ATP- and Polyphosphate-Dependent Glucokinases from Aerobic Methanotrophs

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria and Growth Conditions
2.2. Identification of Genes Coding for PolyP- and ATP-Dependent Glucokinases
2.3. Cloning of Glucokinases Encoding Genes
2.4. Purification of the Recombinant Proteins
2.5. Determination of Molecular Masses of Glucokinases
2.6. Essay of Enzyme Activities
2.7. Biochemical Characterization of Glucokinases
2.8. Analysis of Nucleotide and Amino Acid Sequences
3. Results
3.1. Identification of Glucokinase Encoding Genes in Methylomonas sp. 12 and Purification of Glucokinases
3.2. Biochemical Properties of ATP-Glk
3.3. Biochemical Properties of PolyP-Glk
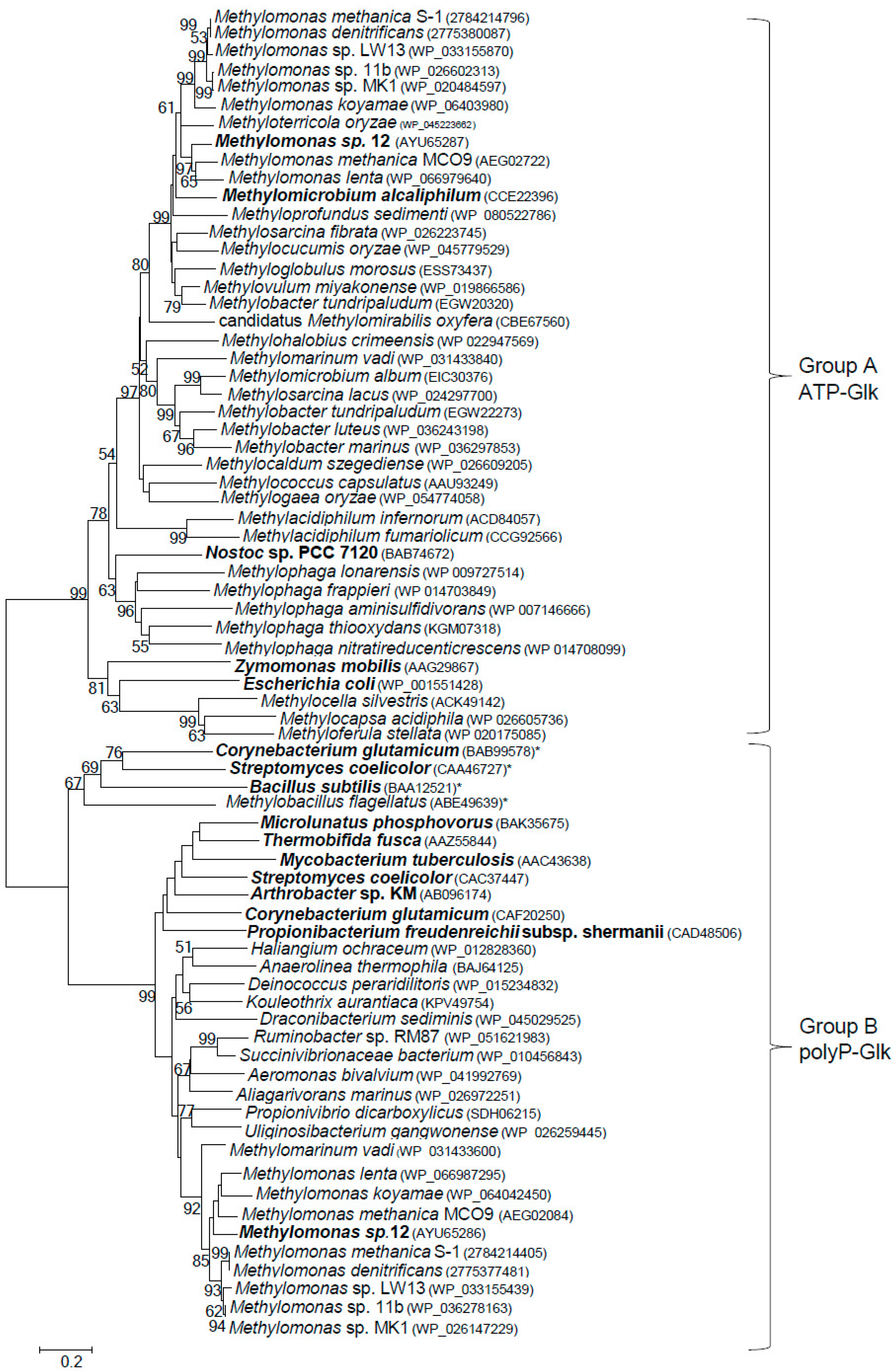
3.4. Distribution of Glucokinases in Methanotrophs
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Curtis, S.J.; Epstein, W. Phosphorylation of D-glucose in Escherichia coli mutants defective in glucosephosphotransferase, mannosephosphotransferase, and glucokinase. J. Bacteriol. 1975, 122, 1189–1199. [Google Scholar] [PubMed]
- Postma, P.W.; Lengeler, J.W.; Jacobson, G.R. Phosphoenolpyruvate:carbohydrate phosphotransferase systems of bacteria. Microbiol. Rev. 1993, 57, 543–594. [Google Scholar]
- Hanson, R.S.; Hanson, T.E. Methanotrophic Bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar] [PubMed]
- McDonald, I.R.; Bodrossy, L.; Chen, Y.; Murrell, J.C. Molecular ecology techniques for the study of aerobic methanotrophs. Appl. Environ. Microbiol. 2008, 74, 1305–1315. [Google Scholar] [CrossRef] [PubMed]
- Op den Camp, H.J.M.; Islam, T.; Stott, M.B.; Harhangi, H.R.; Hynes, A.; Schouten, S.; Jetten, M.S.M.; Birkeland, N.-K.; Pol, A.; Dunfield, P.F. Environmental, genomic and taxonomic perspectives on methanotrophic Verrucomicrobia. Environ. Microbiol. 2009, 1, 293–306. [Google Scholar] [CrossRef]
- Ettwig, K.F.; Butler, M.K.; Le Paslier, D.; Pelletier, E.; Mangenot, S.; Kuypers, M.M.; Schreiber, F.; Dutilh, B.E.; Zedelius, J.; de Beer, D.; et al. Nitrite-driven anaerobic methane oxidation by oxygenic bacteria. Nature 2010, 464, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Mustakhimov, I.I.; Rozova, O.N.; Solntseva, N.P.; Khmelenina, V.N.; Reshetnikov, A.S.; Trotsenko, Y.A. The properties and potential metabolic role of glucokinase in halotolerant obligate methanotroph Methylomicrobium alcaliphilum 20Z. Ant. Leeuwenhoek. 2017, 110, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Galchenko, V.F.; Shishkina, V.N.; Tiurin, V.S.; Trotsenko, Y.A. Isolation of pure methanotrophic cultures and their properties. Mikrobiologiia 1975, 44, 844–850. [Google Scholar]
- Gayazov, R.R.; Shishkina, V.N.; Mshensky, Y.N.; Trotsenko, Y.A. Effect of temperature on growth and metabolism of Methylomonas methanica. Doklady of Academy of Science 1985, 284, 746–748. (in Russian). [Google Scholar]
- Trotsenko, Y.A.; Shishkina, V.N. Studies on phosphate metabolism in obligate methanotrophs. FEMS Microbiol. Rev. 1990, 87, 267–271. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.J.; Costello, A.; Lidstrom, M.E.; Murrell, J.C. Evidence that particulate methane monooxygenase and ammonia monooxygenase may be evolutionarily related. FEMS Microbiol. Lett. 1995, 132, 203–208. [Google Scholar] [CrossRef]
- Costello, A.M.; Lidstrom, M.E. Molecular characterization of functional and phylogenetic genes from natural populations of methanotrophs in lake sediments. Appl. Environ. Microbiol. 1999, 65, 5066–5074. [Google Scholar] [PubMed]
- Sambrook, J.F.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Slater, G.G. Stable pattern formation and determination of molecular size by pore-limit electrophoresis. Anal. Chem. 1969, 41, 1039–1041. [Google Scholar] [CrossRef] [PubMed]
- Leitao, J.M.; Lorenz, B.; Bachinski, N.; Wilhelm, C.; Muller, W.; Schroder, H.C. Osmotic-stress-induced synthesis and degradation of inorganic polyphosphates in the alga Phaeodactylum tricornutum. Mar. Ecol. Prog. Ser. 1995, 121, 279–288. [Google Scholar] [CrossRef]
- Thomson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucl. Acids Res. 1997, 24, 4876–4882. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Boil. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Philips, N.F.B.; Horn, P.J.; Wood, H.G. The polyphosphate and ATP-dependent glucokinase from Propionibacterium shermanii: Both activities are catalyzed be the same protein. Arch. Biochem. Biophys. 1993, 300, 309–319. [Google Scholar] [CrossRef]
- Lindner, S.N.; Knebel, S.; Pallerla, S.R.; Schoberth, S.M.; Wendisch, V.F. Cg2091 encodes a polyphosphate/ATP-dependent glucokinase of Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2010, 87, 703–713. [Google Scholar] [CrossRef]
- Hansen, T.; Reichstein, B.; Schmid, R.; Schonheit, P. The first archaeal ATP-dependent glucokinase, from the hyperthermophilic crenarchaeon Aeropyrum pernix, represents a monomeric, extremely thermophilic ROK glucokinase with broad hexose specificity. J. Bacteriol. 2002, 184, 5955–5965. [Google Scholar] [CrossRef] [PubMed]
- Pimentel-Schmitt, E.; Thomae, A.; Amon, J.; Klieber, M.A.; Roth, H.M.; Muller, Y.A.; Jahreis, K.; Burkovski, A.; Titgemeyer, F. A glucose kinase from Mycobacterium smegmatis. J. Mol. Microbiol. Biotechnol. 2007, 12, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Myung, S.; Zhang, Y.H. One-step purification and immobilization of thermophilic polyphosphate glucokinase from Thermobifida fusca YX: Glucose-6-phosphate generation without ATP. Appl. Microbiol. Biotechnol. 2012, 93, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Mukai, T.; Kawai, S.; Matsukawa, H.; Matuo, Y.; Murata, K. Characterization and molecular cloning of a novel enzyme, inorganic polyphosphate/ATP-glucomannokinase, of Arthrobacter sp. strain KM. Appl. Environ. Microbiol. 2003, 69, 3849–3857. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Lee, S.O.; Hamaoka, K.; Kato, J.; Takiguchi, N.; Nakamura, K.; Ohtake, H.; Kuroda, A. Strictly polyphosphate-dependent glucokinase in a polyphosphate-accumulating bacterium, Microlunatus phosphovorus. J. Bacteriol. 2003, 185, 5654–5656. [Google Scholar] [CrossRef]
- Klemke, F.; Beyer, G.; Sawade, L.; Saitov, A.; Korte, T.; Maldener, I.; Lockau, W.; Nurnberg, D.J.; Volkmer, T. All1371 is a polyphosphate-dependent glucokinase in Anabaena sp. PCC 7120. Microbiology 2014, 160, 2807–2819. [Google Scholar] [CrossRef]
- Alia, A.; Wathesb, D.C.; Swalic, A.; Burnsa, H.; Burnsa, S. A novel mammalian glucokinase exhibiting exclusive inorganic polyphosphate dependence in the cell nucleus. Biochem. Biophys. 2017, 12, 151–157. [Google Scholar] [CrossRef]
- Hsieh, P.C.; Shenoy, B.C.; Jentoft, J.E.; Phillips, N.F. Purification of polyphosphate and ATP glucose phosphotransferase from Mycobacterium tuberculosis H37Ra: Evidence that poly(P) and ATP glucokinase activities are catalyzed by the same enzyme. Protein Expr. Purif. 1993, 4, 76–84. [Google Scholar] [CrossRef]
- Lunin, V.V.; Li, Y.; Schrag, J.D.; Iannuzzi, P.; Cygler, M.; Matte, A. Crystal structures of Escherichia coli ATP-dependent glucokinase and its complex with glucose. J. Bacteriol. 2004, 86, 6915–6927. [Google Scholar] [CrossRef]
- Kawai, S.; Mukai, T.; Mori, S.; Mikami, B.; Murata, K. Hypothesis: Structures, evolution, and ancestor of glucose kinases in the hexokinase family. J. Biosci. Bioeng. 2005, 99, 320–330. [Google Scholar] [CrossRef]
- Shishkina, V.N.; Trotsenko, Y.A. Effect of glucose on growth and metabolism of obligate methanotrophs. Microbiology 1988, 57, 917–923. [Google Scholar]
- Hirayama, H.; Fuse, H.; Abe, M.; Miyazaki, M.; Nakamura, T.; Nunoura, T.; Furushima, Y.; Yamamoto, H.; Takai, K. Methylomarinum vadi gen. nov., sp. nov., a methanotroph isolated from two distinct marine environments. Int. J. Syst. Evol. Microbiol. 2013, 63, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.D.; Hirayama, H.; Sakai, Y.; Dunfield, P.F.; Klotz, M.G.; Knief, C.; Op den Camp, H.J.M.; Jetten, M.S.M.; Khmelenina, V.N.; Trotsenko, Y.A.; et al. Draft genome sequences of gammaproteobacterial methanotrophs isolated from marine ecosystems. Genome Announc. 2016, 4, e01629-15. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effector, mM | ATP-Glk * | PolyP-Glk ** |
|---|---|---|
| Control | 100 * | 100 ** |
| Phosphoenolpyruvate, 5 | 99.8 | 111.3 |
| Pyruvate, 5 | 114.9 | 103.3 |
| Fructose-1-phosphate, 5 | 78.8 | 101.4 |
| Fructose-6-phosphate, 5 | 115.8 | 103.0 |
| Fructose-1,6- phosphate, 5 | 101.6 | 101.4 |
| Glucose-1- phosphate, 5 | 101.7 | 107.2 |
| Ribose-5-phosphate | nt | 117.8 |
| Adenosine triphosphate | 100 | 155.0 |
| Adenosine diphosphate, 5 | 48.9 | 93.0 |
| Adenosine monophosphate, 5 | 110.3 | 84.1 |
| KH2PO4, 3 | 86.1 | nt |
| Glycerate, 1 | 109. 0 | nt |
| Oxaloacetate, 1 | 108.8 | 105.3 |
| α-Ketoglutarate, 1 | 108.8 | 99.7 |
| Malate, 1 | 111.9 | 101.6 |
| Citrate, 1 | 76.6 | 106.7 |
| Isocitrate, 1 | 115.7 | nt |
| Succinate, 1 | 114.9 | nt |
| Polyphosphate, 0.075 | 117.3 | 100 |
| Inorganic pyrophosphate, 2 | 82.3 | 104.0 |
| Glyceraldehyde-3-phosphate | nt | 111.6 |
| Parameter | Substrate | Values | |
|---|---|---|---|
| ATP-Glk | PolyP-Glk | ||
| Vmax (U/mg) | Glucose | 92.5 ± 1.8 | 7.04 ± 0.39 |
| Glucosamine | 94.4 | ||
| ATP | 106.4 ± 2.4 | - | |
| PolyP(n=45) | - | 7.28 ± 0.45 | |
| Km app (mM) | Glucose | 0.08 ± 0.007 | 0.45 ± 0.12 |
| ATP | 0.078 ± 0.0056 | - | |
| PolyP (n=45) | - | 0.021 ± 0.0039 | |
| kcat substrate (1/min) | Glucose | 6.57 | 0.395 |
| ATP | 7.55 | - | |
| PolyP45 | - | 0.409 | |
| kcat/Km (1/min mM) | Glucose | 82.09 | 0.87 |
| ATP | 96.85 | - | |
| PolyP (n=45) | - | 19.476 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reshetnikov, A.S.; Solntseva, N.P.; Rozova, O.N.; Mustakhimov, I.I.; Trotsenko, Y.A.; Khmelenina, V.N. ATP- and Polyphosphate-Dependent Glucokinases from Aerobic Methanotrophs. Microorganisms 2019, 7, 52. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7020052
Reshetnikov AS, Solntseva NP, Rozova ON, Mustakhimov II, Trotsenko YA, Khmelenina VN. ATP- and Polyphosphate-Dependent Glucokinases from Aerobic Methanotrophs. Microorganisms. 2019; 7(2):52. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7020052
Chicago/Turabian StyleReshetnikov, Alexander S., Natalia P. Solntseva, Olga N. Rozova, Ildar I. Mustakhimov, Yuri A. Trotsenko, and Valentina N. Khmelenina. 2019. "ATP- and Polyphosphate-Dependent Glucokinases from Aerobic Methanotrophs" Microorganisms 7, no. 2: 52. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7020052