
Effect of the Algaecide Palmitoleic Acid on the Immune Function of the Bay Scallop Argopecten irradians

 ,
, 
Abstract
:1. Introduction
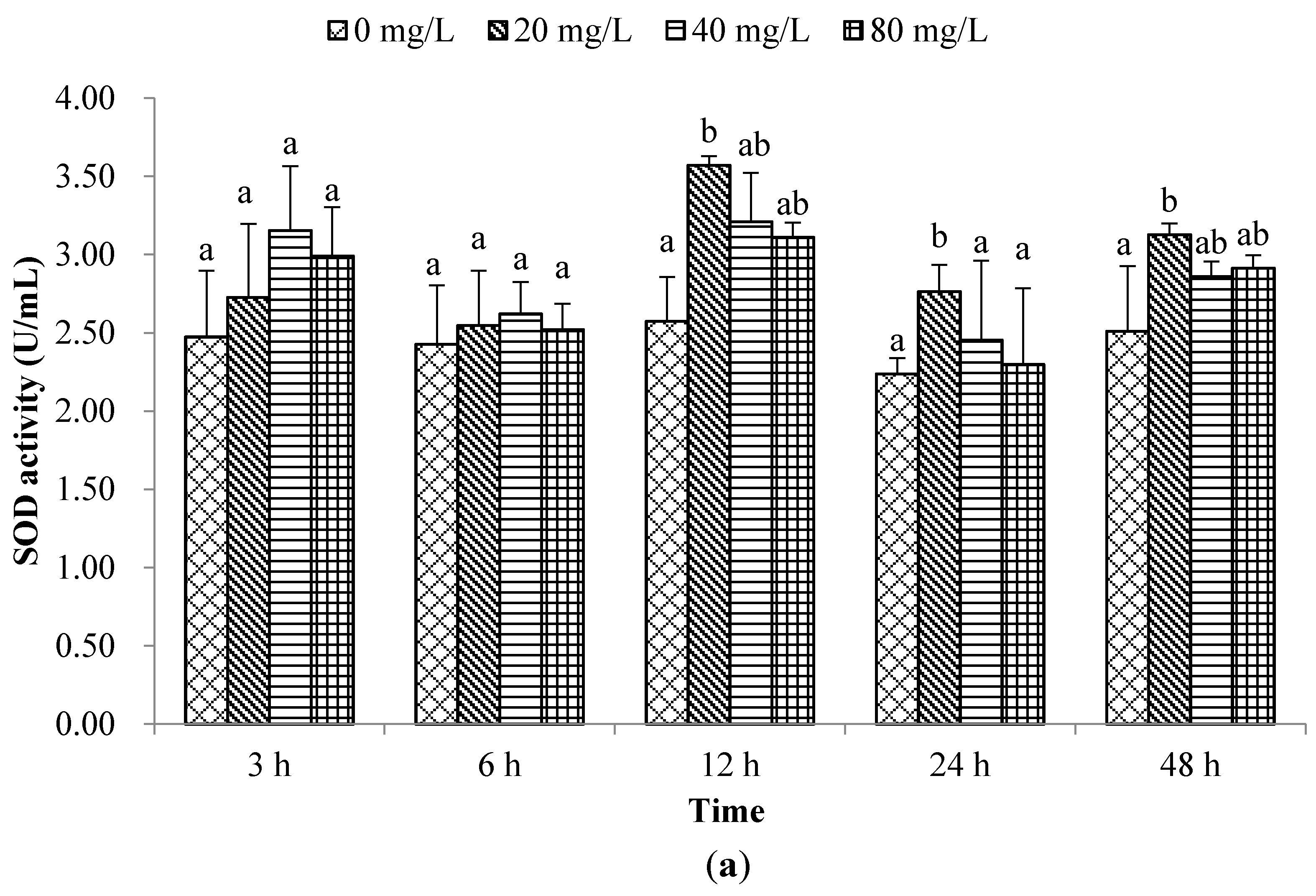
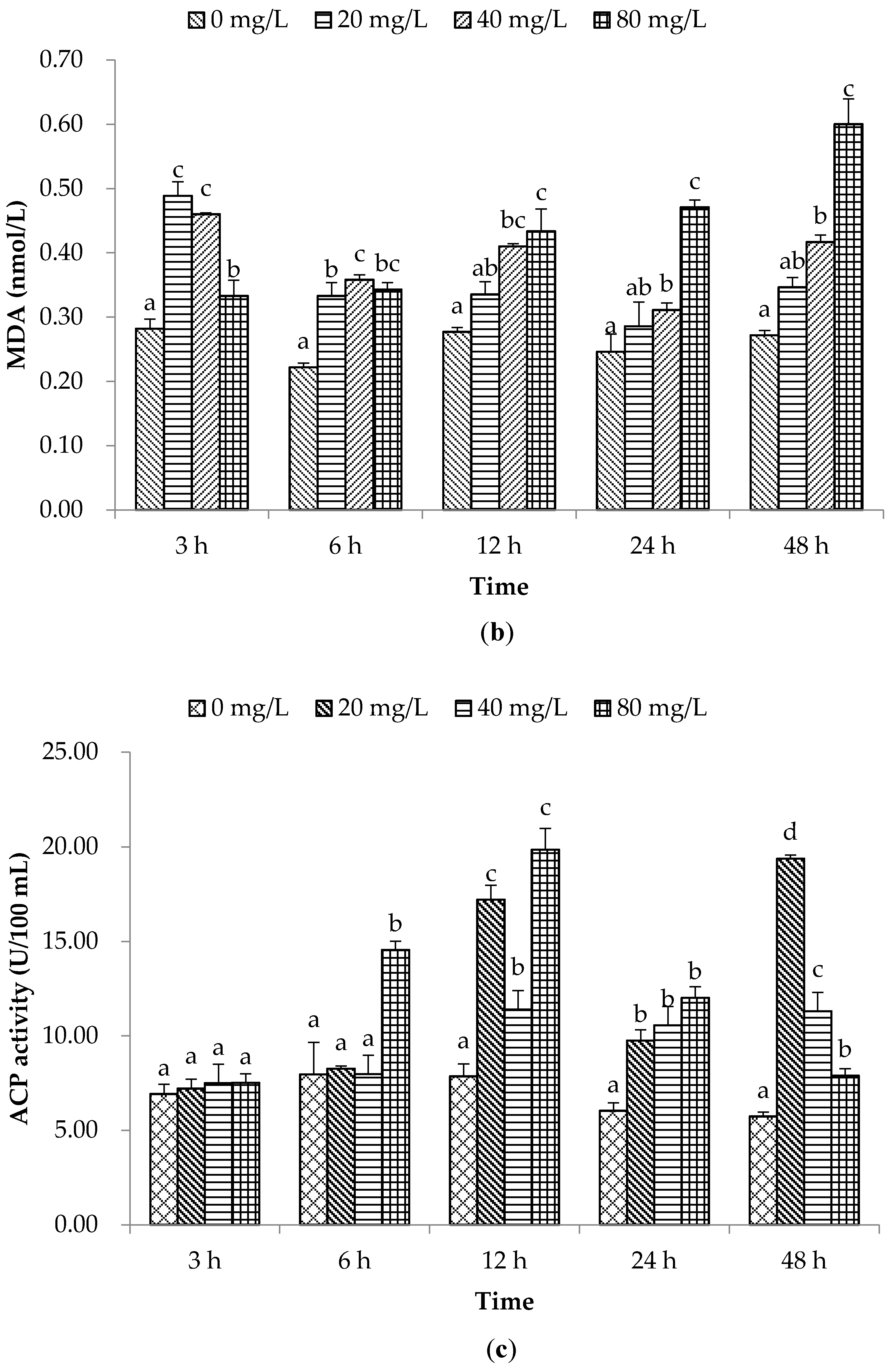
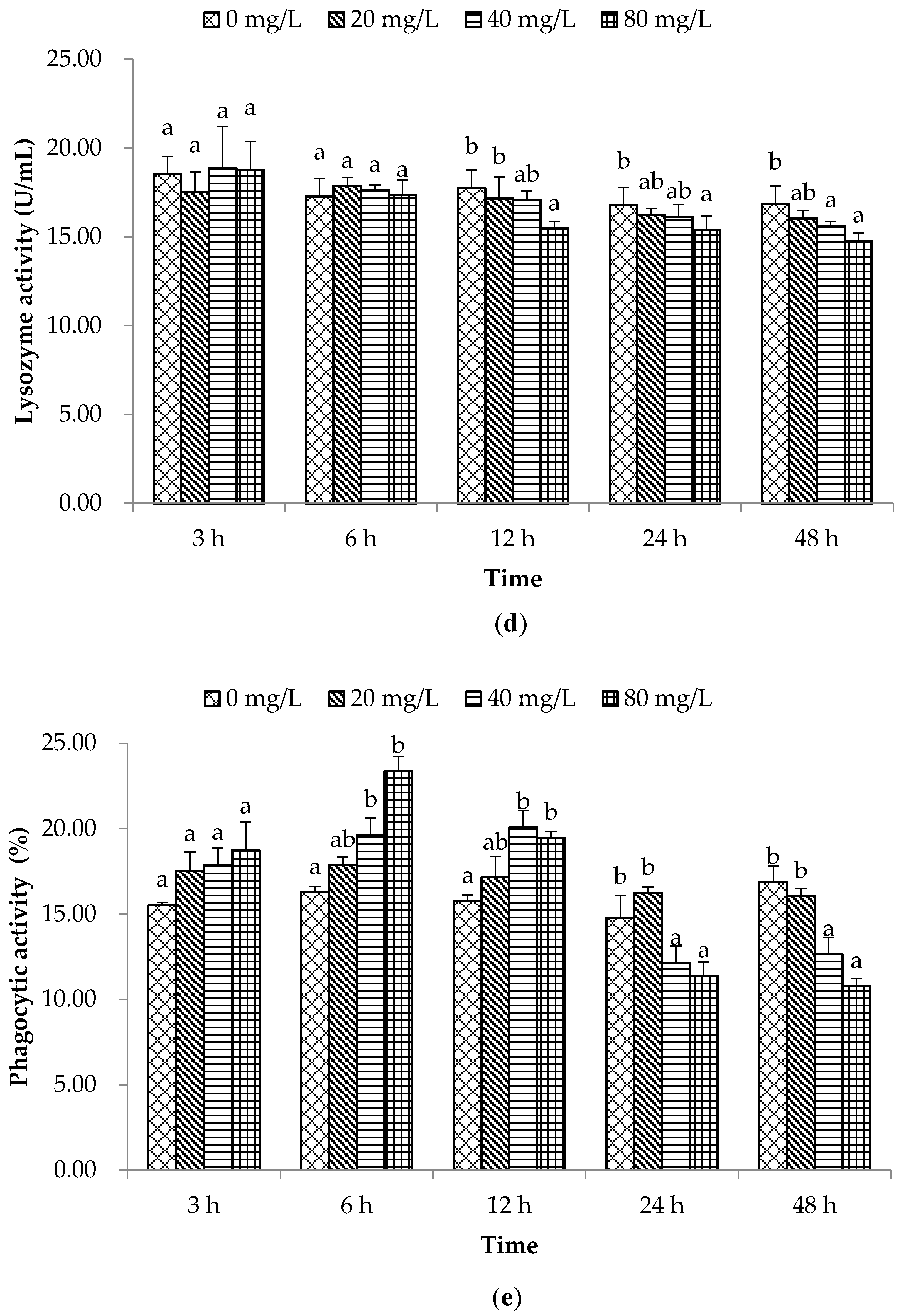
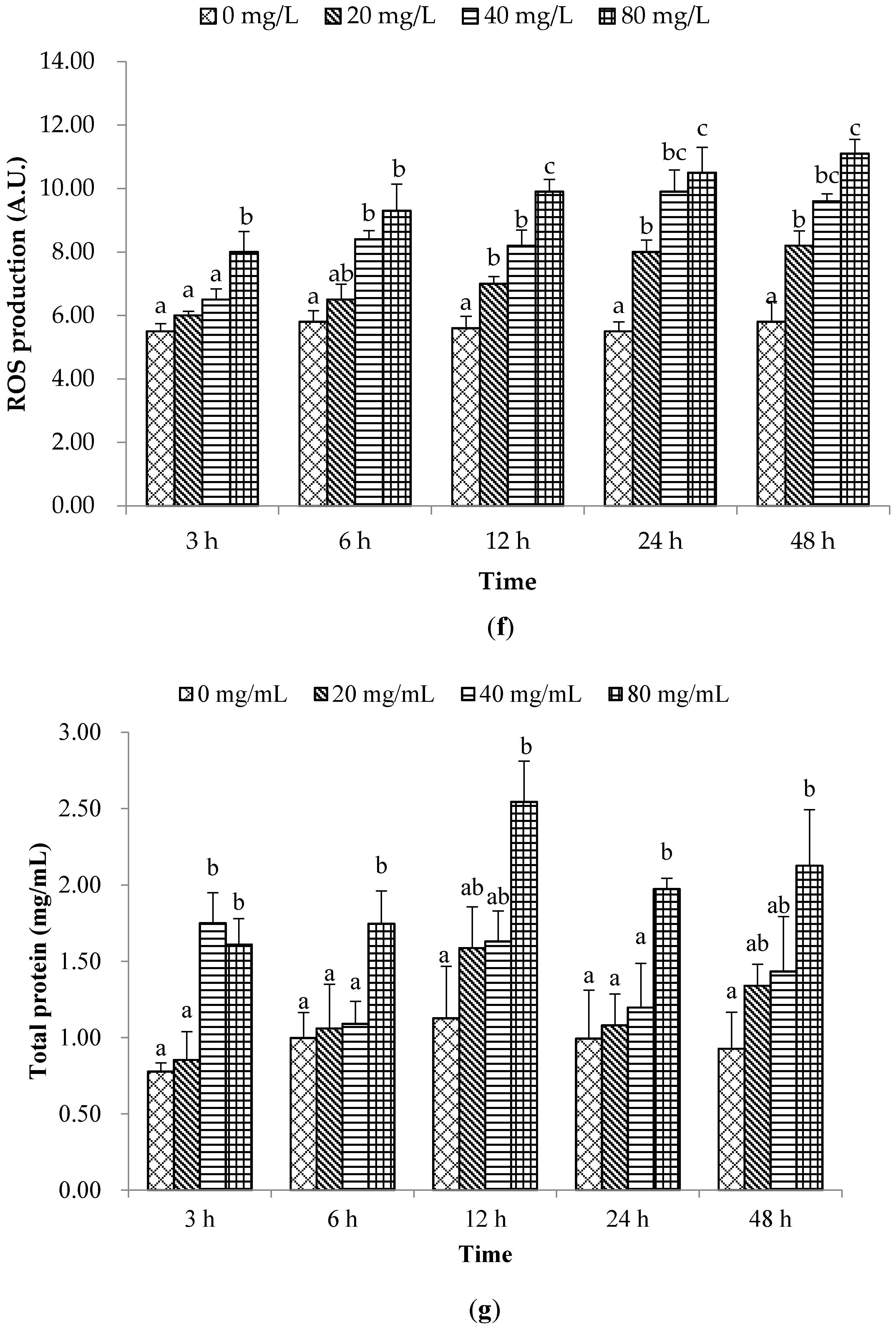
2. Results and Discussion
2.1. Non-Special Immune Responses
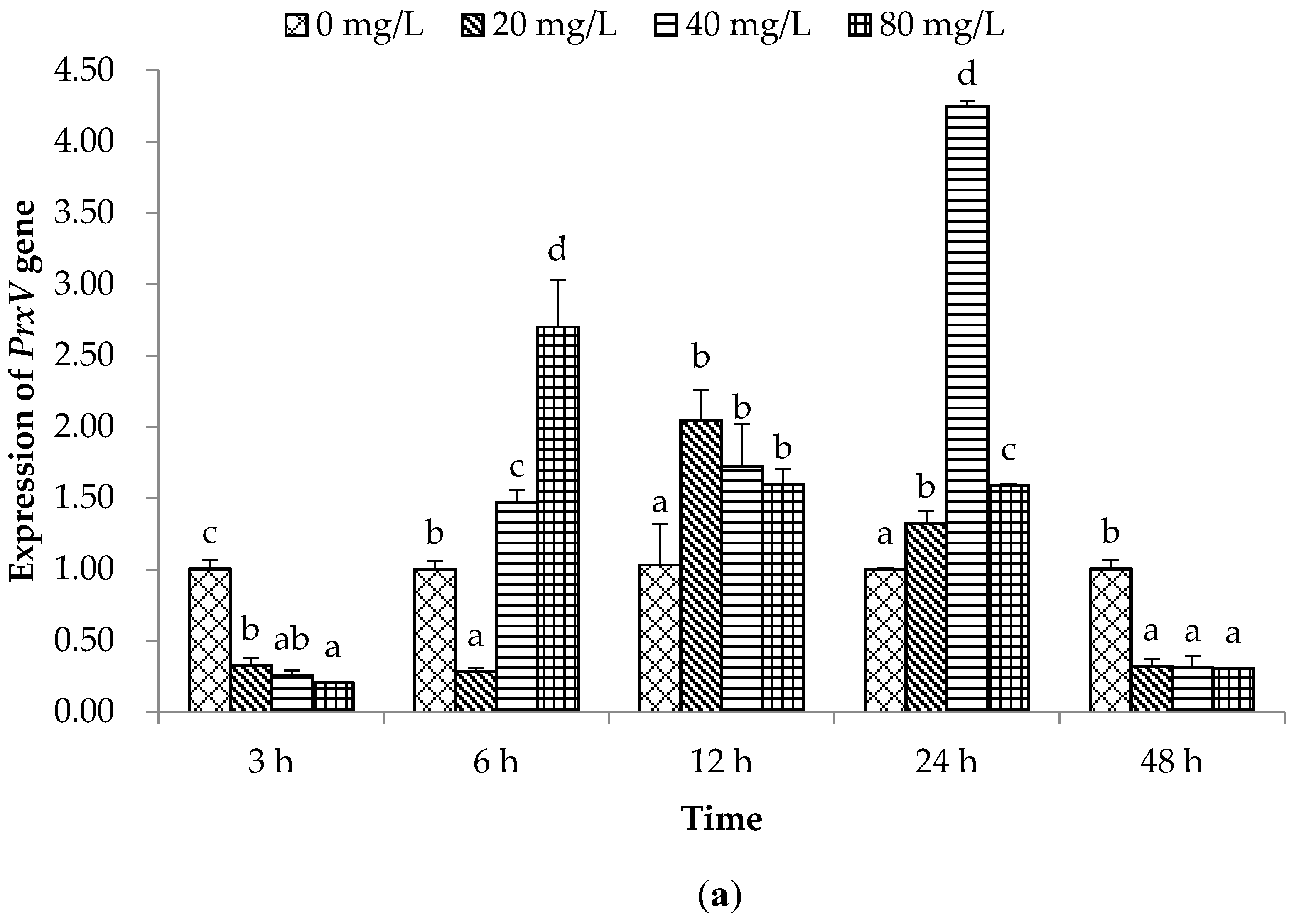
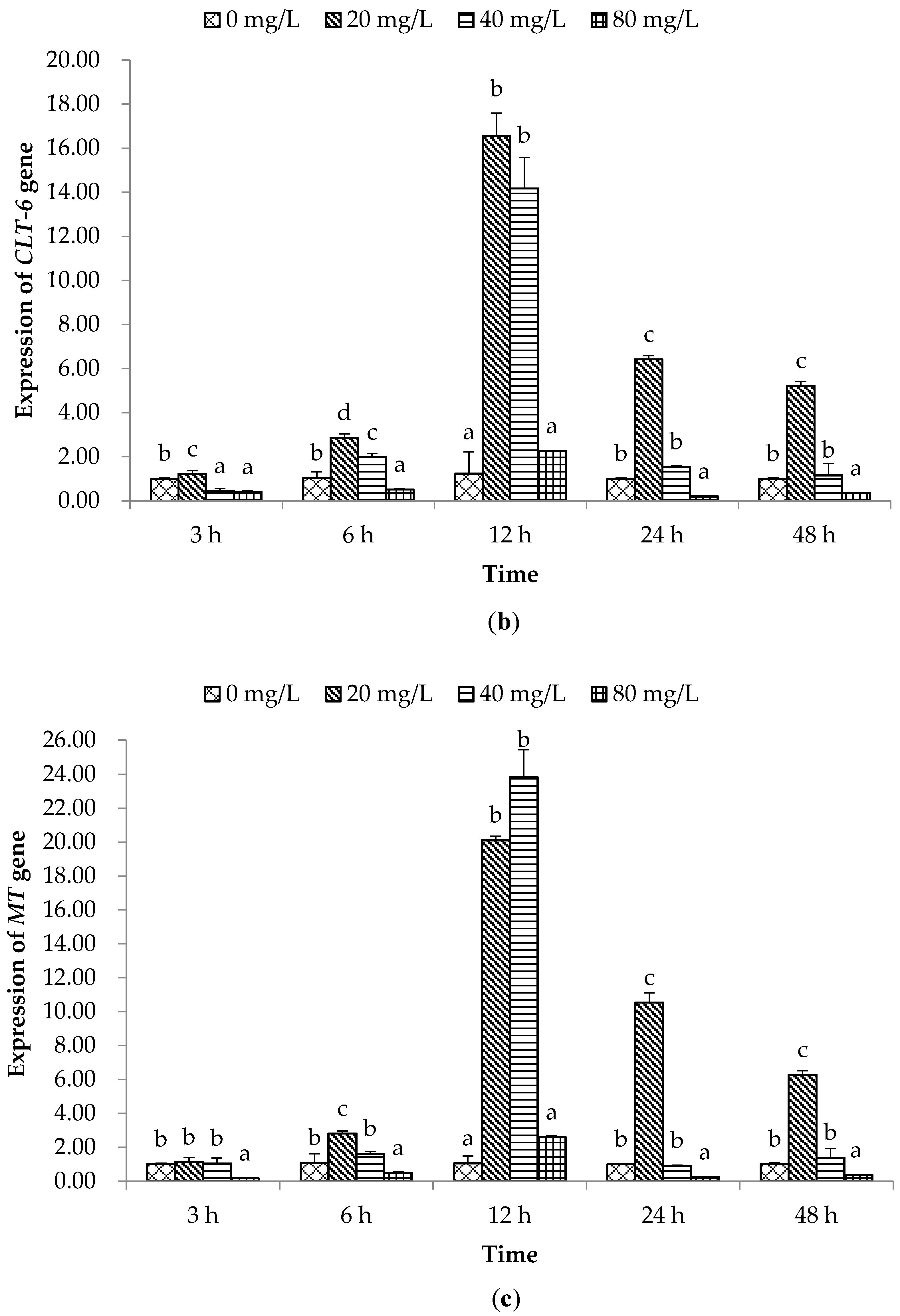
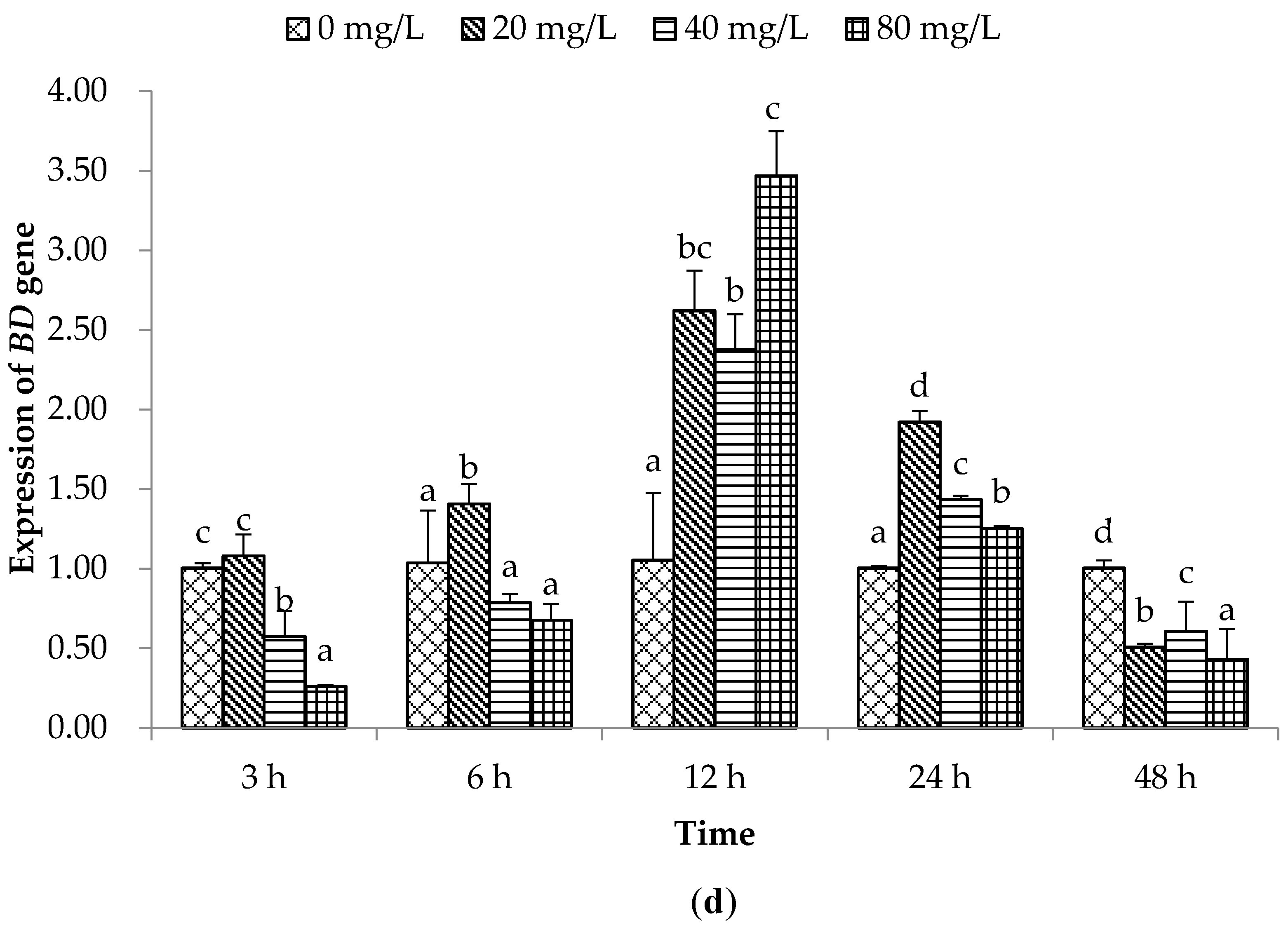
2.2. Expression of Immune Related Genes
3. Materials and Methods
3.1. Palmitoleic Acid
3.2. Animals
3.3. Measurement of Non-Specific Immune Responses
3.4. RNA Extraction and Reverse Transcription
3.5. Real-Time Quantitative PCR Analyses of Gene Expression
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, D.; Zhang, H.; Fu, L.; An, X.; Zhang, B.; Li, Y.; Chen, Z.; Zheng, W.; Yi, L.; Zheng, T. A novel algicide: Evidence of the effect of a fatty acid compound from the marine bacterium, Vibrio sp. BS02 on the harmful dinoflagellate, Alexandrium tamarense. PLoS ONE 2014, 9, e91201. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yang, X.; Zhou, Y.; Lv, J.; Su, J.; Tian, Y.; Zhang, J.; Lin, G.; Zheng, T. An algicidal protein produced by bacterium isolated from the Donghai Sea, China. Harmful Algae 2012, 13, 83–88. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, B.; Zhang, J.; Huang, L.; Lin, J.; Li, X.; Zhou, Y.; Wang, H.; Yang, X.; Su, J.; et al. A marine algicidal actinomycete and its active substance against the harmful algal bloom species Phaeocystis globosa. Appl. Microbiol. Biotechnol. 2013, 97, 9207–9215. [Google Scholar] [CrossRef] [PubMed]
- Gainey, L.G.; Shumway, S.E. A compendium of the responses of bivalve molluscs to toxic dinoflagellate. J. Shellfish Res. 1998, 7, 623–628. [Google Scholar]
- Zheng, T.; Su, J.; Maskaoui, K.; Yu, Z.; Hu, Z.; Xu, J.; Hong, H. Microbial modulation in the biomass and toxin production of a red-tide causing alga. Mar. Pollut. Bull. 2005, 51, 1018–1025. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Zhou, M.; Fu, M.; Yu, R.; Wang, Y.; Li, J. Effects of the dinoflagellate Alexandrium tamarense on early development of the scallop Argopecten irradians concentricus. Aquaculture 2003, 217, 167–178. [Google Scholar] [CrossRef]
- Da Silva, P.M.; Hégaret, H.; Lambert, C.; Wikfors, G.H.; le Goïc, N.; Shumway, S.E.; Soudant, P. Immunological responses of the Manila clam (Ruditapes philippinarum) with varying parasite (Perkinsus olseni) burden, during a long-term exposure to the harmful alga, Karenia selliformis, and possible interactions. Toxicon 2008, 51, 563–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minchin, D. Introductions: Some biological and ecological characteristics of scallops. Aquat. Living Resour. 2003, 16, 521–532. [Google Scholar] [CrossRef]
- Song, L.; Xu, W.; Li, C.; Li, H.; Wu, T.; Xiang, J.; Guo, X. Development of expressed sequence tags from the bay scallop, Argopecten irradians. Mar. Biotechnol. 2006, 8, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Loker, E.S.; Adema, C.M.; Zhang, S.M.; Kepler, T.B. Invertebrate immune systems–not homogeneous, not simple, not well understood. Immunol. Rev. 2004, 198, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Wang, L.; Zhang, H.; Wang, M. The immune system and its modulation mechanism in scallop. Shellfish Immunol. 2015, 46, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Cong, M.; Song, L.; Wang, L.; Zhao, J.; Qiu, L.; Li, L.; Zhang, H. The enhanced immune protection of Zhikong scallop Chlamys farreri on the secondary encounter with Listonella anguillarum. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 151, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pan, L.Q.; Zhang, L.; Miao, J.; Wang, J. Immune responses, ROS generation and the haemocyte damage of scallop Chlamys farreri exposed to Aroclor 1254. Fish Shellfish Immunol. 2009, 26, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Li, J.; Gao, Y.; Mao, Y.; Jiang, Z.; Du, M.; Zhang, Y. Effects of temperature change on physiological and biochemical responses of Yesso scallop, Patinopecten yessoensis. Aquaculture 2016, 451, 463–472. [Google Scholar] [CrossRef]
- Labreuche, Y.; Lambert, C.; Soudant, P.; Boulo, V.; Huvet, A.; Nicolas, J.L. Cellular and molecular hemocyte responses of the Pacific oyster, Crassostrea gigas, following bacterial infection with Vibrio aestuarianus strain 01/32. Microbes Infect. 2006, 8, 2715–2724. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Yang, C.; Wang, L.; Wang, X.; Wang, J.; Yue, F.; Liu, R.; Zhang, H.; Song, L. The protein expression profile in hepatopancreas of scallop Chlamys farreri under heat stress and Vibrio anguillarum challenge. Fish Shellfish Immunol. 2014, 36, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Tomanek, L. Environmental proteomics: Changes in the proteome of marine organisms in response to environmental stress, pollutants, infection, symbiosis, and development. Annu. Rev. Mar. Sci. 2011, 3, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Blaise, C.; Trottier, S.; Gagné, F.; Lallement, C.; Hansen, P.D. Immunocompetence of bivalve hemocytes as evaluated by a miniaturized phagocytosis assay. Environ. Toxicol. 2002, 17, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Hannam, M.L.; Bamber, S.D.; Moody, J.A.; Galloway, T.S.; Jones, M.B. Immune function in the Arctic Scallop, Chlamys islandica, following dispersed oil exposure. Aquat. Toxicol. 2009, 92, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Regoli, F.; Cerrano, C.; Chierici, E.; Chiantore, M.; Bavestrello, G. Seasonal variability of prooxidant pressure and antioxidant adaptation to symbiosis in the Mediterranean demosponge Petrosia ficiformis. Mar. Ecol. Prog. Ser. 2004, 275, 129–137. [Google Scholar] [CrossRef]
- Chen, M.Y.; Yang, H.S.; Delaporte, M.; Zhao, S.J.; Xing, K. Immune responses of the scallop Chlamys farreri after air exposure to different temperatures. J. Exp. Mar. Biol. Ecol. 2007, 345, 52–60. [Google Scholar] [CrossRef]
- Jing, G.; Li, Y.; Xie, L.; Zhang, R. Metal accumulation and enzyme activities in gills and digestive gland of pearl oyster (Pinctada fucata) exposed to copper. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 144, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Song, L.; Li, C.; Zhao, J.; Zhang, H.; Ni, D.; Xu, W. Cloning and characterization of a novel C-type lectin from Zhikong scallop Chlamys farreri. Mol. Immunol. 2007, 44, 722–731. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, L.; Yao, C.; Qiu, L.; Zhang, H.; Zhi, Z.; Song, L. Alternation of immune parameters and cellular energy allocation of Chlamys farreri under ammonia-N exposure and Vibrio anguillarum challenge. Fish Shellfish Immunol. 2012, 32, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.O.; Ren, J.; Liu, J. Responses of antioxidant systems and LPO level to benzo(α)pyrene and benzo(κ)fluoranthene in the haemolymph of the scallop Chlamys ferrari. Environ. Pollut. 2006, 141, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Knoops, B.; Clippe, A.; Bogard, C.; Arsalane, K.; Wattiez, R.; Hermans, C.; Duconseille, E.; Falmagne, P.; Bernard, A. Cloning and characterization of AOEB166, a novel mammalian antioxidant enzyme of the peroxiredoxin family. J. Biol. Chem. 1999, 274, 30451–30458. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, L.; Zhang, S.; Zhang, G. Cloning, genomic structure, and expression analysis of peroxiredoxin V from bay scallop Argopecten irradians. Fish Shellfish Immunol. 2011, 30, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Song, X.; Wang, L.; Kong, P.; Yang, J.; Liu, L.; Qiu, L.; Zhang, Y.; Qiu, L.; Song, L. AiCTL-6, a novel C-type lectin from bay scallop Argopecten irradians with a long C-type lectin-like domain. Fish Shellfish Immunol. 2011, 30, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Gestal, C.; Roch, P.; Renault, T.; Pallavicini, A.; Paillard, C.; Novoa, B.; Oubella, R.; Venier, P.; Figueras, A. Study of diseases and the immune system of bivalves using molecular biology and genomics. Rev. Fish. Sci. 2008, 16, 133–156. [Google Scholar] [CrossRef] [Green Version]
- Miles, A.; Hawksworth, G.; Beattie, J.; Rodilla, V. Induction, regulation, degradation, and biological significance of mammalian metallothioneins. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 35–70. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Song, L.; Ni, D.; Zhang, H.; Liu, W. Alteration of metallothionein mRNA in bay scallop Argopecten irradians under cadmium exposure and bacteria challenge. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 149, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Buico, A.; Cassino, C.; Dondero, F.; Vergani, L.; Osella, D. Radical scavenging abilities of fish MT-A and mussel MT-10 metallothionein isoforms: An ESR study. J. Inorg. Biochem. 2008, 102, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Diamond, G. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000, 8, 402–410. [Google Scholar] [CrossRef]
- Zhao, J.; Song, L.; Li, C.; Ni, D.; Wu, L.; Zhu, L.; Wang, H.; Xu, W. Molecular cloning, expression of a big defensin gene from bay scallop Argopecten irradians and the antimicrobial activity of its recombinant protein. Mol. Immunol. 2007, 44, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Ōyanagui, Y. Reevaluation of assay methods and establishment of kit for superoxide dismutase activity. Anal. Biochem. 1984, 142, 290–296. [Google Scholar] [CrossRef]
- King, J. Practical Clinical Enzymology; Van Nostrand: Princeton, NJ, USA, 1965; p. 363. [Google Scholar]
- Xue, J.; Xu, Y.; Jin, L.; Liu, G.; Sun, Y.; Li, S.; Zhang, J. Effects of traditional Chinese medicine on immune responses in abalone, Haliotis discus hannai Ino. Fish Shellfish Immunol. 2008, 24, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence | Accession No. |
|---|---|---|
| β-actin | F: 5′CAAACAGCAGCCTCCTCGTCA 3′ | AY335441 |
| R: 5′CTGGGCACCTGAACCTTTCGTT 3′ | ||
| PrxV | F: 5′AATCAAGGAGCGGCTGGCA 3′ | HM461987 |
| R: 5′TCAACTTCTCAATCTTCCCGTCAT 3′ | ||
| CTL-6 | F: 5′CAGTTGCTACAGGGTTCG 3′ | GQ202279 |
| R: 5′GGGCGTTATCTGGCTCAT 3′ | ||
| MT | F: 5′AACTTGCTGTAGTGGGAATG 3′ | EU734181 |
| R: 5′AGGCTGGAAACTGCTGTGGT 3′ | ||
| BD | F: 5′CGTGCCATACCCATTGCTTA 3′ | DQ334340 |
| R: 5′ATGATTGTCGTTGCTCCTTGAT 3′ |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chi, C.; Giri, S.S.; Jun, J.W.; Kim, H.J.; Kim, S.G.; Yun, S.; Park, S.C. Effect of the Algaecide Palmitoleic Acid on the Immune Function of the Bay Scallop Argopecten irradians. Molecules 2016, 21, 610. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21050610
Chi C, Giri SS, Jun JW, Kim HJ, Kim SG, Yun S, Park SC. Effect of the Algaecide Palmitoleic Acid on the Immune Function of the Bay Scallop Argopecten irradians. Molecules. 2016; 21(5):610. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21050610
Chicago/Turabian StyleChi, Cheng, Sib Sankar Giri, Jin Woo Jun, Hyoun Joong Kim, Sang Guen Kim, Saekil Yun, and Se Chang Park. 2016. "Effect of the Algaecide Palmitoleic Acid on the Immune Function of the Bay Scallop Argopecten irradians" Molecules 21, no. 5: 610. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21050610