998, 61, 1221–1225. [Google Scholar] [CrossRef]
- Pereira, A.; Cao, Z.; Murray, T.F.; Gerwick, W.H. Hoiamide A, a Sodium Channel Activator of Unusual Architecture from a Consortium of Two Papua New Guinea Cyanobacteria. Chem. Biol. 2009, 16, 893–906. [Google Scholar] [CrossRef] [Green Version]
- Graber, M.A.; Gerwick, W.H. Kalkipyrone, a toxic gamma-pyrone from an assemblage of the marine cyanobacteria Lyngbya majuscula and Tolypothrix sp. J. Nat. Prod. 1998, 61, 677–680. [Google Scholar] [CrossRef]
- Pereira, A.R.; Cao, Z.; Engene, N.; Soria-Mercado, I.E.; Murray, T.F.; Gerwick, W.H. Palmyrolide A, an unusually stabilized neuroactive macrolide from Palmyra Atoll cyanobacteria. Org. Lett. 2010, 12, 4490–4493. [Google Scholar] [CrossRef]
- Pereira, A.R.; Etzbach, L.; Engene, N.; Müller, R.; Gerwick, W.H. Molluscicidal metabolites from an assemblage of Palmyra Atoll cyanobacteria. J. Nat. Prod. 2011, 74, 1175–1181. [Google Scholar] [CrossRef]
- Nogle, L.M.; Gerwick, W.H. Somocystinamide A, a Novel Cytotoxic Disulfide Dimer from a Fijian Marine Cyanobacterial Mixed Assemblage. Org. Lett. 2002, 4, 1095–1098. [Google Scholar] [CrossRef]
- Williamson, R.T.; Singh, I.P.; Gerwick, W.H. Taveuniamides: New chlorinated toxins from a mixed assemblage of marine cyanobacteria. Tetrahedron 2004, 60, 7025–7033. [Google Scholar] [CrossRef]
- Harrigan, G.G.; Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Nagle, D.G.; Biggs, J.; Park, P.U.; Paul, V.J. Tumonoic acids, novel metabolites from a cyanobacterial assemblage of Lyngbya majuscula and Schizothrix calcicola. J. Nat. Prod. 1999, 62, 464–467. [Google Scholar] [CrossRef]
- Sitachitta, N.; Williamson, R.T.; Gerwick, W.H. Yanucamides A and B, two new depsipeptides from an assemblage of the marine cyanobacteria Lyngbya majuscula and Schizothrix species. J. Nat. Prod. 2000, 63, 197–200. [Google Scholar] [CrossRef]
- Osborne, N.J.; Shaw, G.R.; Webb, P.M. Health effects of recreational exposure to Moreton Bay, Australia waters during a Lyngbya majuscula bloom. Environ. Int. 2007, 33, 309–314. [Google Scholar] [CrossRef]
- Osborne, N.J.; Shaw, G.R. Dermatitis associated with exposure to a marine cyanobacterium during recreational water exposure. BMC Dermatol. 2008, 8, 5. [Google Scholar] [CrossRef]
- Engene, N.; Cameron Coates, R.; Gerwick, W.H. 16S Rrna gene heterogeneity in the filamentous marine cyanobacterial genus Lyngbya. J. Phycol. 2010, 46, 591–601. [Google Scholar] [CrossRef]
- Engene, N.; Rottacker, E.C.; Kaštovský, J.; Byrum, T.; Choi, H.; Ellisman, M.H.; Komárek, J.; Gerwick, W.H. Moorea producens gen. nov., sp. nov. and Moorea bouillonii comb. nov., tropical marine cyanobacteria rich in bioactive secondary metabolites. Int. J. Syst. Evol. Microbiol. 2012, 62, 1171–1178. [Google Scholar] [CrossRef]
- Engene, N.; Paul, V.J.; Byrum, T.; Gerwick, W.H.; Thor, A.; Ellisman, M.H. Five chemically rich species of tropical marine cyanobacteria of the genus Okeania gen. nov. (Oscillatoriales, Cyanoprokaryota). J. Phycol. 2013, 49, 1095–1106. [Google Scholar] [CrossRef]
- Komarek, J.; Zapomelova, E.; Smarda, J.; Kopecky, J.; Rejmankova, E.; Woodhouse, J.; Neilan, B.A.; Komarkova, J. Polyphasic evaluation of Limnoraphis robusta, a water-bloom forming cyanobacterium from Lake Atitlan, Guatemala, with a description of Limnoraphis gen. nov. Fottea 2013, 13, 39–52. [Google Scholar] [CrossRef]
- Mcgregor, G.B.; Sendall, B.C. Phylogeny and toxicology of Lyngbya wollei (Cyanobacteria, Oscillatoriales) from north-eastern Australia, with a description of Microseira gen. nov. J. Phycol. 2015, 51, 109–119. [Google Scholar] [CrossRef]
- Engene, N.; Choi, H.; Esquenazi, E.; Rottacker, E.C.; Ellisman, M.H.; Dorrestein, P.C.; Gerwick, W.H. Underestimated biodiversity as a major explanation for the perceived rich secondary metabolite capacity of the cyanobacterial genus Lyngbya. Environ. Microbiol. 2011, 13, 1601–1610. [Google Scholar] [CrossRef]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.D.; Fewer, D.P.; Talla, E.; Calteau, A.; Cai, F.; Tandeau de Marsac, N.; Rippka, R.; et al. Improving the coverage of the cyanobacterial phylum using diversity-driven genome sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 1053–1058. [Google Scholar] [CrossRef]
- Médigue, C.; Calteau, A.; Cruveiller, S.; Gachet, M.; Gautreau, G.; Josso, A.; Lajus, A.; Langlois, J.; Pereira, H.; Planel, R.; et al. MicroScope—An integrated resource for community expertise of gene functions and comparative analysis of microbial genomic and metabolic data. Brief. Bioinform. 2017. [Google Scholar] [CrossRef]
- Moore, R.E.; Blackman, A.J.; Cheuk, C.E.; Mynderse, J.S.; Matsumoto, G.K.; Clardy, J.; Woodard, R.W.; Craig, J.C. Absolute Stereochemistries of the Aplysiatoxins and Oscillatoxin A. J. Org. Chem. 1984, 49, 2484–2489. [Google Scholar] [CrossRef]
- Cardellina, J.H.; Marner, F.-J.; Moore, R.E. Seaweed dermatitis: Structure of Lyngbyatoxin A. Science 1979, 204, 193–195. [Google Scholar] [CrossRef]
- Ohtani, I.; Moore, R.E.; Runnegar, M.T.C. Cylindrospermopsin: A Potent Hepatotoxin from the Blue-Green Alga Cylindrospermopsis raciborskii. J. Am. Chem. Soc. 1992, 114, 7941–7942. [Google Scholar] [CrossRef]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Jokela, J.; Heinilä, L.M.P.; Shishido, T.K.; Wahlsten, M.; Fewer, D.P.; Fiore, M.F.; Wang, H.; Haapaniemi, E.; Permi, P.; Sivonen, K. Production of high amounts of hepatotoxin nodularin and new protease inhibitors pseudospumigins by the brazilian benthic Nostoc sp. CENA543. Front. Microbiol. 2017, 8, 1963. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. Antioxidant activity of carotenoids. Mol. Asp. Med. 2003, 24, 345–351. [Google Scholar] [CrossRef]
- Lanfer-Marquez, U.M.; Barros, R.M.C.; Sinnecker, P. Antioxidant activity of chlorophylls and their derivatives. Food Res. Int. 2005, 38, 885–891. [Google Scholar] [CrossRef]
- Jain, S.; Prajapat, G.; Abrar, M.; Ledwani, L.; Singh, A.; Agrawal, A. Cyanobacteria as efficient producers of mycosporine-like amino acids. J. Basic Microbiol. 2017, 57, 715–727. [Google Scholar] [CrossRef]
- Romay, C.; Armesto, J.; Remirez, D.; González, R.; Ledon, N.; García, I. Antioxidant and anti-inflammatory properties of C-phycocyanin from blue-green algae. Inflamm. Res. 1998, 47, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Balunas, M.J.; Grosso, M.F.; Villa, F.A.; Engene, N.; McPhail, K.L.; Tidgewell, K.; Pineda, L.M.; Gerwick, L.; Spadafora, C.; Kyle, D.E.; et al. Coibacins A-D, antileishmanial marine cyanobacterial polyketides with intriguing biosynthetic origins. Org. Lett. 2012, 14, 3878–3881. [Google Scholar] [CrossRef]
- Choi, H.; Mascuch, S.J.; Villa, F.A.; Byrum, T.; Teasdale, M.E.; Smith, J.E.; Preskitt, L.B.; Rowley, D.C.; Gerwick, L.; Gerwick, W.H. Honaucins A-C, potent inhibitors of inflammation and bacterial quorum sensing: Synthetic derivatives and structure-activity relationships. Chem. Biol. 2012, 19, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Kapuścik, A.; Hrouzek, P.; Kuzma, M.; Bártová, S.; Novák, P.; Jokela, J.; Pflüger, M.; Eger, A.; Hundsberger, H.; Kopecký, J. Novel Aeruginosin-865 from Nostoc sp. as a Potent Anti-inflammatory Agent. ChemBioChem 2013, 14, 2329–2337. [Google Scholar] [CrossRef]
- Shaala, L.A.; Youssef, D.T.A.; McPhail, K.L.; Elbandy, M. Malyngamide 4, a new lipopeptide from the Red Sea marine cyanobacterium Moorea producens (formerly Lyngbya majuscula). Phytochem. Lett. 2013, 6, 183–188. [Google Scholar] [CrossRef]
- Romay, C.; Gonzalez, R.; Ledon, N.; Remirez, D.; Rimbau, V. C-Phycocyanin: A Biliprotein with Antioxidant, Anti-Inflammatory and Neuroprotective Effects. Curr. Protein Pept. Sci. 2003, 4, 207–216. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sonani, R.R.; Madamwar, D. Cyanobacterial Sunscreen Scytonemin: Role in Photoprotection and Biomedical Research. Appl. Biochem. Biotechnol. 2015, 176, 1551–1563. [Google Scholar] [CrossRef]
- Prinsep, M.R.; Thomson, R.A.; West, M.L.; Wylie, B.L. Tolypodiol, an antiinflammatory diterpenoid from the cyanobacterium Tolypothrix nodosa. J. Nat. Prod. 1996, 59, 786–788. [Google Scholar] [CrossRef]
- Hayashi, T.; Hayashi, K.; Maeda, M.; Kojima, I. Calcium spirulan, an inhibitor of enveloped virus replication, from a blue-green alga Spirulina platensis. J. Nat. Prod. 1996, 59, 83–87. [Google Scholar] [CrossRef]
- Entzeroth, M.; Moore, R.E.; Niemczura, W.P.; Patterson, G.M.L.; Shoolery, J.N. O-Acetyl-O-butyryl-O-carbamoyl-O,O-dimethyl-.alpha.-cyclodextrins from the cyanophyte Tolypothrix byssoidea. J. Org. Chem. 1986, 51, 5307–5310. [Google Scholar] [CrossRef]
- Thammana, S.; Suzuki, H.; Lobkovsky, E.; Clardy, J.; Shimizu, Y. Isolation and structure assignment of an iminotetrasaccharide from a cultured filamentous cyanobacterium Anabaena sp. J. Nat. Prod. 2006, 69, 365–368. [Google Scholar] [CrossRef]
- Sueyoshi, K.; Kaneda, M.; Sumimoto, S.; Oishi, S.; Fujii, N.; Suenaga, K.; Teruya, T. Odoamide, a cytotoxic cyclodepsipeptide from the marine cyanobacterium Okeania sp. Tetrahedron 2016, 72, 5472–5478. [Google Scholar] [CrossRef]
- Mo, S.; Krunic, A.; Pegan, S.D.; Franzblau, S.G.; Orjala, J. An antimicrobial guanidine-bearing sesterterpene from the cultured cyanobacterium Scytonema sp. J. Nat. Prod. 2009, 72, 2043–2045. [Google Scholar] [CrossRef]
- Sung, S.Y.; Sin, L.T.; Tee, T.T.; Bee, S.T.; Rahmat, A.R.; Rahman, W.A.W.A.; Tan, A.C.; Vikhraman, M. Antimicrobial agents for food packaging applications. Trends Food Sci. Technol. 2013, 33, 110–123. [Google Scholar] [CrossRef]
- Abushelaibi, A.A.; Al Shamsi, M.S.; Afifi, H.S. Use of antimicrobial agents in food processing systems. Recent Pat. Food. Nutr. Agric. 2012, 4, 2–7. [Google Scholar] [CrossRef]
- Sturdy, M.; Krunic, A.; Cho, S.; Franzblau, S.; Orjala, J. Eucapsitrione, an anti- mycobacterium tuberculosis anthraquinone derivative from the cultured freshwater cyanobacterium Eucapsis sp. J. Nat. Prod. 2010, 73, 1441–1443. [Google Scholar] [CrossRef]
- Nogle, L.M.; Gerwick, W.H. Isolation of four new cyclic depsipeptides, antanapeptins A-D, and dolastatin 16 from a Madagascan collection of Lyngbya majuscula. J. Nat. Prod. 2002, 65, 21–24. [Google Scholar] [CrossRef]
- Kosalec, I.; Kremer, D.; Locatelli, M.; Epifano, F.; Genovese, S.; Carlucci, G.; Randić, M.; Zovko Končić, M. Anthraquinone profile, antioxidant and antimicrobial activity of bark extracts of Rhamnus alaternus, R. fallax, R. intermedia and R. pumila. Food Chem. 2013, 136, 335–341. [Google Scholar] [CrossRef]
- Yang, E.J.; Kim, S.H.; Lee, K.Y.; Song, K.S. Neuroprotective and anti-neuroinflammatory activities of anthraquinones isolated from Photorhabdus temperata culture broth. J. Microbiol. Biotechnol. 2018, 28, 12–21. [Google Scholar] [CrossRef]
- Park, J.G.; Kim, S.C.; Kim, Y.H.; Yang, W.S.; Kim, Y.; Hong, S.; Kim, K.H.; Yoo, B.C.; Kim, S.H.; Kim, J.H.; et al. Anti-Inflammatory and Antinociceptive Activities of Anthraquinone-2-Carboxylic Acid. Mediat. Inflamm. 2016, 2016, 1903849. [Google Scholar] [CrossRef]
- Yen, G.C.; Duh, P.D.; Chuang, D.Y. Antioxidant activity of anthraquinones and anthrone. Food Chem. 2000, 70, 437–441. [Google Scholar] [CrossRef]
- Reese, M.T.; Gulavita, N.K.; Nakao, Y.; Hamann, M.T.; Yoshida, W.Y.; Coval, S.J.; Scheuer, P.J. Kulolide: A Cytotoxic Depsipeptide from a Cephalaspidean. J. Am. Chem. Soc. 1996, 7863, 11081–11084. [Google Scholar] [CrossRef]
- Bunyajetpong, S.; Yoshida, W.Y.; Sitachitta, N.; Kaya, K. Trungapeptins A-C, cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2006, 69, 1539–1542. [Google Scholar] [CrossRef]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Lee, P.P.F.; Tan, L.T. Hantupeptins B and C, cytotoxic cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. Phytochemistry 2010, 71, 307–311. [Google Scholar] [CrossRef]
- Malloy, K.L. Structure Elucidation of Biomedically Relevant Marine Cyanobacterial Natural Products. Ph.D. Thesis, UC San Diego, San Diego, CA, USA, 2011. [Google Scholar]
- Mevers, E.; Liu, W.T.; Engene, N.; Mohimani, H.; Byrum, T.; Pevzner, P.A.; Dorrestein, P.C.; Spadafora, C.; Gerwick, W.H. Cytotoxic veraguamides, alkynyl bromide-containing cyclic depsipeptides from the marine cyanobacterium cf. Oscillatoria margaritifera. J. Nat. Prod. 2011, 74, 928–936. [Google Scholar] [CrossRef]
- Montaser, R.; Paul, V.J.; Luesch, H. Pitipeptolides C-F, antimycobacterial cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula from Guam. Phytochemistry 2011, 72, 2068–2074. [Google Scholar] [CrossRef]
- Salvador, L.A.; Biggs, J.S.; Paul, V.J.; Luesch, H. Veraguamides A-G, cyclic hexadepsipeptides from a dolastatin 16-producing cyanobacterium Symploca cf. hydnoides from Guam. J. Nat. Prod. 2011, 74, 917–927. [Google Scholar] [CrossRef]
- Almaliti, J.; Malloy, K.L.; Glukhov, E.; Spadafora, C.; Gutiérrez, M.; Gerwick, W.H. Dudawalamides A-D, Antiparasitic Cyclic Depsipeptides from the Marine Cyanobacterium Moorea producens. J. Nat. Prod. 2017, 80, 1827–1836. [Google Scholar] [CrossRef]
- Iwasaki, A.; Shiota, I.; Sumimoto, S.; Matsubara, T.; Sato, T.; Suenaga, K. Kohamamides A, B, and C, Cyclic Depsipeptides from an Okeania sp. Marine Cyanobacterium. J. Nat. Prod. 2017, 80, 1948–1952. [Google Scholar] [CrossRef]
- Luesch, H.; Pangilinan, R.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Pitipeptolides A and B, new cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2001, 64, 304–307. [Google Scholar] [CrossRef]
- Medina, R.A. Biologically Active Cyclic Depsipeptides from Marine Cyanobacteria. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 2009; 160p. [Google Scholar]
- Costa, M.S.; Rego, A.; Ramos, V.; Afonso, T.B.; Freitas, S.; Preto, M.; Lopes, V.; Vasconcelos, V.; Magalhães, C.; Leaõ, P.N. The conifer biomarkers dehydroabietic and abietic acids are widespread in Cyanobacteria. Sci. Rep. 2016, 6, 23436. [Google Scholar] [CrossRef] [PubMed]
- Acuña, U.M.; Zi, J.; Orjala, J.; Carcache de Blanco, E.J. Ambiguine I Isonitrile from Fischerella ambigua Induces Caspase-Independent Cell Death in MCF-7 Hormone Dependent Breast Cancer Cells. Int. J. Cancer Res. 2015, 49, 1655–1662. [Google Scholar]
- Becher, P.G.; Jüttner, F. Insecticidal compounds of the biofilm-forming cyanobacterium Fischerella sp. (ATCC 43239). Environ. Toxicol. 2005, 20, 363–372. [Google Scholar] [CrossRef]
- Becher, P.G.; Keller, S.; Jung, G.; Süssmuth, R.D.; Jüttner, F. Insecticidal activity of 12-epi-hapalindole J isonitrile. Phytochemistry 2007, 68, 2493–2497. [Google Scholar] [CrossRef]
- Cagide, E.; Becher, P.G.; Louzao, M.C.; Espiña, B.; Vieytes, M.R.; Jüttner, F.; Botana, L.M. Hapalindoles from the cyanobacterium Fischerella: Potential sodium channel modulators. Chem. Res. Toxicol. 2014, 27, 1696–1706. [Google Scholar] [CrossRef]
- Etchegaray, A.; Rabello, E.; Dieckmann, R.; Moon, D.H.; Fiore, M.F.; Von Döhren, H.; Tsai, S.M.; Neilan, B.A. Algicide production by the filamentous cyanobacterium Fischerella sp. CENA 19. J. Appl. Phycol. 2004, 16, 237–243. [Google Scholar] [CrossRef]
- Hillwig, M.L.; Zhu, Q.; Liu, X. Biosynthesis of Ambiguine Indole Alkaloids in Cyanobacterium Fischerella ambigua. ACS Chem. Biol. 2014, 9, 372–377. [Google Scholar] [CrossRef]
- Hillwig, M.L.; Fuhrman, H.A.; Ittiamornkul, K.; Sevco, T.J.; Kwak, D.H.; Liu, X. Identification and characterization of a welwitindolinone alkaloid biosynthetic gene cluster in the stigonematalean cyanobacterium Hapalosiphon welwitschii. ChemBioChem 2014, 15, 665–669. [Google Scholar] [CrossRef]
- Huber, U.; Moore, R.E.; Patterson, G.M.L. Isolation of a nitrile-containing indole alkaloid from the terrestrial blue-green alga Hapalosiphon delicatulus. J. Nat. Prod. 1998, 61, 1304–1306. [Google Scholar] [CrossRef]
- Kim, H.; Lantvit, D.; Hwang, C.H.; Kroll, D.J.; Swanson, S.M.; Franzblau, S.G.; Orjala, J. Indole alkaloids from two cultured cyanobacteria, Westiellopsis sp. and Fischerella muscicola. Bioorg. Med. Chem. 2012, 20, 5290–5295. [Google Scholar] [CrossRef]
- Kim, H.; Krunic, A.; Lantvit, D.; Shen, Q.; Kroll, D.J.; Swanson, S.M.; Orjala, J. Nitrile-containing fischerindoles from the cultured cyanobacterium Fischerella sp. Tetrahedron 2012, 68, 3205–3209. [Google Scholar] [CrossRef]
- Klein, D.; Daloze, D.; Braekman, J.C.; Hoffmann, L.; Demoulin, V. New hapalindoles from the cyanophyte Hapalosiphon laingii. J. Nat. Prod. 1995, 58, 1781–1785. [Google Scholar] [CrossRef]
- Koodkaew, I.; Sunohara, Y.; Matsuyama, S.; Matsumoto, H. Isolation of ambiguine D isonitrile from Hapalosiphon sp. and characterization of its phytotoxic activity. Plant Growth Regul. 2012, 68, 141–150. [Google Scholar] [CrossRef]
- Micallef, M.L.; Sharma, D.; Bunn, B.M.; Gerwick, L.; Viswanathan, R.; Moffitt, M.C. Comparative analysis of hapalindole, ambiguine and welwitindolinone gene clusters and reconstitution of indole-isonitrile biosynthesis from cyanobacteria. BMC Microbiol. 2014, 14, 213. [Google Scholar] [CrossRef]
- Micallef, M.L.; D’Agostino, P.M.; Sharma, D.; Viswanathan, R.; Moffitt, M.C. Genome mining for natural product biosynthetic gene clusters in the Subsection V cyanobacteria. BMC Genom. 2015, 16, 669. [Google Scholar] [CrossRef]
- Mo, S.; Krunic, A.; Chlipala, G.; Orjala, J. Antimicrobial ambiguine isonitriles from the cyanobacterium Fischerella ambigua. J. Nat. Prod. 2009, 72, 894–899. [Google Scholar] [CrossRef]
- Mo, S.; Krunic, A.; Santarsiero, B.D.; Franzblau, S.G.; Orjala, J. Hapalindole-related alkaloids from the cultured cyanobacterium Fischerella ambigua. Phytochemistry 2010, 71, 2116–2123. [Google Scholar] [CrossRef]
- Moore, R.E.; Cheuk, C.; Patterson, G.M.L. Hapalindoles: New Alkaloids from the Blue-Green Alga Hapalosiphon fontinalis. J. Am. Chem. Soc. 1984, 106, 6456–6457. [Google Scholar] [CrossRef]
- Moore, R.E.; Yang, X.Q.G.; Patterson, G.M.L. Fontonamide and Anhydrohapaloxindole A, Two New Alkaloids from the Blue-Green Alga Hapalosiphon fontinalis. J. Org. Chem. 1987, 52, 3773–3777. [Google Scholar] [CrossRef]
- Moore, R.E.; Cheuk, C.; Yang, X.Q.G.; Patterson, G.M.L.; Bonjouklian, R.; Smitka, T.A.; Mvnderse, J.S.; Foster, R.S.; Jones, N.D.; Swartzendruber, J.K.; et al. Hapalindoles, Antibacterial and Antimycotic Alkaloids from the Cyanophyte Hapalosiphon fontinalis. J. Org. Chem. 1987, 52, 1036–1043. [Google Scholar] [CrossRef]
- Moore, R.E.; Yang, X.Q.G.; Patterson, G.M.L.; Bonjouklian, R.; Smitka, T.A. Hapalonamides and other oxidized hapalindoles from Hapalosiphon fontinalis. Phytochemistry 1989, 28, 1565–1567. [Google Scholar] [CrossRef]
- Park, A.; Moore, R.E.; Patterson, G.M. Fischerindole L, a new isonitrile from the terrestrial blue-green alga Fischerella muscicola. Tetrahedron Lett. 1992, 33, 3257–3260. [Google Scholar] [CrossRef]
- Raveh, A.; Carmeli, S. Antimicrobial ambiguines from the cyanobacterium Fischerella sp. collected in Israel. J. Nat. Prod. 2007, 70, 196–201. [Google Scholar] [CrossRef]
- Schwartz, R.E.; Hirsch, C.F.; Pettibone, D.J.; Zink, D.L.; Springer, J.P. Unusual Cyclopropane-Containing Hapalindolinones from a Cultured Cyanobacterium. J. Org. Chem. 1987, 52, 3704–3706. [Google Scholar] [CrossRef]
- Smitka, T.A.; Bonjouklian, R.; Doolin, L.; Jones, N.D.; Deeter, J.B.; Yoshida, W.Y.; Prinsep, M.R.; Moore, R.E.; Patterson, G.M.L. Ambiguine Isonitriles, Fungicidal Hapalindole-Type Alkaloids from Three Genera of Blue-Green Algae Belonging to the Stigonemataceae. J. Org. Chem. 1992, 57, 857–861. [Google Scholar] [CrossRef]
- Stratmann, K.; Moore, R.E.; Patterson, G.M.L.; Bonjouklian, R.; Deeter, J.B.; Shaffer, S.; Smitka, T.A.; Smith, C.D. Welwitindolinones, Unusual Alkaloids from the Blue-Green Algae Hapalosiphon welwitschii and Westiella intricata. Relationship to Fischerindoles and Hapalinodoles. J. Am. Chem. Soc. 1994, 116, 9935–9942. [Google Scholar] [CrossRef]
- Walton, K.; Gantar, M.; Gibbs, P.D.L.; Schmale, M.C.; Berry, J.P. Indole alkaloids from Fischerella inhibit vertebrate development in the zebrafish (Danio rerio) embryo model. Toxins 2014, 6, 3568–3581. [Google Scholar] [CrossRef]
- Mason, C.P.; Edwards, K.R.; Carlson, R.E.; Pignatello, J.; Gleason, F.K.; Wood, J.M. Isolation of Chlorine-Containing Antibiotic from the Freshwater Cyanobacterium Scytonema hofmanni. Science 1982, 215, 400–402. [Google Scholar] [CrossRef]
- Gromov, B.V.; Vepritskiy, A.A.; Titova, N.N.; Mamkayeva, K.A.; Alexandrova, O.V. Production of the antibiotic cyanobacterin LU-1 by Nostoc linckia CALU 892 (cyanobacterium). J. Appl. Phycol. 1991, 3, 55–59. [Google Scholar] [CrossRef]
- Gleason, F.K.; Case, D.E. Activity of the natural algicide, cyanobacterin, on angiosperms. Plant Physiol. 1986, 80, 834–837. [Google Scholar] [CrossRef]
- Ishibashi, F.; Park, S.; Kusano, T.; Kuwano, K. Synthesis and algicidal activity of (+)-cyanobacterin and its stereoisomer. Biosci. Biotechnol. Biochem. 2005, 69, 391–396. [Google Scholar] [CrossRef]
- Gross, E.M.; Wolk, C.P.; Jüttner, F. Fischerellin, a new allelochemical from the freshwater cyanobacterium Fischerella muscicola. J. Phycol. 1991, 27, 686–692. [Google Scholar] [CrossRef]
- Hagmann, L.; Jüttner, F. Fischerellin A, a novel photosystem-II-inhibiting allelochemical of the cyanobacterium Fischerella muscicola with antifungal and herbicidal activity. Tetrahedron Lett. 1996, 37, 6539–6542. [Google Scholar] [CrossRef]
- Papke, U.; Gross, E.M.; Francke, W. Isolation, identification and determination of the absolute configuration of Fischerellin B. A new algicide from the freshwater cyanobacterium Fischerella muscicola (Thuret). Tetrahedron Lett. 1997, 38, 379–382. [Google Scholar] [CrossRef]
- Srivastava, V.C.; Manderson, G.J.; Bhamidimarri, R. Inhibitory metabolites production by the cyanobacterium Fischerella muscicola. Microbiol. Res. 1999, 153, 309–317. [Google Scholar] [CrossRef]
- Ogino, J.; Moore, R.E.; Patterson, G.M.L.; Smith, C.D. Dendroamides, new cyclic hexapeptides from a blue-green alga. Multidrug- resistance reversing activity of dendroamide A. J. Nat. Prod. 1996, 59, 581–586. [Google Scholar] [CrossRef]
- Todorova, A.K.; Jüttner, F.; Linden, A.; Plüiss, T.; von Philipsborn, W. Nostocyclamide: A New Macrocyclic, Thiazole-Containing Allelochemical from Nostoc sp. 31 (Cyanobacteria). J. Org. Chem. 1995, 60, 7891–7895. [Google Scholar] [CrossRef]
- Prinsep, M.R.; Moore, R.E.; Levine, I.A.; Patterson, G.M.L. Westiellamide, a bistratamide-related cyclic peptide from the blue-green alga Westiellopsis prolifica. J. Nat. Prod. 1992, 55, 140–142. [Google Scholar] [CrossRef]
- Admi, V.; Afek, U.; Carmeli, S. Raocyclamides A and B, novel cyclic hexapeptides isolated from the cyanobacterium Oscillatoria raoi. J. Nat. Prod. 1996, 59, 396–399. [Google Scholar] [CrossRef]
- Banker, R.; Carmeli, S. Tenuecyclamides A−D, Cyclic Hexapeptides from the Cyanobacterium Nostoc spongiaeforme var. tenue. J. Nat. Prod. 1998, 61, 1248–1251. [Google Scholar] [CrossRef]
- Jüttner, F.; Todorova, A.K.; Walch, N.; Von Philipsborn, W. Nostocyclamide M: A cyanobacterial cyclic peptide with allelopathic activity from Nostoc 31. Phytochemistry 2001, 57, 613–619. [Google Scholar] [CrossRef]
- Falch, B.S.; König, G.M.; Wright, A.D.; Sticher, O.; Röegger, H.; Bernardinelli, G. Ambigol A and B: New Biologically Active Polychlorinated Aromatic Compounds from the Terrestrial Blue-Green Alga Fischerella ambigua. J. Org. Chem. 1993, 58, 6570–6575. [Google Scholar] [CrossRef]
- Wright, A.D.; Papendorf, O.; König, G.M. Ambigol C and 2, 4 dichlorobenzoic acid, natural products produced by terrestrial cyanobacterium Fischerella ambigua. J. Nat. Prod. 2005, 68, 459–461. [Google Scholar] [CrossRef]
- An, T.; Kumar, T.K.S.; Wang, M.; Liu, L.; Lay, J.O.; Liyanage, R.; Berry, J.; Gantar, M.; Marks, V.; Gawley, R.E.; et al. Structures of pahayokolides A and B, cyclic peptides from a Lyngbya sp. J. Nat. Prod. 2007, 70, 730–735. [Google Scholar] [CrossRef]
- Berry, J.P.; Gantar, M.; Gawley, R.E.; Wang, M.; Rein, K.S. Pharmacology and toxicology of pahayokolide A, a bioactive metabolite from a freshwater species of Lyngbya isolated from the Florida Everglades. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2004, 139, 231–238. [Google Scholar] [CrossRef]
- Dias, F.; Antunes, J.T.; Ribeiro, T.; Azevedo, J.; Vasconcelos, V.; Leão, P.N. Cyanobacterial allelochemicals but not cyanobacterial cells markedly reduce microbial community diversity. Front. Microbiol. 2017, 8, 1495. [Google Scholar] [CrossRef]
- Leao, P.N.; Pereira, A.R.; Liu, W.-T.; Ng, J.; Pevzner, P.A.; Dorrestein, P.C.; Konig, G.M.; Vasconcelos, V.M.; Gerwick, W.H. Synergistic allelochemicals from a freshwater cyanobacterium. Proc. Natl. Acad. Sci. USA 2010, 107, 11183–11188. [Google Scholar] [CrossRef] [Green Version]
- Mehner, C.; Müller, D.; Krick, A.; Kehraus, S.; Löser, R.; Gütschow, M.; Maier, A.; Fiebig, H.H.; Brun, R.; König, G.M. A novel β-amino acid in cytotoxic peptides from the cyanobacterium Tychonema sp. Eur. J. Org. Chem. 2008, 2008, 1732–1739. [Google Scholar] [CrossRef]
- Pergament, I.; Carmeli, S. Schizotrin A; a Nwel Antimicrobial Cyclic Peptide from a Cyanobacterium. Tetrahedron Lett. 1994, 35, 8473–8476. [Google Scholar] [CrossRef]
- Ribeiro, T.; Lemos, F.; Preto, M.; Azevedo, J.; Sousa, M.L.; Leão, P.N.; Campos, A.; Linder, S.; Vitorino, R.; Vasconcelos, V.; et al. Cytotoxicity of portoamides in human cancer cells and analysis of the molecular mechanisms of action. PLoS ONE 2017, 12, e0188817. [Google Scholar] [CrossRef] [PubMed]
- Zainuddin, E.N.; Jansen, R.; Nimtz, M.; Wray, V.; Preisitsch, M.; Lalk, M.; Mundt, S. Lyngbyazothrins A-D, antimicrobial cyclic undecapeptides from the cultured cyanobacterium Lyngbya sp. J. Nat. Prod. 2009, 72, 1373–1378. [Google Scholar] [CrossRef]
- Foster, M.P.; Concepción, G.P.; Caraan, G.B.; Ireland, C.M. Bistratamides C and D. Two New Oxazole-Containing Cyclic Hexapeptides Isolated from a Philippine Lissoclinum bistratum Ascidian. J. Org. Chem. 1992, 57, 6671–6675. [Google Scholar] [CrossRef]
- Ramsay, E.E.; Hogg, P.J.; Dilda, P.J. Mitochondrial metabolism inhibitors for cancer therapy. Pharm. Res. 2011, 28, 2731–2744. [Google Scholar] [CrossRef]
- Bui, T.H.; Wray, V.; Nimtz, M.; Fossen, T.; Preisitsch, M.; Schröder, G.; Wende, K.; Heiden, S.E.; Mundt, S. Balticidins A-D, antifungal hassallidin-like lipopeptides from the Baltic Sea cyanobacterium Anabaena cylindrica Bio33. J. Nat. Prod. 2014, 77, 1287–1296. [Google Scholar] [CrossRef]
- Neuhof, T.; Schmieder, P.; Seibold, M.; Preussel, K.; Von Dö Hren, H. Hassallidin B—Second antifungal member of the Hassallidin family. Bioorg. Med. Chem. Lett. 2006, 16, 4220–4222. [Google Scholar] [CrossRef]
- Neuhof, T.; Schmieder, P.; Preussel, K.; Dieckmann, R.; Pham, H.; Bartl, F.; Von Döhren, H. Hassallidin A, a Glycosylated Lipopeptide with Antifungal Activity from the Cyanobacterium Hassallia sp. J. Nat. Prod. 2005, 68, 695–700. [Google Scholar] [CrossRef]
- Pancrace, C.; Jokela, J.; Sassoon, N.; Ganneau, C.; Desnos-Ollivier, M.; Wahlsten, M.; Humisto, A.; Calteau, A.; Bay, S.; Fewer, D.P.; et al. Rearranged Biosynthetic Gene Cluster and Synthesis of Hassallidin in Planktothrix serta PCC 8927. ACS Chem. Biol. 2017, 12, 1796–1804. [Google Scholar] [CrossRef] [PubMed]
- Vestola, J.; Shishido, T.K.; Jokela, J.; Fewer, D.P.; Aitio, O.; Permi, P.; Wahlsten, M.; Wang, H.; Rouhiainen, L.; Sivonen, K. Hassallidins, antifungal glycolipopeptides, are widespread among cyanobacteria and are the end-product of a nonribosomal pathway. Proc. Natl. Acad. Sci. USA 2014, 111, E1909–E1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.; Mevers, E.; Byrum, T.; Valeriote, F.A.; Gerwick, W.H. Lyngbyabellins K-N from two Palmyra atoll collections of the marine cyanobacterium Moorea bouillonii. Eur. J. Org. Chem. 2012, 2012, 5141–5150. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; McPhail, K.L.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Isolation and structure of five Lyngbyabellin derivatives from a Papua New Guinea collection of the marine cyanobacterium Lyngbya majuscula. Tetrahedron 2005, 61, 11723–11729. [Google Scholar] [CrossRef]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J.; Mooberry, S.L. Isolation, structure determination, and biological activity of Lyngbyabellin A from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2000, 63, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Marquez, B.L.; Watts, K.S.; Yokochi, A.; Roberts, M.A.; Verdier-Pinard, P.; Jimenez, J.I.; Hamel, E.; Scheuer, P.J.; Gerwick, W.H. Structure and absolute stereochemistry of hectochlorin, a potent stimulator of actin assembly. J. Nat. Prod. 2002, 65, 866–871. [Google Scholar] [CrossRef]
- Milligan, K.E.; Marquez, B.L.; Williamson, R.T.; Gerwick, W.H. Lyngbyabellin B, a toxic and antifungal secondary metabolite from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2000, 63, 1440–1443. [Google Scholar] [CrossRef]
- Ramaswamy, A.V.; Sorrels, C.M.; Gerwick, W.H. Cloning and Biochemical Characterization of the Hectochlorin Biosynthetic Gene Cluster from the Marine Cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2007, 70, 1977–1986. [Google Scholar] [CrossRef]
- Williams, P.G.; Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Continuing studies on the cyanobacterium Lyngbya sp.: Isolation and structure determination of 15-norlyngbyapeptin A and Lyngbyabellin D. J. Nat. Prod. 2003, 66, 595–598. [Google Scholar] [CrossRef]
- Matthew, S.; Salvador, L.A.; Schupp, P.J.; Paul, V.J.; Luesch, H. Cytotoxic halogenated macrolides and modified peptides from the apratoxin-producing marine cyanobacterium Lyngbya bouillonii from Guam. J. Nat. Prod. 2010, 73, 1544–1552. [Google Scholar] [CrossRef]
- Adiv, S.; Carmeli, S. Protease inhibitors from Microcystis aeruginosa bloom material collected from the Dalton reservoir, israel. J. Nat. Prod. 2013, 76, 2307–2315. [Google Scholar] [CrossRef]
- Gesner-Apter, S.; Carmeli, S. Three novel metabolites from a bloom of the cyanobacterium Microcystis sp. Tetrahedron 2008, 64, 6628–6634. [Google Scholar] [CrossRef]
- Lifshits, M.; Carmeli, S. Metabolites of Microcystis aeruginosa bloom material from Lake Kinneret, Israel. J. Nat. Prod. 2012, 75, 209–219. [Google Scholar] [CrossRef]
- Marner, F.J.; Moore, R.E.; Hirotsu, K.; Clardy, J. Majusculamides A and B, Two Epimeric Lipodipeptides from Lyngbya majuscula Gomont. J. Org. Chem. 1977, 42, 2815–2819. [Google Scholar] [CrossRef]
- Carter, D.C.; Moore, R.E.; Mynderse, J.S.; Niemczura, W.P.; Todd, J.S. Structure of Majusculamide C, a Cyclic Depsipeptide from Lyngbya majuscula. J. Org. Chem. 1984, 49, 236–241. [Google Scholar] [CrossRef]
- Moore, R.E.; Entzeroth, M. Majusculamide D and deoxymajusculamide D, two cytotoxins from Lyngbya majuscula. Phytochemistry 1988, 27, 3101–3103. [Google Scholar] [CrossRef]
- Koehn, F.E.; Longley, R.E.; Reed, J.K. Microcolins a and b, new immunosuppressive peptides from the blue-green alga Lyngbya majuscula. J. Nat. Prod. 1992, 55, 613–619. [Google Scholar] [CrossRef]
- Meickle, T.; Matthew, S.; Ross, C.; Luesch, H.; Paul, V. Bioassay-guided isolation and identification of desacetylmicrocolin B from Lyngbya cf. polychroa. Planta Med. 2009, 75, 1427–1430. [Google Scholar] [CrossRef]
- Simmons, T.L.; Nogle, L.M.; Media, J.; Valeriote, F.A.; Mooberry, S.L.; Gerwick, W.H. Desmethoxymajusculamide C, a cyanobacterial depsipeptide with potent cytotoxicity in both cyclic and ring-opened forms. J. Nat. Prod. 2009, 72, 1011–1016. [Google Scholar] [CrossRef]
- Tan, L.T.; Goh, B.P.L.; Tripathi, A.; Lim, M.G.; Dickinson, G.H.; Lee, S.S.C.; Teo, S.L.M. Natural antifoulants from the marine cyanobacterium Lyngbya majuscula. Biofouling 2010, 26, 685–695. [Google Scholar] [CrossRef]
- World Health Organisation (WHO). Number of Deaths Due to HIV/AIDS. Available online: https://www.who.int/gho/hiv/epidemic_status/deaths_text/en/ (accessed on 9 April 2019).
- Gupta, D.K.; Kaur, P.; Leong, S.T.; Tan, L.T.; Prinsep, M.R.; Chu, J.J.H. Anti-Chikungunya viral activities of aplysiatoxin-related compounds from the marine cyanobacterium Trichodesmium erythraeum. Mar. Drugs 2014, 12, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Chlipala, G.E.; Sturdy, M.; Krunic, A.; Lantvit, D.D.; Shen, Q.; Porter, K.; Swanson, S.M.; Orjala, J. Cylindrocyclophanes with proteasome inhibitory activity from the Cyanobacterium Nostoc sp. J. Nat. Prod. 2010, 73, 1529–1537. [Google Scholar] [CrossRef]
- Han, B.N.; Liang, T.T.; Keen, L.J.; Fan, T.T.; Zhang, X.D.; Xu, L.; Zhao, Q.; Wang, S.P.; Lin, H.W. Two Marine Cyanobacterial Aplysiatoxin Polyketides, Neo-debromoaplysiatoxin A and B, with K+Channel Inhibition Activity. Org. Lett. 2018, 20, 578–581. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Madamwar, D.; Incharoensakdi, A. Bloom Dynamics of Cyanobacteria and Their Toxins: Environmental Health Impacts and Mitigation Strategies. Front. Microbiol. 2015, 6, 1254. [Google Scholar] [CrossRef] [Green Version]
- Chlipala, G.E.; Tri, P.H.; Van Hung, N.; Krunic, A.; Shim, S.H.; Soejarto, D.D.; Orjala, J. Nhatrangins A and B, aplysiatoxin-related metabolites from the marine cyanobacterium Lyngbya majuscula from Vietnam. J. Nat. Prod. 2010, 73, 784–787. [Google Scholar] [CrossRef] [PubMed]
- Mynderse, J.S.; Moore, R.E.; Kashiwagi, M.; Norton, T.R. Antileukemia activity in the Oscillatoriaceae: Isolation of debromoaplysiatoxin from Lyngbya. Science 1977, 196, 538–540. [Google Scholar] [CrossRef]
- Boyd, M.R.; Gustafson, K.R.; McMahon, J.B.; Shoemaker, R.H.; O’Keefe, B.R.; Mori, T.; Gulakowski, R.J.; Wu, L.; Rivera, M.I.; Laurencot, C.M.; et al. Discovery of cyanovirin-N, a novel human immunodeficiency virus- inactivating protein that binds viral surface envelope glycoprotein gp120: Potential applications to microbicide development. Antimicrob. Agents Chemother. 1997, 41, 1521–1530. [Google Scholar] [CrossRef]
- Dey, B.; Lerner, D.L.; Lusso, P.; Boyd, M.R.; Elder, J.H.; Berger, E.A. Multiple antiviral activities of cyanovirin-N: Blocking of human immunodeficiency virus type 1 gp120 interaction with CD4 and coreceptor and inhibition of diverse enveloped viruses. J. Virol. 2000, 74, 4562–4569. [Google Scholar] [CrossRef]
- Matei, E.; Basu, R.; Furey, W.; Shi, J.; Calnan, C.; Aiken, C.; Gronenborn, A.M. Structure and glycan binding of a new cyanovirin-N homolog. J. Biol. Chem. 2016, 291, 18967–18976. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Hayashi, T.; Kojima, I. A Natural Sulfated Polysaccharide, Calcium Spirulan, Isolated from Spirulina platensis: In vitro and ex Vivo Evaluation of Anti-Herpes simplex Virus and Anti-Human Immunodeficiency Virus Activities. AIDS Res. Hum. Retrovir. 1996, 12, 1463–1471. [Google Scholar] [CrossRef]
- Mader, J.; Gallo, A.; Schommartz, T.; Handke, W.; Nagel, C.H.; Günther, P.; Brune, W.; Reich, K. Calcium spirulan derived from Spirulina platensis inhibits Herpes simplex virus 1 attachment to human keratinocytes and protects against herpes labialis. J. Allergy Clin. Immunol. 2016, 137, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Wijesekara, I.; Pangestuti, R.; Kim, S.K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011, 84, 14–21. [Google Scholar] [CrossRef]
- Baba, M.; Pauwels, R.; Balzarini, J.; Arnout, J.; Desmyter, J.; De Clercq, E. Mechanism of inhibitory effect of dextran sulfate and heparin on replication of human immunodeficiency virus in vitro. Proc. Natl. Acad. Sci. USA 1988, 85, 6132–6136. [Google Scholar] [CrossRef]
- World Health Organisation. Malaria. Available online: https://www.who.int/news-room/fact-sheets/detail/malaria (accessed on 10 April 2019).
- Ogawa, H.; Iwasaki, A.; Sumimoto, S.; Iwatsuki, M.; Ishiyama, A.; Hokari, R.; Otoguro, K.; Omura, S.; Suenaga, K. Isolation and Total Synthesis of Hoshinolactam, an Antitrypanosomal Lactam from a Marine Cyanobacterium. Org. Lett. 2017, 19, 890–893. [Google Scholar] [CrossRef] [PubMed]
- Davies-Coleman, M.T.; Dzeha, T.M.; Gray, C.A.; Hess, S.; Pannell, L.K.; Hendricks, D.T.; Arendse, C.E. Isolation of homodolastatin 16, a new cyclic depsipeptide from a Kenyan collection of Lyngbya majuscula. J. Nat. Prod. 2003, 66, 712–715. [Google Scholar] [CrossRef]
- Fennell, B.J.; Carolan, S.; Pettit, G.R.; Bell, A. Effects of the antimitotic natural product dolastatin 10, and related peptides, on the human malarial parasite Plasmodium falciparum. J. Antimicrob. Chemother. 2003, 51, 833–841. [Google Scholar] [CrossRef]
- Harrigan, G.G.; Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Nagle, D.G.; Paul, V.J.; Mooberry, S.L.; Corbett, T.H.; Valeriote, F.A. Symplostatin 1: A Dolastatin 10 Analogue from the Marine Cyanobacterium Symploca hydnoides. J. Nat. Prod. 1998, 61, 1075–1077. [Google Scholar] [CrossRef]
- Harrigan, G.G.; Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Nagle, D.G.; Paul, V.J. Symplostatin 2: A dolastatin 13 analogue from the marine cyanobacterium Symploca hydnoides. J. Nat. Prod. 1999, 62, 655–658. [Google Scholar] [CrossRef] [PubMed]
- Luesch, H.; Moore, R.E.; Paul, V.J.; Mooberry, S.L.; Corbett, T.H. Isolation of dolastatin 10 from the marine cyanobacterium Symploca species VP642 and total stereochemistry and biological evaluation of its analogue symplostatin 1. J. Nat. Prod. 2001, 64, 907–910. [Google Scholar] [CrossRef] [PubMed]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J.; Mooberry, S.L.; Corbett, T.H. Symplostatin 3, a new dolastatin 10 analogue from the marine cyanobacterium Symploca sp. VP452. J. Nat. Prod. 2002, 65, 16–20. [Google Scholar] [CrossRef]
- Mitchell, S.S.; Faulkner, D.J.; Rubins, K.; Bushman, F.D. Dolastatin 3 and two novel cyclic peptides from a palauan collection of Lyngbya majuscula. J. Nat. Prod. 2000, 63, 279–282. [Google Scholar] [CrossRef]
- Pettit, G.R. The Dolastatins. Prog. Chem. Org. Nat. Prod. 1997, 70, 2–79. [Google Scholar]
- Flahive, E.; Srirangam, J. The Dolastatins: Novel Antitumor Agents from Dolabella auricularia. In Anticancer Agents from Natural Products; CRC Press: Boca Raton, FL, USA, 2005; p. 600. [Google Scholar]
- Bai, R.; Pettit, G.R.; Hamel, E. Structure-activity studies with chiral isomers and with segments of the antimitotic marine peptide dolastatin 10. Biochem. Pharmacol. 1990, 40, 1859–1864. [Google Scholar] [CrossRef]
- World Health Organisation. Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 10 April 2019).
- Stewart, B.W.; Wild, C.P. (Eds.) World Cancer Report 2014; The International Agency for Research on Cancer: Lyon, France, 2014. [Google Scholar]
- Mirsalis, J.C.; Schindler-Horvat, J.; Hill, J.R.; Tomaszewski, J.E.; Donohue, S.J.; Tyson, C.A. Toxicity of dolastatin 10 in mice, rats and dogs and its clinical relevance. Cancer Chemother. Pharmacol. 1999, 44, 395–402. [Google Scholar] [CrossRef]
- Francisco, J.A.; Cerveny, C.G.; Meyer, D.L.; Mixan, B.J.; Klussman, K.; Chace, D.F.; Rejniak, S.X.; Gordon, K.A.; DeBlanc, R.; Toki, B.E.; et al. cAC10-vcMMAE, an anti-CD30-monomethyl auristatin E conjugate with potent and selective antitumor activity. Blood 2003, 102, 1458–1465. [Google Scholar] [CrossRef]
- Rose, A.A.N.; Biondini, M.; Curiel, R.; Siegel, P.M. Targeting GPNMB with glembatumumab vedotin: Current developments and future opportunities for the treatment of cancer. Pharmacol. Ther. 2017, 179, 127–141. [Google Scholar] [CrossRef]
- Barchi, J.J.; Norton, T.R.; Furusawa, E.; Patterson, G.M.L.; Moore, R.E. Identification of a cytotoxin from Tolypothrix byssoidea as tubercidin. Phytochemistry 1983, 22, 2851–2852. [Google Scholar] [CrossRef]
- Stewart, J.B.; Bornemann, V.; Chen, J.L.; Moore, R.E.; Caplan, F.R.; Karuso, H.; Larsen, L.K.; Patterson, G.M.L. Cytotoxic, fungicidal nucleosides from blue green algae belonging to the Scytonemataceae. J. Antibiot. (Tokyo) 1988, 61, 1048–1056. [Google Scholar] [CrossRef]
- Mooberry, S.L.; Stratman, K.; Moore, R.E. Tubercidin stabilizes microtubules against vinblastine-induced depolymerization, a taxol-like effect. Cancer Lett. 1995, 96, 261–266. [Google Scholar] [CrossRef]
- Altmann, K.-H. Microtubule-stabilizing agents: A growing class of important anticancer drugs. Curr. Opin. Chem. Biol. 2001, 5, 424–431. [Google Scholar] [CrossRef]
- Han, B.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Aurilides B and C, cancer cell toxins from a Papua New Guinea collection of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2006, 69, 572–575. [Google Scholar] [CrossRef]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Tan, L.T. Lagunamides A and B: Cytotoxic and antimalarial cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010, 73, 1810–1814. [Google Scholar] [CrossRef]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Chan, K.P.; Chen, D.Y.K.; Tan, L.T. Lagunamide C, a cytotoxic cyclodepsipeptide from the marine cyanobacterium Lyngbya majuscula. Phytochemistry 2011, 72, 2369–2375. [Google Scholar] [CrossRef]
- Williams, P.G.; Yoshida, W.Y.; Quon, M.K.; Moore, R.E.; Paul, V.J. The Structure of Palau’amide, a Potent Cytotoxin from a Species of the Marine Cyanobacterium Lyngbya. J. Nat. Prod. 2003, 66, 1545–1549. [Google Scholar] [CrossRef]
- Andrianasolo, E.H.; Gross, H.; Goeger, D.; Musafija-Girt, M.; McPhail, K.; Leal, R.M.; Mooberry, S.L.; Gerwick, W.H. Isolation of swinholide A and related glycosylated derivatives from two field collections of marine cyanobacteria. Org. Lett. 2005, 7, 1375–1378. [Google Scholar] [CrossRef]
- Humisto, A.; Jokela, J.; Liu, L.; Wahlsten, M.; Wang, H.; Permi, P.; Machado, J.P.; Antunes, A.; Fewer, D.P.; Sivonen, K. The swinholide biosynthesis gene cluster from a terrestrial cyanobacterium, Nostoc sp. strain UHCC 0450. Appl. Environ. Microbiol. 2018, 84, e02321-17. [Google Scholar] [CrossRef]
- Tao, Y.; Li, P.; Zhang, D.; Glukhov, E.; Gerwick, L.; Zhang, C.; Murray, T.F.; Gerwick, W.H. Samholides, Swinholide-Related Metabolites from a Marine Cyanobacterium cf. Phormidium sp. J. Org. Chem. 2018, 83, 3034–3046. [Google Scholar] [CrossRef]
- Oftedal, L.; Myhren, L.; Jokela, J.; Gausdal, G.; Sivonen, K.; Doskeland, S.O.; Herfindal, L. The lipopeptide toxins anabaenolysin A and B target biological membranes in a cholesterol-dependent manner. Biochim. Biophys. Acta Biomembr. 2012, 1818, 3000–3009. [Google Scholar] [CrossRef] [Green Version]
- Jokela, J.; Oftedal, L.; Herfindal, L.; Permi, P.; Wahlsten, M.; Døskeland, S.O.; Sivonen, K. Anabaenolysins, Novel Cytolytic Lipopeptides from Benthic Anabaena Cyanobacteria. PLoS ONE 2012, 7, e41222. [Google Scholar] [CrossRef]
- Rao, J.Y.; Li, N. Microfilament Actin Remodeling as a Potential Target for Cancer Drug Development. Curr. Cancer Drug Targets 2004, 4, 345–354. [Google Scholar] [CrossRef]
- Suenaga, K.; Mutou, T.; Shibata, T.; Itoh, T.; Fujita, T.; Takada, N.; Hayamizu, K.; Takagi, M.; Irifune, T.; Kigoshi, H.; et al. Aurilide, a cytotoxic depsipeptide from the sea hare Dolabella auricularia: Isolation, structure determination, synthesis, and biological activity. Tetrahedron 2004, 60, 8509–8527. [Google Scholar] [CrossRef]
- Nakao, Y.; Yoshida, W.Y.; Takada, Y.; Kimura, J.; Yang, L.; Mooberry, S.L.; Scheuer, P.J. Kulokekahilide-2, a cytotoxic depsipeptide from a cephalaspidean mollusk Philinopsis speciosa. J. Nat. Prod. 2004, 67, 1332–1340. [Google Scholar] [CrossRef]
- Carmeli, S.; Kashman, Y. Structure of swinholide-a, a new macrolide from the marine sponge Theonella swinhoei. Tetrahedron Lett. 1985, 26, 511–514. [Google Scholar] [CrossRef]
- Bubb, M.R.; Spector, I.; Bershadsky, A.D.; Korn, E.D. Swinholide A is a microfilament disrupting marine toxin that stabilizes actin dimers and severs actin filaments. J. Biol. Chem. 1995, 270, 3463–3466. [Google Scholar] [CrossRef]
- Golakoti, T.; Ohtani, I.; Patterson, G.M.L.; Moore, R.E.; Corbett, T.H.; Valeriote, F.A.; Demchik, L. Total Structures of Cryptophycins, Potent Antitumor Depsipeptides from the Blue-Green Alga Nostoc sp. Strain GSV 224. J. Am. Chem. Soc. 1994, 116, 4729–4737. [Google Scholar]
- Golakoti, T.; Ogino, J.; Heltzel, C.E.; Husebo, T.L.; Jensen, C.M.; Larsen, L.K.; Patterson, G.M.L.; Moore, R.E.; Mooberry, S.L.; Corbett, T.H.; et al. Structure Determination, Conformational Analysis, Chemical Stability Studies, and Antitumor Evaluation of the Cryptophycins. Isolation of 18 New Analogs from Nostoc sp. Strain GSV 224. J. Am. Chem. Soc. 1995, 117, 12030–12049. [Google Scholar] [CrossRef]
- Subbaraju, G.V.; Golakoti, T.; Patterson, G.M.L.; Moore, R.E. Three new cryptophycins from Nostoc sp. GSV 224. J. Nat. Prod. 1997, 60, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.D.; Zhang, X.; Mooberry, S.L.; Patterson, G.M.; Moore, R.E. Cryptophycin: A new antimicrotubule agent active against drug-resistant cells. Cancer Res. 1994, 54, 3779–3784. [Google Scholar]
- Stratmann, K.; Burgoyne, D.L.; Moore, R.E.; Patterson, G.M.L.; Smith, C.D. Hapalosin, a Cyanobacterial Cyclic Depsipeptide with Multidrug-Resistance Reversing Activity. J. Org. Chem. 1994, 59, 7219–7226. [Google Scholar] [CrossRef]
- Schmidt, E.W.; Nelson, J.T.; Rasko, D.A.; Sudek, S.; Eisen, J.A.; Haygood, M.G.; Ravel, J. Patellamide A and C biosynthesis by a microcin-like pathway in Prochloron didemni, the cyanobacterial symbiont of Lissoclinum patella. Proc. Natl. Acad. Sci. USA 2005, 102, 7315–7320. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.B.; Jacobs, R.S. A marine natural product, patellamide D, reverses multidrug resistance in a human leukemic cell line. Cancer Lett. 1993, 71, 97–102. [Google Scholar] [CrossRef]
- Prinsep, M.R.; Caplan, F.R.; Moore, R.E.; Patterson, G.M.L.; Smith, C.D. Tolyporphin, a Novel Multidrug Resistance Reversing Agent from the Blue-Green Alga Tolypothrix nodosa. J. Am. Chem. Soc. 1992, 114, 385–387. [Google Scholar] [CrossRef]
- Prinsep, M.R.; Patterson, G.M.L.; Larsen, L.K.; Smith, C.D. Further tolyporphins from the Blue-Green alga Tolypothrix nodosa. Tetrahedron 1995, 51, 10523–10530. [Google Scholar] [CrossRef]
- Nanayakkara, A.K.; Follit, C.A.; Chen, G.; Williams, N.S.; Vogel, P.D.; Wise, J.G. Targeted inhibitors of P-glycoprotein increase chemotherapeutic-induced mortality of multidrug resistant tumor cells. Sci. Rep. 2018, 8, 967. [Google Scholar] [CrossRef]
- Ilies, M.A.; Supuran, C.T.; Scozzafava, A. Therapeutic applications of serine protease inhibitors. Expert Opin. Ther. Pat. 2002, 12, 1181–1214. [Google Scholar] [CrossRef]
- Drag, M.; Salvesen, G.S. Emerging principles in protease-based drug discovery. Nat. Rev. Drug Discov. 2010, 9, 690–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, T.; Kalita, J.; Weldon, S.; Taggart, C. Proteases and Their Inhibitors in Chronic Obstructive Pulmonary Disease. J. Clin. Med. 2018, 7, 244. [Google Scholar] [CrossRef]
- Guay, C.; Laviolette, M.; Tremblay, G.M. Targeting serine proteases in asthma. Curr. Top. Med. Chem. 2006, 6, 393–402. [Google Scholar] [CrossRef]
- Williams, M.R.; Nakatsuji, T.; Sanford, J.A.; Vrbanac, A.F.; Gallo, R.L. Staphylococcus aureus Induces Increased Serine Protease Activity in Keratinocytes. J. Investig. Dermatol. 2017, 137, 377–384. [Google Scholar] [CrossRef]
- Lee, H.; Ren, J.; Nocadello, S.; Rice, A.J.; Ojeda, I.; Light, S.; Minasov, G.; Vargas, J.; Nagarathnam, D.; Anderson, W.F.; et al. Identification of novel small molecule inhibitors against NS2B/NS3 serine protease from Zika virus. Antivir. Res. 2017, 139, 49–58. [Google Scholar] [CrossRef]
- Adams, J. The development of proteasome inhibitors as anticancer drugs. Cancer Cell 2004, 5, 417–421. [Google Scholar] [CrossRef] [Green Version]
- Koblinski, J.E.; Ahram, M.; Sloane, B.F. Unraveling the role of proteases in cancer. Clin. Chim. Acta 2000, 291, 113–135. [Google Scholar] [CrossRef]
- Kos, J.; Mitrović, A.; Mirković, B. The current stage of cathepsin B inhibitors as potential anticancer agents. Future Med. Chem. 2014, 6, 1355–1371. [Google Scholar] [CrossRef]
- Bian, B.; Mongrain, S.; Cagnol, S.; Langlois, M.-J.; Boulanger, J.; Bernatchez, G.; Carrier, J.C.; Boudreau, F.; Rivard, N. Cathepsin B promotes colorectal tumorigenesis, cell invasion, and metastasis. Mol. Carcinog. 2016, 55, 671–687. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.; Platet, N.; Liaudet, E.; Laurent, V.; Derocq, D.; Brouillet, J.P.; Rochefort, H. Biological and clinical significance of cathepsin D in breast cancer metastasis. Stem Cells 1996, 14, 642–650. [Google Scholar] [CrossRef]
- Lankelma, J.M.; Voorend, D.M.; Barwari, T.; Koetsveld, J.; Van der Spek, A.H.; De Porto, A.P.N.A.; Van Rooijen, G.; Van Noorden, C.J.F. Cathepsin L, target in cancer treatment? Life Sci. 2010, 86, 225–233. [Google Scholar] [CrossRef]
- Leto, G.; Sepporta, M.V.; Crescimanno, M.; Flandina, C.; Tumminello, F.M. Cathepsin L in metastatic bone disease: Therapeutic implications. Biol. Chem. 2010, 391, 655–664. [Google Scholar] [CrossRef]
- Fujii, K.; Sivonen, K.; Adachi, K.; Noguchi, K.; Sano, H.; Hirayama, K.; Suzuki, M.; Harada, K. Comparative study of toxic and non-toxic cyanobacterial products: Novel peptides from toxic Nodularia spumigena AV1. Tetrahedron Lett. 1997, 38, 5525–5528. [Google Scholar] [CrossRef]
- Anas, A.R.J.; Kisugi, T.; Umezawa, T.; Matsuda, F.; Campitelli, M.R.; Quinn, R.J.; Okino, T. Thrombin Inhibitors from the Freshwater Cyanobacterium Anabaena compacta. J. Nat. Prod. 2012, 75, 1546–1552. [Google Scholar] [CrossRef]
- Fewer, D.P.; Jokela, J.; Rouhiainen, L.; Wahlsten, M.; Koskenniemi, K.; Stal, L.J.; Sivonen, K. The non-ribosomal assembly and frequent occurrence of the protease inhibitors spumigins in the bloom-forming cyanobacterium Nodularia spumigena. Mol. Microbiol. 2009, 73, 924–937. [Google Scholar] [CrossRef]
- Gondi, C.S.; Rao, J.S. Cathepsin B as a cancer target. Expert Opin. Ther. Targets 2013, 17, 281–291. [Google Scholar] [CrossRef] [Green Version]
- Adiv, S.; Ahronov-Nadborny, R.; Carmeli, S. New aeruginazoles, a group of thiazole-containing cyclic peptides from Microcystis aeruginosa blooms. Tetrahedron 2012, 68, 1376–1383. [Google Scholar] [CrossRef]
- Al-Awadhi, F.H.; Salvador, L.A.; Law, B.K.; Paul, V.J.; Luesch, H. Kempopeptin C, a novel marine-derived serine protease inhibitor targeting invasive breast cancer. Mar. Drugs 2017, 15, 290. [Google Scholar] [CrossRef]
- Banker, R.; Carmeli, S. Inhibitors of serine proteases from a waterbloom of the cyanobacterium Microcystis sp. Tetrahedron 1999, 55, 10835–10844. [Google Scholar] [CrossRef]
- Beresovsky, D.; Hadas, O.; Livne, A.; Sukenik, A.; Kaplan, A.; Carmeli, S. Toxins and Biologically Active Secondary Metabolites of Microcystis sp. isolated from Lake Kinneret. Isr. J. Chem. 2006, 46, 79–87. [Google Scholar] [CrossRef]
- Blom, J.F.; Bister, B.; Bischoff, D.; Nicholson, G.; Jung, G.; Süssmuth, R.D.; Jüttner, F. Oscillapeptin J, a new grazer toxin of the freshwater cyanobacterium Planktothrix rubescens. J. Nat. Prod. 2003, 66, 431–434. [Google Scholar] [CrossRef]
- Bonjouklian, R.; Smitka, T.A.; Hunt, A.H.; Occolowitz, J.L.; Perun, T.J.; Doolin, L.; Stevenson, S.; Knauss, L.; Wijayaratne, R.; Szewczyk, S.; et al. A90720A, a serine protease inhibitor isolated from a terrestrial blue-green alga Microchaete loktakensis. Tetrahedron 1996, 52, 395–404. [Google Scholar] [CrossRef]
- Choi, H.; Oh, S.K.; Yih, W.; Chin, J.; Kang, H.; Rho, J.-R. Cyanopeptoline CB071: A cyclic depsipeptide isolated from the freshwater cyanobacterium Aphanocapsa sp. Chem. Pharm. Bull. (Tokyo) 2008, 56, 1191–1193. [Google Scholar] [CrossRef]
- Elkobi-Peer, S.; Carmeli, S. New prenylated aeruginosin, microphycin, anabaenopeptin and micropeptin analogues from a Microcystis bloom material collected in Kibbutz Kfar Blum, Israel. Mar. Drugs 2015, 13, 2347–2375. [Google Scholar] [CrossRef]
- Fujii, K.; Sivonen, K.; Naganawa, E.; Harada, K. ichi Non-toxic peptides from toxic cyanobacteria, Oscillatoria agardhii. Tetrahedron 2000, 56, 725–733. [Google Scholar] [CrossRef]
- Gallegos, D.A.; Saurí, J.; Cohen, R.D.; Wan, X.; Videau, P.; Vallota-Eastman, A.O.; Shaala, L.A.; Youssef, D.T.A.; Williamson, R.T.; Martin, G.E.; et al. Jizanpeptins, Cyanobacterial Protease Inhibitors from a Symploca sp. Cyanobacterium Collected in the Red Sea. J. Nat. Prod. 2018, 81, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Grach-Pogrebinsky, O.; Sedmak, B.; Carmeli, S. Protease inhibitors from a Slovenian Lake Bled toxic waterbloom of the cyanobacterium Planktothrix rubescens. Tetrahedron 2003, 59, 8329–8336. [Google Scholar] [CrossRef]
- Gunasekera, S.P.; Miller, M.W.; Kwan, J.C.; Luesch, H.; Paul, V.J. Molassamide, a depsipeptide serine protease inhibitor from the marine cyanobacterium Dichothrix utahensis. J. Nat. Prod. 2010, 73, 459–462. [Google Scholar] [CrossRef]
- Harada, K.I.; Mayumi, T.; Shimada, T.; Fujii, K.; Kondo, F.; Park, H.D.; Watanabe, M.F. Co-production of microcystins and aeruginopeptins by natural cyanobacterial bloom. Environ. Toxicol. 2001, 16, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Matsuda, H.; Murakami, M.; Yamaguchi, K. Micropeptins 478-A and -B, plasmin inhibitors from the cyanobacterium Microcystis aeruginosa. J. Nat. Prod. 1997, 60, 184–187. [Google Scholar] [CrossRef]
- Ishida, K.; Matsuda, H.; Murakami, M. Micropeptins 88-A to 88-F, chymotrypsin inhibitors from the cyanobacterium Microcystis aeruginosa (NIES-88). Tetrahedron 1998, 54, 5545–5556. [Google Scholar] [CrossRef]
- Ishida, K.; Murakami, M.; Matsuda, H.; Yamaguchi, K. Micropeptin 90, a plasmin and trypsin inhibitor from the blue-green alga Microcystis aeruginosa (NIES-90). Tetrahedron Lett. 1995, 36, 3535–3538. [Google Scholar] [CrossRef]
- Itou, Y.; Ishida, K.; Shin, H.J.; Murakami, M. Oscillapeptins A to F, serine protease inhibitors from the three strains of Oscillatoria agardhii. Tetrahedron 1999, 55, 6871–6882. [Google Scholar] [CrossRef]
- Jakobi, C.; Oberer, L.; Quiquerez, C.; König, W.A.; Weckesser, J. Cyanopeptolin S, a sulfate-containing depsipeptide from a water bloom of Microcystis sp. FEMS Microbiol. Lett. 1995, 129, 129–133. [Google Scholar]
- Kang, H.S.; Krunic, A.; Orjala, J. Stigonemapeptin, an Ahp-containing depsipeptide with elastase inhibitory activity from the bloom-forming freshwater cyanobacterium Stigonema sp. J. Nat. Prod. 2012, 75, 807–811. [Google Scholar] [CrossRef]
- Kaya, K.; Sano, T.; Beattie, K.A.; Codd, G.A. Nostocyclin, a novel 3-amino-6-hydroxy-2-piperidone-containing cyclic depsipeptide from the cyanobacterium Nostoc sp. Tetrahedron Lett. 1996, 37, 6725–6728. [Google Scholar] [CrossRef]
- Kodani, S.; Suzuki, S.; Ishida, K.; Murakami, M. Five new cyanobacterial peptides from water bloom materials of Lake Teganuma (Japan). FEMS Microbiol. Lett. 1999, 178, 343–348. [Google Scholar] [CrossRef]
- Kwan, J.C.; Taori, K.; Paul, V.J.; Luesch, H. Lyngbyastatins 8-10, elastase inhibitors with cyclic depsipeptide scaffolds isolated from the marine cyanobacterium Lyngbya semiplena. Mar. Drugs 2009, 7, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Lifshits, M.; Zafrir-Ilan, E.; Raveh, A.; Carmeli, S. Protease inhibitors from three fishpond water blooms of Microcystis spp. Tetrahedron 2011, 67, 4017–4024. [Google Scholar] [CrossRef]
- Linington, R.G.; Edwards, D.J.; Shuman, C.F.; McPhail, K.L.; Matainaho, T.; Gerwick, W.H. Symplocamide A, a potent cytotoxin and chymotrypsin inhibitor from the marine cyanobacterium Symploca sp. J. Nat. Prod. 2008, 71, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Lodin-Friedman, A.; Carmeli, S. Metabolites from Microcystis aeruginosa bloom material collected at a water reservoir near Kibbutz Hafetz Haim, Israel. J. Nat. Prod. 2013, 76, 1196–1200. [Google Scholar] [CrossRef]
- Martin, C.; Oberer, L.; Buschdtt, M.; Weckesser, J. Cyanopeptolins, new depsipeptides from the cyanobacterium Microcystis sp. PCC 7806. J. Antibiot. 1993, 46, 1550–1556. [Google Scholar] [CrossRef]
- Matern, U.; Oberer, L.; Falchetto, R.A.; Erhard, M.; König, W.A.; Herdman, M.; Weckesser, J. Scyptolin A and B, cyclic depsipeptides from axenic cultures of Scytonema hofmanni PCC 7110. Phytochemistry 2001, 58, 1087–1095. [Google Scholar] [CrossRef]
- Matern, U.; Oberer, L.; Erhard, M.; Herdmand, M.; Weckesser, J. Hofmannolin, a cyanopeptolin from Scytonema hofmanni PCC 7110. Phytochemistry 2003, 64, 1061–1067. [Google Scholar] [CrossRef]
- Matthew, S.; Ross, C.; Paul, V.J.; Luesch, H. Pompanopeptins A and B, new cyclic peptides from the marine cyanobacterium Lyngbya confervoides. Tetrahedron 2008, 64, 4081–4089. [Google Scholar] [CrossRef]
- Taori, K.; Matthew, S.; Rocca, J.R.; Paul, V.J.; Luesch, H. Lyngbyastatins 5-7, potent elastase inhibitors from Floridian marine cyanobacteria, Lyngbya spp. J. Nat. Prod. 2007, 70, 1593–1600. [Google Scholar] [CrossRef] [PubMed]
- Mehner, C.; Müller, D.; Kehraus, S.; Hautmann, S.; Gütschow, M.; König, G.M. New Peptolides from the Cyanobacterium Nostoc insulare as Selective and Potent Inhibitors of Human Leukocyte Elastase. ChemBioChem 2008, 9, 2692–2703. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Kodani, S.; Ishida, K.; Matsuda, H.; Yamaguchi, K. Micropeptin 103, a chymotrypsin inhibitor from the cyanobacterium Microcystis viridis (NIES-103). Tetrahedron Lett. 1997, 38, 3035–3038. [Google Scholar] [CrossRef]
- Okano, T.; Sano, T.; Kaya, K. Micropeptin T-20, a novel phosphate-containing cyclic depsipeptide from the cyanobacterium Microcystis aeruginosa. Tetrahedron Lett. 1999, 40, 2379–2382. [Google Scholar] [CrossRef]
- Okino, T.; Murakami, M.; Haraguchi, R.; Munekata, H.; Matsuda, H.; Yamaguchi, K. Micropeptins A and B, plasmin and trypsin inhibitors from the blue-green alga Microcystis aeruginosa. Tetrahedron Lett. 1993, 34, 8131–8134. [Google Scholar] [CrossRef]
- Okino, T.; Qi, S.; Matsuda, H.; Murakami, M.; Yamaguchi, K. Nostopeptins A and B, elastase inhibitors from the cyanobacterium Nostoc minutum. J. Nat. Prod. 1997, 60, 158–161. [Google Scholar] [CrossRef]
- Okumura, H.S.; Philmus, B.; Portmann, C.; Hemscheidt, T.K. Homotyrosine-containing cyanopeptolins 880 and 960 and anabaenopeptins 908 and 915 from Planktothrix agardhii CYA 126/8. J. Nat. Prod. 2009, 72, 172–176. [Google Scholar] [CrossRef]
- Ploutno, A.; Carmeli, S. Modified peptides from a water bloom of the cyanobacterium Nostoc sp. Tetrahedron 2002, 58, 9949–9957. [Google Scholar] [CrossRef]
- Ploutno, A.; Shoshan, M.; Carmeli, S. Three novel protease inhibitors from a natural bloom of the cyanobacterium Microcystis aeruginosa. J. Nat. Prod. 2002, 65, 973–978. [Google Scholar] [CrossRef]
- Reshef, V.; Carmeli, S. Protease inhibitors from a water bloom of the cyanobacterium Microcystis aeruginosa. Tetrahedron 2001, 57, 2885–2894. [Google Scholar] [CrossRef]
- Sano, T.; Kaya, K. Oscillamide-Y, a Chymotrypsin Inhibitor from Toxic Oscillatoria agardhii. Tetrahedron Lett. 1995, 36, 5933–5936. [Google Scholar] [CrossRef]
- Sano, T.; Kaya, K. Oscillapeptin G, a tyrosinase inhibitor from toxic Oscillatoria agardhii. J. Nat. Prod. 1996, 59, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Murakami, M.; Matsuda, H.; Ishida, K.; Yamaguchi, K. Oscillapeptin, an elastase and chymotrypsin inhibitor from the cyanobacterium Oscillatoria agardhii (NIES-204). Tetrahedron Lett. 1995, 36, 5235–5238. [Google Scholar] [CrossRef]
- Stolze, S.C.; Deu, E.; Kaschani, F.; Li, N.; Florea, B.I.; Richau, K.H.; Colby, T.; Van Der Hoorn, R.A.L.; Overkleeft, H.S.; Bogyo, M.; et al. The antimalarial natural product symplostatin 4 is a nanomolar inhibitor of the food vacuole falcipains. Chem. Biol. 2012, 19, 1546–1555. [Google Scholar] [CrossRef]
- Taori, K.; Paul, V.J.; Luesch, H. Kempopeptins A and B, serine protease inhibitors with different selectivity profiles from a marine cyanobacterium, Lyngbya sp. J. Nat. Prod. 2008, 71, 1625–1629. [Google Scholar] [CrossRef]
- Thorskov Bladt, T.; Kalifa-Aviv, S.; Ostenfeld Larsen, T.; Carmeli, S. Micropeptins from Microcystis sp. collected in Kabul Reservoir, Israel. Tetrahedron 2014, 70, 936–943. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Painuly, P.; Young, K.A.; Yang, X.; Shimizu, Y.; Cornell, L. Microcystilide A: A Novel Cell-Differentiation-Promoting Depsipeptide from Microcystis aeruginosa NO-15-1840. J. Am. Chem. Soc. 1993, 115, 11046–11047. [Google Scholar] [CrossRef]
- Vegman, M.; Carmeli, S. Eight micropeptins from a Microcystis spp. bloom collected from a fishpond near Kibbutz Lehavot HaBashan, Israel. Tetrahedron 2013, 69, 10108–10115. [Google Scholar] [CrossRef]
- Von Elert, E.; Oberer, L.; Merkel, P.; Huhn, T.; Blom, J.F. Cyanopeptolin 954, a chlorine-containing chymotrypsin inhibitor of Microcystis aeruginosa NIVA Cya 43. J. Nat. Prod. 2005, 68, 1324–1327. [Google Scholar] [CrossRef]
- Zafrir-Ilan, E.; Carmeli, S. Eight novel serine proteases inhibitors from a water bloom of the cyanobacterium Microcystis sp. Tetrahedron 2010, 66, 9194–9202. [Google Scholar] [CrossRef]
- Fujii, K.; Sivonen, K.; Nakano, T.; Harada, K.I. Structural elucidation of cyanobacterial peptides encoded by peptide synthetase gene in Anabaena species. Tetrahedron 2002, 58, 6863–6871. [Google Scholar] [CrossRef]
- Pereira, A.R.; Kale, A.J.; Fenley, A.T.; Byrum, T.; Debonsi, H.M.; Gilson, M.K.; Valeriote, F.A.; Moore, B.S.; Gerwick, W.H. The Carmaphycins: New Proteasome Inhibitors Exhibiting an α,β-Epoxyketone Warhead from a Marine Cyanobacterium. ChemBioChem 2012, 13, 810–817. [Google Scholar] [CrossRef] [PubMed]
- LaMonte, G.M.; Almaliti, J.; Bibo-Verdugo, B.; Keller, L.; Zou, B.Y.; Yang, J.; Antonova-Koch, Y.; Orjuela-Sanchez, P.; Boyle, C.A.; Vigil, E.; et al. Development of a Potent Inhibitor of the Plasmodium Proteasome with Reduced Mammalian Toxicity. J. Med. Chem. 2017, 60, 6721–6732. [Google Scholar] [CrossRef] [PubMed]
- Hirota, M.; Ohmuraya, M.; Baba, H. The role of trypsin, trypsin inhibitor, and trypsin receptor in the onset and aggravation of pancreatitis. J. Gastroenterol. 2006, 41, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Hilpert, K.; Ackermann, J.; Banner, D.W.; Gast, A.; Gubernator, K.; Hadvary, P.; Labler, L.; Mueller, K.; Schmid, G. Design and Synthesis of Potent and Highly Selective Thrombin Inhibitors. J. Med. Chem. 1994, 37, 3889–3901. [Google Scholar] [CrossRef] [PubMed]
- Bui, H.T.N.; Jansen, R.; Pham, H.T.L.; Mundt, S. Carbamidocyclophanes A-E, chlorinated paracyclophanes with cytotoxic and antibiotic activity from the Vietnamese cyanobacterium Nostoc sp. J. Nat. Prod. 2007, 70, 499–503. [Google Scholar] [CrossRef]
- Chen, J.L.; Moore, R.E.; Patterson, G.M.L. Structures of Nostocyclophanes A-D. J. Org. Chem. 1991, 56, 4360–4364. [Google Scholar] [CrossRef]
- Kang, H.S.; Santarsiero, B.D.; Kim, H.; Krunic, A.; Shen, Q.; Swanson, S.M.; Chai, H.; Kinghorn, A.D.; Orjala, J. Merocyclophanes A and B, antiproliferative cyclophanes from the cultured terrestrial Cyanobacterium Nostoc sp. Phytochemistry 2012, 79, 109–115. [Google Scholar] [CrossRef]
- Luo, S.; Kang, H.S.; Krunic, A.; Chlipala, G.E.; Cai, G.; Chen, W.L.; Franzblau, S.G.; Swanson, S.M.; Orjala, J. Carbamidocyclophanes F and G with anti-Mycobacterium tuberculosis activity from the cultured freshwater cyanobacterium Nostoc sp. Tetrahedron Lett. 2014, 55, 686–689. [Google Scholar] [CrossRef]
- Moore, B.S.; Chen, J.-L.; Patterson, G.M.L.; Moore, R.E. Structures of Cylindrocyclophanes A-F. Tetrahedron 1992, 48, 3001–3006. [Google Scholar] [CrossRef]
- Nakamura, H.; Hamer, H.A.; Sirasani, G.; Balskus, E.P. Cylindrocyclophane biosynthesis involves functionalization of an unactivated carbon center. J. Am. Chem. Soc. 2012, 134, 18518–18521. [Google Scholar] [CrossRef]
- Preisitsch, M.; Harmrolfs, K.; Pham, H.T.; Heiden, S.E.; Füssel, A.; Wiesner, C.; Pretsch, A.; Swiatecka-Hagenbruch, M.; Niedermeyer, T.H.; Müller, R.; et al. Anti-MRSA-acting carbamidocyclophanes H-L from the Vietnamese cyanobacterium Nostoc sp. CAVN2. J. Antibiot. (Tokyo) 2015, 68, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Golakoti, T.; Yoshida, W.Y.; Chaganty, S.; Moore, R.E. Isolation and Structure Determination of Nostocyclopeptides A1 and A2 from the Terrestrial Cyanobacterium Nostoc sp. ATCC53789. J. Nat. Prod. 2001, 64, 54–59. [Google Scholar] [CrossRef]
- Krunic, A.; Vallat, A.; Mo, S.; Lantvit, D.D.; Swanson, S.M.; Orjala, J. Scytonemides A and B, cyclic peptides with 20S proteasome inhibitory activity from the cultured cyanobacterium Scytonema hofmanii. J. Nat. Prod. 2010, 73, 1927–1932. [Google Scholar] [CrossRef]
- Shim, S.H.; Chlipala, G.; Orjala, J. Isolation and structure determination of a proteasome inhibitory metabolite from a culture of Scytonema hofmanni. J. Microbiol. Biotechnol. 2008, 18, 1655–1658. [Google Scholar]
- Kobayashi, A.; Kaiiyama, S.; Inawaka, K.; Kanzaki, H.; Kawazu, K. Nostodione A, a Novel Mitotic Spindle Poison* from a Blue-Green Alga Nostoc commune. Z. Nat. C 1994, 49, 464–470. [Google Scholar] [CrossRef]
- Almaliti, J.; Miller, B.; Pietraszkiewicz, H.; Glukhov, E.; Naman, C.B.; Kline, T.; Hanson, J.; Li, X.; Zhou, S.; Valeriote, F.A.; et al. Exploration of the carmaphycins as payloads in antibody drug conjugate anticancer agents. Eur. J. Med. Chem. 2018, 161, 416–432. [Google Scholar] [CrossRef] [PubMed]
- Salvador-Reyes, L.A.; Luesch, H. Biological targets and mechanisms of action of natural products from marine cyanobacteria. Nat. Prod. Rep. 2015, 32, 478–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolden, J.E.; Peart, M.J.; Johnstone, R.W. Anticancer activities of histone deacetylase inhibitors. Nat. Rev. Drug Discov. 2006, 5, 769–784. [Google Scholar] [CrossRef] [PubMed]
- Taori, K.; Paul, V.J.; Luesch, H. Structure and activity of largazole, a potent antiproliferative agent from the Floridian marine cyanobacterium Symploca sp. J. Am. Chem. Soc. 2008, 130, 1806–1807. [Google Scholar] [CrossRef]
- Liu, Y.; Salvador, L.A.; Byeon, S.; Ying, Y.; Kwan, J.C.; Law, B.K.; Hong, J.; Luesch, H. Anticolon Cancer Activity of Largazole, a Marine-Derived Tunable Histone Deacetylase Inhibitor. J. Pharmacol. Exp. Ther. 2010, 335, 351–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.C.; Wen, Z.S.; Qiu, Y.T.; Chen, X.Q.; Chen, H.B.; Wei, M.M.; Liu, Z.; Jiang, S.; Zhou, G.B. Largazole arrests cell cycle at g1 phase and triggers proteasomal degradation of E2F1 in lung cancer cells. ACS Med. Chem. Lett. 2013, 4, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Jiang, S.; Chen, J.; Ren, X.; Jin, J.; Su, S.B. Largazole, an inhibitor of class I histone deacetylases, attenuates inflammatory corneal neovascularization. Eur. J. Pharmacol. 2014, 740, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Poli, G.; Di Fabio, R.; Ferrante, L.; Summa, V.; Botta, M. Largazole Analogues as Histone Deacetylase Inhibitors and Anticancer Agents: An Overview of Structure–Activity Relationships. ChemMedChem 2017, 12, 1917–1926. [Google Scholar] [CrossRef]
- Ungermannova, D.; Parker, S.J.; Nasveschuk, C.G.; Wang, W.; Quade, B.; Zhang, G.; Kuchta, R.D.; Phillips, A.J.; Liu, X. Largazole and its derivatives selectively inhibit ubiquitin activating enzyme (E1). PLoS ONE 2012, 7, e29208. [Google Scholar] [CrossRef] [PubMed]
- Pavlik, C.M.; Wong, C.Y.B.; Ononye, S.; Lopez, D.D.; Engene, N.; McPhail, K.L.; Gerwick, W.H.; Balunas, M.J. Santacruzamate A, a potent and selective histone deacetylase inhibitor from the Panamanian marine cyanobacterium cf. Symploca sp. J. Nat. Prod. 2013, 76, 2026–2033. [Google Scholar] [CrossRef]
- Hanke, T.; Merk, D.; Steinhilber, D.; Geisslinger, G.; Schubert-Zsilavecz, M. Small molecules with anti-inflammatory properties in clinical development. Pharmacol. Ther. 2016, 157, 163–187. [Google Scholar] [CrossRef]
- Ridker, P.M.; Lüscher, T.F. Anti-inflammatory therapies for cardiovascular disease. Eur. Heart J. 2014, 35, 1782–1791. [Google Scholar] [CrossRef]
- Rocha, N.P.; De Miranda, A.S.; Teixeira, A.L. Insights into neuroinflammation in Parkinson’s disease: From biomarkers to anti-inflammatory based therapies. BioMed Res. Int. 2015, 2015, 628192. [Google Scholar] [CrossRef]
- Ishida, K.; Okita, Y.; Matsuda, H.; Okino, T.; Murakami, M. Aeruginosins, protease inhibitors from the cyanobacterium Microcystis aeruginosa. Tetrahedron 1999, 55, 10971–10988. [Google Scholar] [CrossRef]
- Ishida, K.; Christiansen, G.; Yoshida, W.Y.; Kurmayer, R.; Welker, M.; Valls, N.; Bonjoch, J.; Hertweck, C.; Börner, T.; Hemscheidt, T.; et al. Biosynthesis and Structure of Aeruginoside 126A and 126B, Cyanobacterial Peptide Glycosides Bearing a 2-Carboxy-6-Hydroxyoctahydroindole Moiety. Chem. Biol. 2007, 14, 565–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, H.; Okino, T.; Murakami, M.; Yamaguchi, K. Aeruginosins 102-A and B, New Thrombin Inhibitors from the Cyanobacterium Microcystis viridis (NIES-102). Tetrahedron 1996, 52, 14501–14506. [Google Scholar] [CrossRef]
- Benedetti, S.; Benvenuti, F.; Pagliarani, S.; Francogli, S.; Scoglio, S.; Canestrari, F. Antioxidant properties of a novel phycocyanin extract from the blue-green alga Aphanizomenon flos-aquae. Life Sci. 2004, 75, 2353–2362. [Google Scholar] [CrossRef] [PubMed]
- Kuddus, M.; Singh, P.; Thomas, G.; Al-Hazimi, A. Recent developments in production and biotechnological applications of c-phycocyanin. BioMed Res. Int. 2013, 2013, 742859. [Google Scholar] [CrossRef]
- Patel, A.; Mishra, S.; Ghosh, P.K. Antioxidant potential of C-phycocyanin isolated from cyanobacterial species Lyngbya, Phormidium and Spirulina spp. Indian J. Biochem. Biophys. 2006, 43, 25–31. [Google Scholar] [PubMed]
- Garcia-Pichel, F.; Castenholz, R.W. Characterization and biological implications of scytonemin, a cyanobacterial sheath pigment. J. Phycol. 1991, 27, 395–409. [Google Scholar] [CrossRef]
- Proteau, P.J.; Gerwick, W.H.; Garcia-Pichel, F.; Castenholz, R. The structure of scytonemin, an ultraviolet sunscreen pigment from the sheaths of cyanobacteria. Experientia 1993, 49, 825–829. [Google Scholar] [CrossRef]
- Soule, T.; Stout, V.; Swingley, W.D.; Meeks, J.C.; Garcia-Pichel, F. Molecular genetics and genomic analysis of scytonemin biosynthesis in Nostoc punctiforme ATCC 29133. J. Bacteriol. 2007, 189, 4465–4472. [Google Scholar] [CrossRef]
- Stevenson, C.S.; Capper, E.A.; Roshak, A.K.; Marquez, B.; Grace, K.; Gerwick, W.H.; Jacobs, R.S.; Marshall, L.A. Scytonemin-a marine natural product inhibitor of kinases key in hyperproliferative inflammatory diseases. Inflamm. Res. 2002, 51, 112–114. [Google Scholar] [CrossRef]
- Stevenson, C.S.; Capper, E.A.; Roshak, A.M.Y.K.; Marquez, B.; Eichman, C.; Jackson, J.R.; Mattern, M.; Gerwick, W.H.; Jacobs, R.S.; Marshall, L.A. The identification and characterization of the marine natural product scytonemin as a novel antiproliferative pharmacophore. J. Pharmacol. Exp. Ther. 2002, 303, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Malloy, K.L.; Villa, F.A.; Engene, N.; Matainaho, T.; Gerwick, L.; Gerwick, W.H. Malyngamide 2, an oxidized lipopeptide with nitric oxide inhibiting activity from a Papua New Guinea marine cyanobacterium. J. Nat. Prod. 2011, 74, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Padrines, M.; Wolf, M.; Walz, A.; Baggiolini, M. Interleukin-8 processing by neutrophil elastase, cathepsin G and proteinase-3. FEBS Lett. 1994, 352, 231–235. [Google Scholar] [CrossRef] [Green Version]
- Wiedow, O.; Meyer-Hoffert, U. Neutrophil serine proteases: Potential key regulators of cell signalling during inflammation. J. Intern. Med. 2005, 257, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Pham, C.T.N. Neutrophil serine proteases: Specific regulators of inflammation. Nat. Rev. Immunol. 2006, 6, 541–550. [Google Scholar] [CrossRef]
- Dubois, R.N.; Abramson, S.B.; Crofford, L.; Gupta, R.A.; Simon, L.S.; Van De Putte, L.B.; Lipsky, P.E. Cyclooxygenase in biology and disease. FASEB J. 1998, 12, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Patrignani, P.; Patrono, C. Cyclooxygenase inhibitors: From pharmacology to clinical read-outs. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Vane, J.R.; Botting, R.M. Anti-inflammatory drugs and their mechanism of action. Inflamm. Res. 1998, 47, 78–87. [Google Scholar] [CrossRef]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R.T. Oxidative stress and neurodegenerative diseases: A review of upstream and downstream antioxidant therapeutic options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The Role of Oxidative Stress in Neurodegenerative Diseases. Exp. Neurobiol. 2015, 24, 325. [Google Scholar] [CrossRef]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef]
- Baradaran, A.; Nasri, H.; Rafieian-Kopaei, M. Oxidative stress and hypertension: Possibility of hypertension therapy with antioxidants. J. Res. Med. Sci. 2014, 19, 358–367. [Google Scholar] [PubMed]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, Z.J.; Lao, L.; Wong, C.W.; Feng, Y. The role of oxidative stress and antioxidants in liver diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vascul. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Wada, N.; Sakamoto, T.; Matsugo, S. Multiple roles of photosynthetic and sunscreen pigments in cyanobacteria focusing on the oxidative stress. Metabolites 2013, 3, 463–483. [Google Scholar] [CrossRef]
- Hirschberg, J.; Chamovitz, D. Carotenoids in Cyanobacteria BT—The Molecular Biology of Cyanobacteria. In The Molecular Biology of Cyanobacteria; Springer: Dordrecht, The Netherlands, 1994; pp. 559–579. ISBN 978-94-011-0227-8. [Google Scholar]
- Balskus, E.P.; Walsh, C.T. The genetic and molecular basis for sunscreen biosynthesis in cyanobacteria. Science 2010, 329, 1653–1656. [Google Scholar] [CrossRef] [PubMed]
- Waditee-Sirisattha, R.; Kageyama, H.; Sopun, W.; Tanaka, Y.; Takabe, T. Identification and upregulation of biosynthetic genes required for accumulation of Mycosporine-2-glycine under salt stress conditions in the halotolerant cyanobacterium Aphanothece halophytica. Appl. Environ. Microbiol. 2014, 80, 1763–1769. [Google Scholar] [CrossRef] [PubMed]
- Kosourov, S.; Murukesan, G.; Jokela, J.; Allahverdiyeva, Y. Carotenoid biosynthesis in calothrix sp. 336/3: Composition of carotenoids on full medium, during diazotrophic growth and after long-term H2photoproduction. Plant Cell Physiol. 2016, 57, 2269–2282. [Google Scholar] [CrossRef] [PubMed]
- Al-Awadhi, F.H.; Law, B.K.; Paul, V.J.; Luesch, H. Grassystatins D-F, Potent Aspartic Protease Inhibitors from Marine Cyanobacteria as Potential Antimetastatic Agents Targeting Invasive Breast Cancer. J. Nat. Prod. 2017, 80, 2969–2986. [Google Scholar] [CrossRef]
- Al-Awadhi, F.H.; Ratnayake, R.; Paul, V.J.; Luesch, H. Tasiamide F, a potent inhibitor of cathepsins D and E from a marine cyanobacterium. Bioorg. Med. Chem. 2016, 24, 3276–3282. [Google Scholar] [CrossRef] [Green Version]
- Kwan, J.C.; Eksioglu, E.A.; Liu, C.; Paul, V.J.; Luesch, H. Grassystatins A-C from marine cyanobacteria, potent cathepsin E inhibitors that reduce antigen presentation. J. Med. Chem. 2009, 52, 5732–5747. [Google Scholar] [CrossRef] [PubMed]
- Molinski, T.F.; Reynolds, K.A.; Morinaka, B.I. Symplocin A, a linear peptide from the bahamian cyanobacterium Symploca sp. configurational analysis of N,N-dimethylamino acids by chiral-phase HPLC of naphthacyl esters. J. Nat. Prod. 2012, 75, 425–431. [Google Scholar] [CrossRef]
- Williams, P.G.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Tasiamide, a cytotoxic peptide from the marine cyanobacterium Symploca sp. J. Nat. Prod. 2002, 65, 1336–1339. [Google Scholar] [CrossRef]
- Williams, P.G.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. The isolation and structure elucidation of tasiamide B, a 4-amino-3-hydroxy-5-phenylpentanoic acid containing peptide from the marine cyanobacterium Symploca sp. J. Nat. Prod. 2003, 66, 1006–1009. [Google Scholar] [CrossRef] [PubMed]
- Mevers, E.; Haeckl, F.P.J.; Boudreau, P.D.; Byrum, T.; Dorrestein, P.C.; Valeriote, F.A.; Gerwick, W.H. Lipopeptides from the tropical marine cyanobacterium Symploca sp. J. Nat. Prod. 2014, 77, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Okamoto, K.; Tsukuba, T. Cathepsin E: An Aspartic Protease with Diverse Functions and Biomedical Implications. In Encyclopedia of Cell Biology; Academic Press: Cambridge, MA, USA, 2015; Volume 1, pp. 681–690. ISBN 9780123944474. [Google Scholar]
- Vassar, R.; Bennett, B.D.; Babu-Khan, S.; Kahn, S.; Mendiaz, E.A.; Denis, P.; Teplow, D.B.; Ross, S.; Amarante, P.; Loeloff, R.; et al. β-Secretase cleavage of Alzheimer’s amyloid precursor protein by the transmembrane aspartic protease BACE. Science 1999, 286, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Sitachitta, N.; Gerwick, W.H. Grenadadiene and grenadamide, cyclopropyl-containing fatty acid metabolites from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 1998, 61, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; McPhail, K.L.; Ligresti, A.; Di Marzo, V.; Gerwick, W.H. Semiplenamides A-G, Fatty Acid Amides from a Papua New Guinea Collection of the Marine Cyanobacterium Lyngbya semiplena. J. Nat. Prod. 2003, 66, 1364–1368. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, M.; Pereira, A.R.; Debonsi, H.M.; Ligresti, A.; Di Marzo, V.; Gerwick, W.H. Cannabinomimetic lipid from a marine cyanobacterium. J. Nat. Prod. 2011, 74, 2313–2317. [Google Scholar] [CrossRef] [PubMed]
- Mevers, E.; Matainaho, T.; Allara’, M.; Di Marzo, V.; Gerwick, W.H. Mooreamide A: A cannabinomimetic lipid from the marine cyanobacterium Moorea bouillonii. Lipids 2014, 49, 1127–1132. [Google Scholar] [CrossRef]
- Kleigrewe, K.; Almaliti, J.; Tian, I.Y.; Kinnel, R.B.; Korobeynikov, A.; Monroe, E.A.; Duggan, B.M.; Di Marzo, V.; Sherman, D.H.; Dorrestein, P.C.; et al. Combining Mass Spectrometric Metabolic Profiling with Genomic Analysis: A Powerful Approach for Discovering Natural Products from Cyanobacteria. J. Nat. Prod. 2015, 78, 1671–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aizpurua-Olaizola, O.; Elezgarai, I.; Rico-Barrio, I.; Zarandona, I.; Etxebarria, N.; Usobiaga, A. Targeting the endocannabinoid system: Future therapeutic strategies. Drug Discov. Today 2017, 22, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Hill, M.N.; Cheer, J.F.; Wotjak, C.T.; Holmes, A. The endocannabinoid system as a target for novel anxiolytic drugs. Neurosci. Biobehav. Rev. 2017, 76, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Evolution of the cumulative number of metabolite families according to the number of analyzed publications used for the construction of the database. The arrow indicates a reclassification event of all the structural variants of one molecule in a unique entry of “family” [13,25,26]. We observed a progressive stabilization of the number of compound families in the database that supports the postulation of the exhaustiveness of the present database.
Figure 1.
Evolution of the cumulative number of metabolite families according to the number of analyzed publications used for the construction of the database. The arrow indicates a reclassification event of all the structural variants of one molecule in a unique entry of “family” [13,25,26]. We observed a progressive stabilization of the number of compound families in the database that supports the postulation of the exhaustiveness of the present database.

Figure 2.
Proportion of families of compound by taxonomical level. (A) The pie chart represents the percentage of compound families for each taxonomical family. Note that some compound families can be produced by several cyanobacterial families. The “Other” category concerns other taxonomical families that produce less than two compound families. (B) The histogram shows the number of compound families for each genus. The “Other” category corresponds to genera producing less than four compound families. * indicates cyanobacterial assemblages whom the real metabolite producer is still undetermined. The boxes indicate the environmental origins for the corresponding genera. For both charts, the colors correspond to the taxonomical order of each genus or family.
Figure 2.
Proportion of families of compound by taxonomical level. (A) The pie chart represents the percentage of compound families for each taxonomical family. Note that some compound families can be produced by several cyanobacterial families. The “Other” category concerns other taxonomical families that produce less than two compound families. (B) The histogram shows the number of compound families for each genus. The “Other” category corresponds to genera producing less than four compound families. * indicates cyanobacterial assemblages whom the real metabolite producer is still undetermined. The boxes indicate the environmental origins for the corresponding genera. For both charts, the colors correspond to the taxonomical order of each genus or family.

Figure 3.
Classification of the 260 cyanobacterial metabolite families according to their respective chemical classes. All the molecules have been classified into these different classes according to their respective structural characteristics. For example, the depsipeptides are a class of peptides containing an ester bond and macrolides are molecules exhibiting a macrocycle and one or more lactone functions. Some examples of cyanobacterial molecules belonging to these classes are illustrated. Hapalindole A (alkaloids), Oscillapeptin A (depsipeptides), Minutissamide A (lipopeptides), Caylobolide B (macrolides/lactones), Anabaenopeptin E (peptides), β-carotene (terpenes), Cyclodextrin phosphate (polysaccharides), Lyngbic acid (lipids), and Cylindrocyclophane A (polyketides). The main characteristics of each chemical class are highlighted in red. All the structures were obtained from the ChEMBL Database (https://www.ebi.ac.uk/chembl/).
Figure 3.
Classification of the 260 cyanobacterial metabolite families according to their respective chemical classes. All the molecules have been classified into these different classes according to their respective structural characteristics. For example, the depsipeptides are a class of peptides containing an ester bond and macrolides are molecules exhibiting a macrocycle and one or more lactone functions. Some examples of cyanobacterial molecules belonging to these classes are illustrated. Hapalindole A (alkaloids), Oscillapeptin A (depsipeptides), Minutissamide A (lipopeptides), Caylobolide B (macrolides/lactones), Anabaenopeptin E (peptides), β-carotene (terpenes), Cyclodextrin phosphate (polysaccharides), Lyngbic acid (lipids), and Cylindrocyclophane A (polyketides). The main characteristics of each chemical class are highlighted in red. All the structures were obtained from the ChEMBL Database (https://www.ebi.ac.uk/chembl/).

Figure 4.
Number of metabolite families observed for each type of activity. The percentage represents the proportion of one activity compared to the whole occurrence of activities detected (n = 362). Some compounds present various activities and are considered several times.
Figure 4.
Number of metabolite families observed for each type of activity. The percentage represents the proportion of one activity compared to the whole occurrence of activities detected (n = 362). Some compounds present various activities and are considered several times.

Figure 5.
Classification of the 260 metabolite families according to their respective activities and chemical classes. The number of metabolite families is symbolized by the disc diameters, for each activity and each chemical class. For example, the first circle represents the number of alkaloids that exhibit a hepatotoxic activity (in this case, one family of metabolites). Colors correspond to the different categories of activity targets. For example, cytotoxicity and hepatotoxicity are tested in vitro against cell lines while neurotoxicity, antioxidant, and anti-inflammatory activities can be biochemically tested for specific cellular mechanisms (such as the sodium influx, the scavenging of ROS (reactive oxygen species), and the inhibition of cytokines).
Figure 5.
Classification of the 260 metabolite families according to their respective activities and chemical classes. The number of metabolite families is symbolized by the disc diameters, for each activity and each chemical class. For example, the first circle represents the number of alkaloids that exhibit a hepatotoxic activity (in this case, one family of metabolites). Colors correspond to the different categories of activity targets. For example, cytotoxicity and hepatotoxicity are tested in vitro against cell lines while neurotoxicity, antioxidant, and anti-inflammatory activities can be biochemically tested for specific cellular mechanisms (such as the sodium influx, the scavenging of ROS (reactive oxygen species), and the inhibition of cytokines).

Table 6.
Cytotoxic metabolites extracted from the database.
| Molecule Family | Chemical Classes | Activity | Producing Organisms | References |
|---|---|---|---|---|
| Tubercidin | Nucleoside | -Cytotoxic -Microtubule stabilizer | Tolypothrix byssoidea H-6-2, Scytonema saleyeriense var. indica CV-14-1, Plectonema radiosum DF-6-1, Tolypothrix distorta BL-11-2 | [198,199,200] |
| Aurilides | Depsipeptide | -Cytotoxic -Lethal activity -Anti-swarming -Antiprotozoal -Induce loss of microfilament network | Lyngbya majuscula, Okeania sp., Lyngbya sp. | [70,202,203,204,205] |
| Swinholide-type | Macrolide | -Cytotoxic -Actin microfilament disruption | Symploca sp., Geitlerinema sp., Nostoc sp. UHCC0451, Phormidium sp. | [206,207,208] |
| Anabaenolysins | Lipopeptide | -Cytotoxic -Antifungal -Hemolytic activity -Ability to permeabilize cell membranes | Anabaena sp. XPORK 15F, Anabaena sp. XSPORK 27C | [209,210] |
Table 7.
Serine protease inhibitor metabolites extracted from the database.
| Molecule Family | Chemical Classes | Activity | Producing Organisms | References |
|---|---|---|---|---|
| Spumigins | Peptide | -Proteases inhibitory activity | Nodularia spumigena AV1 & CCY 9414, Anabaena compacta NIES-835 | [239,240,241] |
| Cyanopeptolin-like | Depsipeptide | -Protease inhibitory activity -Other enzyme inhibition -Cytotoxic -Lethal -Antibacterial -Antifungal -Antiprotozoal | Microcystis sp., Microcystis aeruginosa, Aphanocapsa sp.; Microchaete loktahensis, Planktothrix agardhii, Scytonema hofmanni, Lyngbya sp., Lyngbya confervoides, Lyngbya spp., Lyngbya semiplena, Microcystis viridis, Dichothrix utahensis, Nostoc sp., Nostoc minutum, Planktothrix rubescens, Lyngbya majuscula-Schizothrix sp. (Assemblage), Stigonema sp., Symploca sp., Symploca hydnoides, Nostoc insulare | [30,31,159,185,186,188,243,244,245,246,247,248,249,250,251,252,253,254,255,256,257,258,259,260,261,262,263,264,265,266,267,268,269,270,271,272,273,274,275,276,277,278,279,280,281,282,283,284,285,286,287,288,289,290,291,292] |
| Carmaphycins | Peptide | -Protease inhibition -Cytotoxic -Antiprotozoal | Symploca sp. WHG NAC15/Dec/08–5 | [293,294] |
Table 8.
HDACs inhibitor metabolites extracted from the database.
| Molecule Family | Chemical Classes | Activity | Producing Organisms | References |
|---|---|---|---|---|
| Largazole | Depsipeptide | -Histone deacetylases inhibitor -Cytotoxic -Other enzyme inhibition -Pro-drug | Symploca sp. | [311,312,313,314,315,316] |
| Santacruzamate A | Carboxylic acid derived | -Histone deacetylases inhibitor -Cytotoxic | Symploca sp. PAC-19-FEB-10-1 | [317] |
Table 9.
Anti-inflammatory metabolites extracted from the database.
| Molecule Family | Chemical Classes | Activity | Producing Organisms | References |
|---|---|---|---|---|
| Aeruginosins | Peptide | -Anti-inflammatory activity -Protease inhibitor -No cytotoxicity | Microcystis aeruginosa NIES-98, NIES-298, NIES-101, NIES-89. Microcystis viridis NIES-102 Planktothrix agardhii CYA 126/8. Nodularia spumigena CCY9414. Nostoc sp. Lukesova 30/93 | [62,321,322,323] |
| Phycocyanin | Peptide | -Anti-inflammatory -Antioxidant -Specific inhibitor of COX-2 -No lethality | All | [59,64,324,325,326] |
| Scytonemin | Alkaloid | -Anti-inflammatory -Enzyme inhibition -No cytotoxicity | Stigonema sp., Nostoc punctiforme, Anabaena variabilis, Anabaena ambigua, Aphanocapsa/Synechocystis sp. (assembly), Aulosira fertilissima, Calothrix sp., Calothrix parietina, Calothrix crustacea, Chlorogloeopsis sp., Chroococcidiopsis sp., Chroococcus sp.; Cylindrospermum sp., Diplocolon sp., Entophysalis granulos, Gloeocapsa sp., Hapalosiphon sp., Hapalosiphon fontinalis; Lyngbya sp., Lyngbya aestuarii, Nostoc parmelioides, Nostoc commune, Nostoc microscopium, Nostoc pruniforme, Phormidium sp., Pleurocapsa sp., Rivularia atra, Rivularia sp., Schizothrix sp., Scytonema sp., Tolyothrix sp., Tolypothrix tenni, Westiellopsis prolifica, Scytonema hoffmani | [65,327,328,329,330,331] |
Table 10.
Antioxidant metabolites extracted from the database.
| Molecule Family | Chemical Classes | Activity | Producing Organisms | References |
|---|---|---|---|---|
| Carotenoids | Terpenoid | -Antioxidant -Sunscreen | All | [56,345,346] |
| Chlorophylls | Chlorin | -Photosynthesis -Antioxidant -Pro-oxidant (sensitizer for singlet oxygen production) | All | [57,345] |
| MAAs | Cyclohexenone linked with an amino acid | -Antioxidant -Sunscreens | Synechocystis sp. PCC 6803, Gloeocapsa sp. CU-2556, Aphanothece halophytica, Gloeocapsa sp., Euhalothece sp., Microcystis aeruginosa, Arthrospira sp. CU2556, Lyngbya sp. CU2555, Leptolyngbya sp., Phormidium sp., Lyngbya cf. aestuarii, Microcoleus chthonoplastes, Microcoleus sp., Oscillatoria spongelidae, Trichodesmium spp., Anabaena sp., Anabaena doliolum, Anabaena variabilis PCC 7937, Nostoc sp., Nostoc commune var. Vaucher, Nostoc commune, Scytonema sp., Nostoc punctiforme ATCC 29133, Nostoc sp. HKAR-2 and HKAR-6, Nodularia baltica, Nodularia harveyana, Nodularia spumigena, Aphanizomenon flos-aquae, Chlorogloeopsis PCC 6912 | [58,345,347,348] |
| Phycocyanin | Peptide | -Anti-inflammatory -Antioxidant -Specific inhibitor of COX-2 -No lethality | All | [59,64,324,325,326] |
Table 11.
Other metabolites extracted from our database with promising biomedical potential.
| Molecule Family | Chemical Classes | Activity | Producing Organisms | References |
|---|---|---|---|---|
| Grassystatins-Tasiamides | Depsipeptide | -Protease inhibitory activity -Cytotoxic -Reduce antigen presentation in dendritic cells | Lyngbya confervoides, Symploca sp., Symploca sp. NHI304, Lyngbya sp. NIH399 | [350,351,352,353,354,355,356] |
| Columbamides | Acyl amide | -CB1 and CB2 ligands | Moorea bouillonii PNG05-198 | [363] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Demay, J.; Bernard, C.; Reinhardt, A.; Marie, B. Natural Products from Cyanobacteria: Focus on Beneficial Activities. Mar. Drugs 2019, 17, 320. https://0-doi-org.brum.beds.ac.uk/10.3390/md17060320
AMA Style
Demay J, Bernard C, Reinhardt A, Marie B. Natural Products from Cyanobacteria: Focus on Beneficial Activities. Marine Drugs. 2019; 17(6):320. https://0-doi-org.brum.beds.ac.uk/10.3390/md17060320
Chicago/Turabian StyleDemay, Justine, Cécile Bernard, Anita Reinhardt, and Benjamin Marie. 2019. "Natural Products from Cyanobacteria: Focus on Beneficial Activities" Marine Drugs 17, no. 6: 320. https://0-doi-org.brum.beds.ac.uk/10.3390/md17060320
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.