
Activity of Zearalenone in the Porcine Intestinal Tract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Mode of Action in Toxicology
1.2. Low-Dose Hypothesis
1.3. Interactions as an Element of MOA
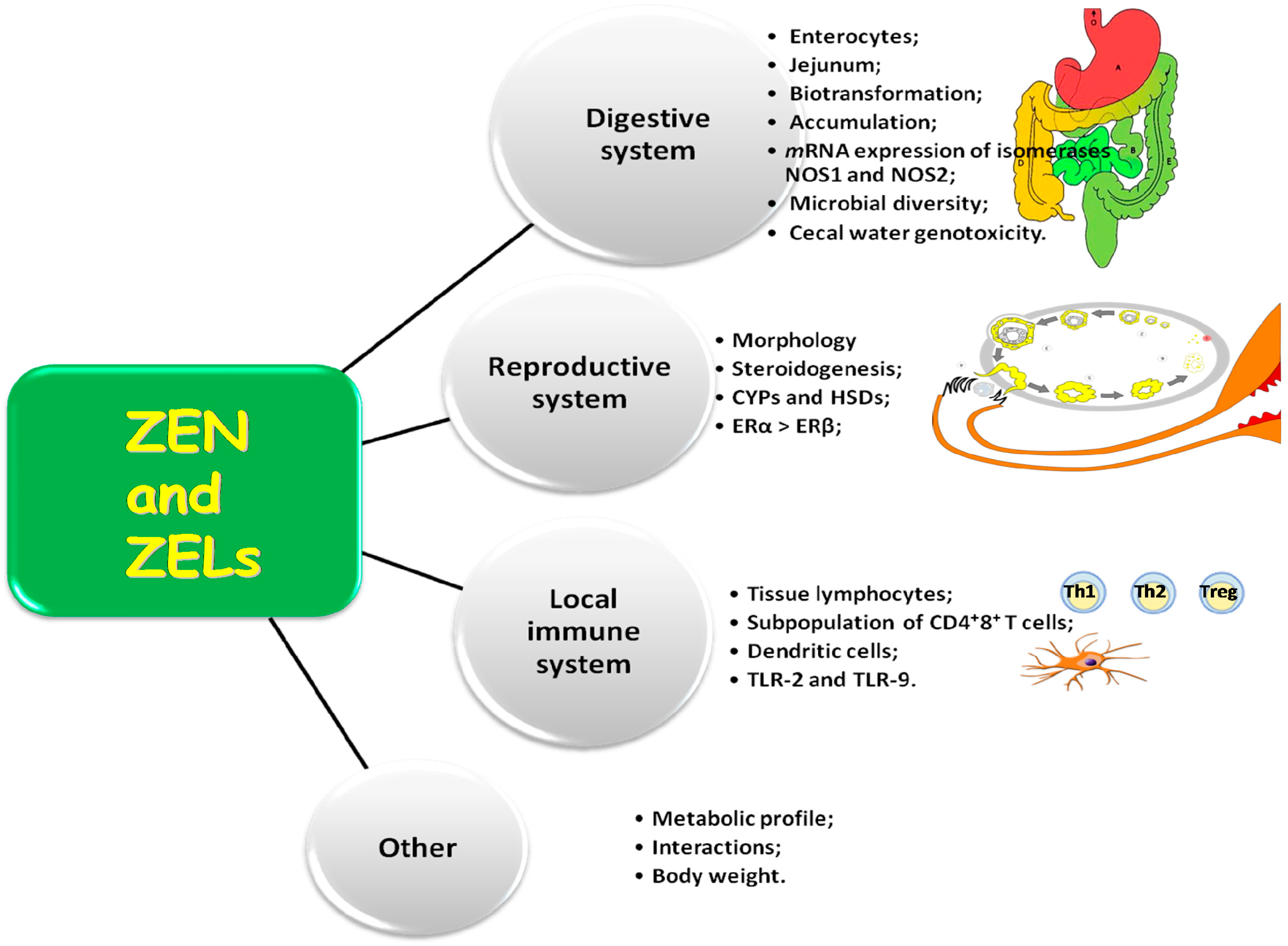
2. Activity of ZEN
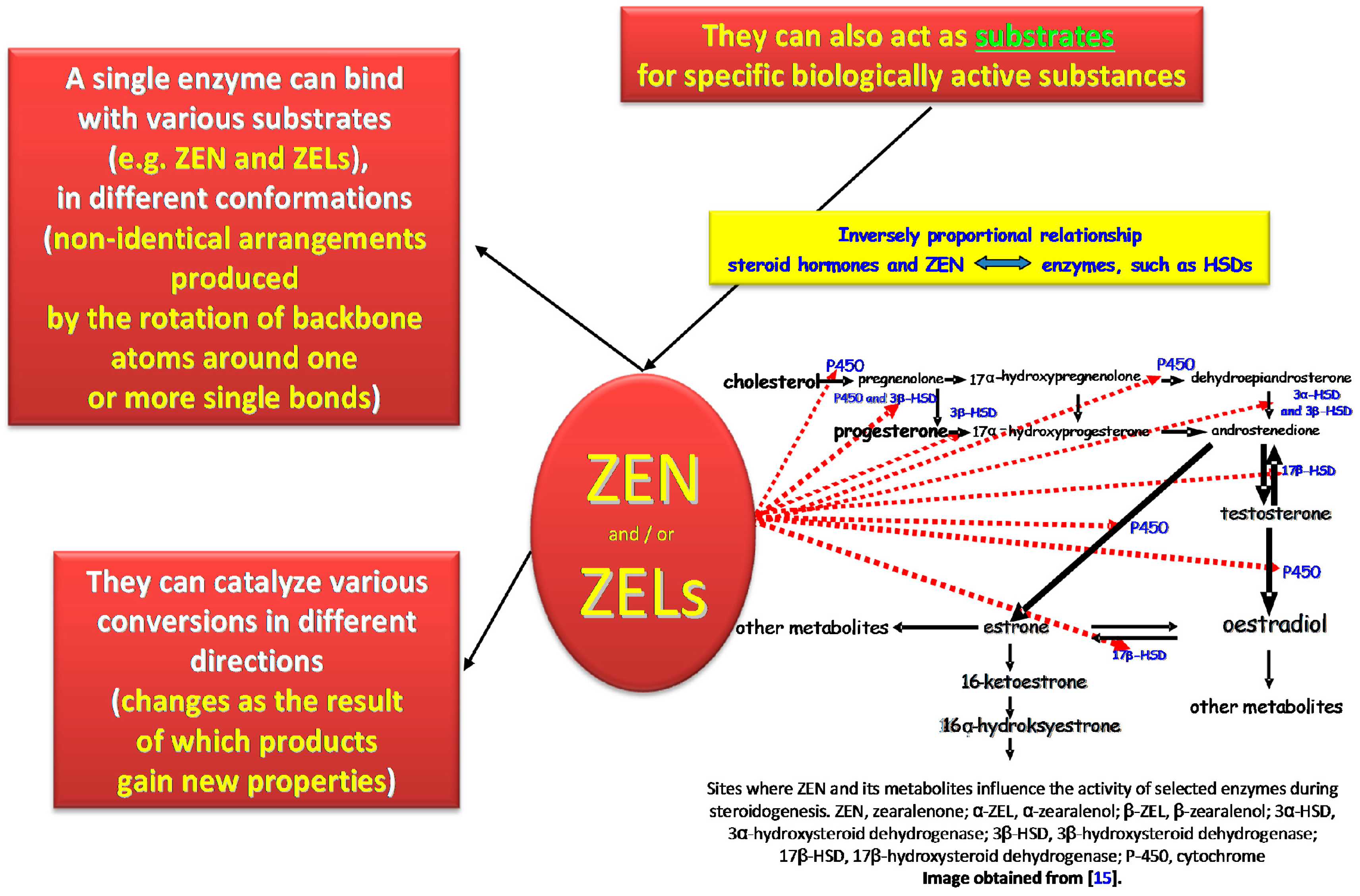
2.1. Multispecificity and Activity of Estrogens
2.1.1. Interactions between Estrogens and Enzymes
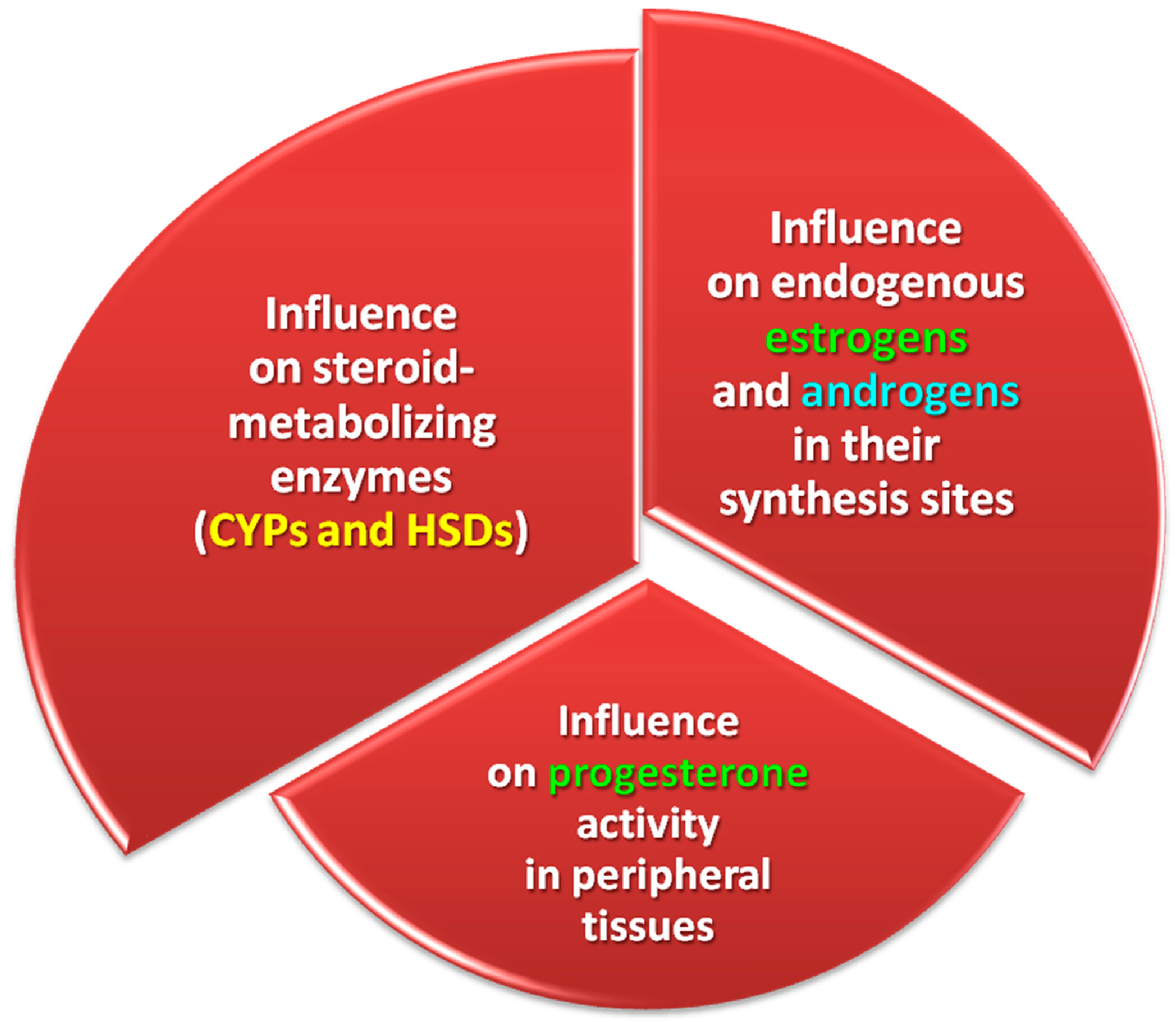
2.1.2. Participation of Selected CYPs and HSDs
2.1.3. The Role of ERs in Estrogen Signaling Pathways
2.2. Morphometry of the Duodenum and Jejunum
2.3. The Carryover Factor and ERs
2.4. Microbiota Diversity in the Intestinal Lumen
2.5. Genotoxicity of Cecal Water
2.6. Expression of Intestinal Nitric Oxide Synthase
2.7. Lymphocyte Subpopulations in Mesenteric Blood Vessels
2.8. Evaluation of the Metabolic Profile
2.9. Body Weight Gains
3. Summary—Key Events Which Play a Major Role in Clinical States
- -
- Disruptions in the proportions of ZEN and carbohydrates which are absorbed in enterocytes in the proximal segment of the intestinal tract;
- -
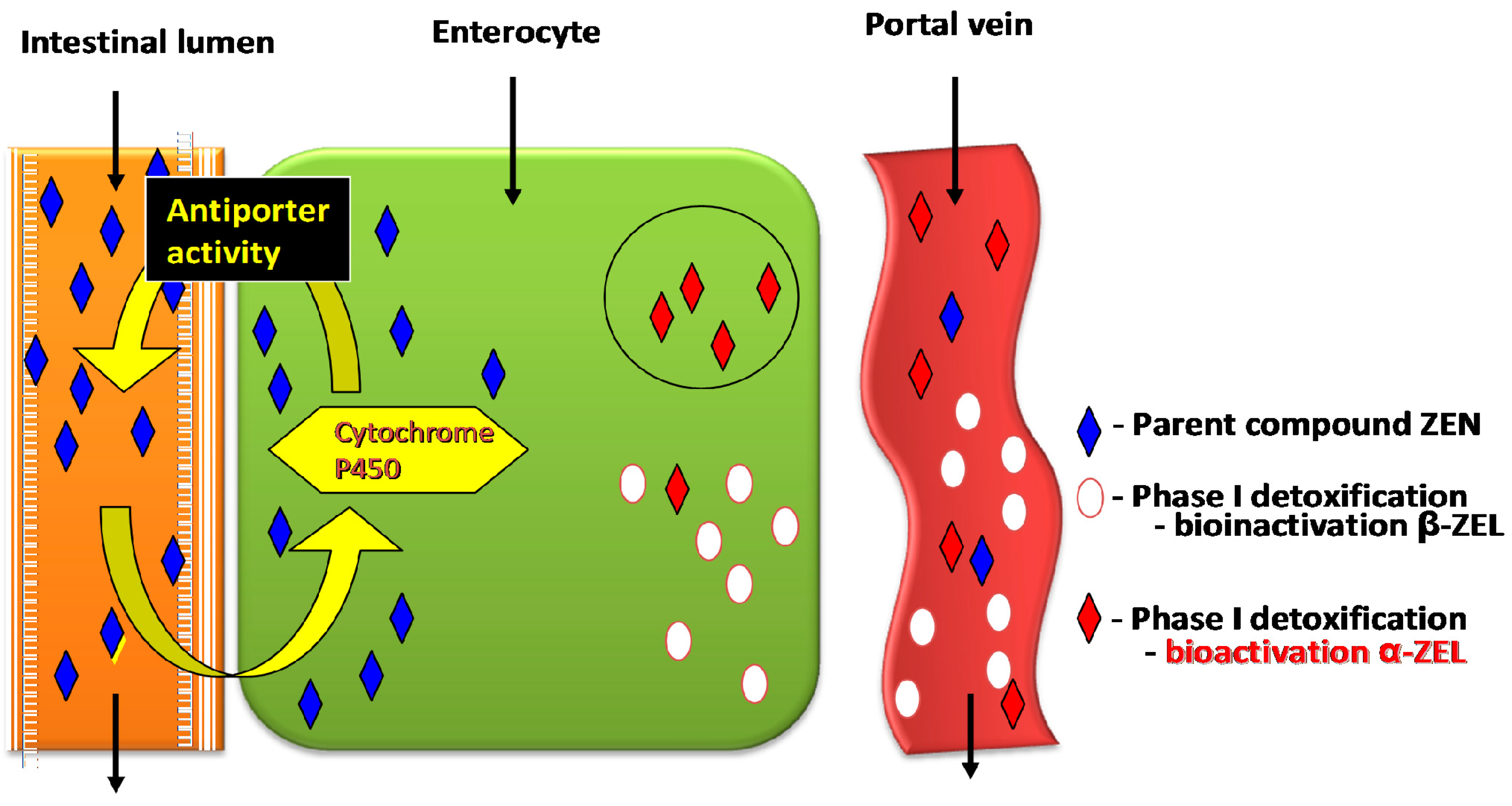
- Absence or limited (below the method detection limit) biotransformation of ZEN in enterocytes after oral administration of the pure parent compound;
- -
- Increase in CF values in selected weeks of exposure in different segments of the intestinal tract, subject to the location and expression of ERβ. In a healthy gut, ERs (mainly ERβ) are found mainly in the duodenum, jejunum and descending colon, and ERβ can modulate the expression of ERα by inhibiting the proliferation of estrogen-dependent cells and promoting apoptosis;
- -
- Activation of ERs due to post-translational modification, which influences non-genomic signal transduction, including by ZEN;
- -
- Defects in ERs and/or inadequate estrogen levels, which influences cellular activity (estrogen signaling pathway);
- -
- Decrease in mRNA expression of both genes controlling NOS isomerases, which slows down intestinal peristalsis, stimulates sphincter contraction, decreases intestinal permeability and inhibits intestinal secretion. The above changes are noted mainly in the distal segment of the digestive tract due. Mycotoxins have bactericidal effects, and they reduce the counts of microbial pathogens, one of the key proinflammatory agents that stimulate NO production;
- -
- Deceleration of gastrointestinal motility, which stimulates antiporter activity in jejunal walls. The above can lead to the accumulation of ZEN in intestinal walls at the beginning of exposure and the inhibition of cell proliferation, including apoptosis (loss of control over proliferation processes);
- -
- Decrease in fecal bacterial counts in the distal colon, which stimulates fecal enzymes β-d-glucosidase and β-d-glucuronidase. Those effects are minimized when LAB concentration is higher. The genotoxicity of cecal water increased in the proximal colon;
- -
- Stimulation of the activity of estrogen-metabolizing enzymes (CYPs and HSDs) and, consequently, endogenous estrogens and androgens at the place of their synthesis, as well as the activity of P4 in peripheral tissues;
- -
- Enhanced proliferative activity of lymphocytes, which is manifested by higher volumetric density of T cells in the jejunal epithelium, an increase in the number of plasma cells in the lamina propria and the number of T cells in lymph nodes, in particular the subpopulations of CD4+8+ T cells in mesenteric venous blood and peripheral blood. The above points to a decrease in the cytotoxic activity of Tc cells, in particular CD4+8+ cells (with cluster of differentiation CD8+), but also CD4−8+ T cells. The local immune system participates in the elimination of subclinical inflammations and/or allergic reactions in the intestines, which is manifested by higher lymphocyte proliferation in various tissues of the digestive tract;
- -
- A metabolic profile analysis revealed an increase in the activity of liver-specific enzymes at the beginning of exposure, followed by a decrease in successive weeks of the experiment. Those fluctuations can probably be attributed to healing processes initiated in response to liver inflammation and reinstatement of liver function. Such discrepancies are noted on a daily basis (balance between catabolism and anabolism), and they neutralize the observed clinical symptoms. The results of the metabolic profile analysis also point to higher demand for oxygen, alleviation of inflammations and considerable loss of energy and protein, which could point to higher feed efficiency, extensive involvement in detoxification or both;
- -
- Increase in body weight gains in pre-pubertal gilts.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hult, K.; Berglund, P. Enzyme promiscuity: Mechanism and applications. Trends Biotechnol. 2007, 25, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Lathe, R.; Kotelevtsev, Y.; Mason, J.I. Steroid promiscuity: Diversity of enzyme action. J. Steroid Biochem. 2015, 151, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.; Meyer, M.R. Nicolaus Copernicus and the rapid vascular responses to aldosterone. Trends Endocrinol. Metab. 2015, 26, 396–398. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Accensi, F.; Beauchamp, E.; Cossalter, A.-M.; Callu, P.; Grosjean, F.; Oswald, I.P. Ingestion of deoxynivalenol (DON) contaminated feed alters the pig vaccinal immune responses. Toxicol. Lett. 2008, 177, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.C.; Madec, S.; Coton, E.; Hymery, N. Natural co-occurrence of mycotoxins in foods and feeds and their in vitro combined toxicological effects. Toxins 2016, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Stuper-Szablewska, K.; Szablewski, T.; Buszko, M.; Perkowski, J. Changes in contents of trichothecenes during commercial grain milling. LWT Food Sci. Technol. 2016, 69, 55–58. [Google Scholar] [CrossRef]
- Wielogórska, E.; Elliott, C.T.; Danaher, M.; Connolly, L. Validation and application of a reporter gene assay for the determination of estrogenic endocrine disruptor activity in milk. Food Chem. Toxicol. 2014, 69, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Zielonka, Ł.; Dąbrowski, M.; Mróz, M.; Gajęcki, M. The effect of low doses of zearalenone and its metabolites on progesterone and 17β-estradiol concentrations in blood and body weights of pre-pubertal female Beagle dogs. Toxicon 2013, 76, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Flores-Flores, M.E.; Lizarraga, E.; de Cerain, A.L.; González-Peñas, E. Presence of mycotoxins in animal milk: A review. Food Control 2015, 53, 163–176. [Google Scholar] [CrossRef]
- Broekaert, N.; Devreese, M.; de Baere, S.; de Backer, P.; Croubels, S. Modified Fusarium mycotoxins unmasked: From occurrence in cereals to animal and human excretion. Food Chem. Toxicol. 2015, 80, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Pistol, G.C.; Neagoe, I.V.; Calin, L.; Taranu, I. Effects of zearalenone on oxidative stress and inflammation in weanling piglets. Food Chem. Toxicol. 2013, 58, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.M.; Wood, K.M.; McEwen, P.L.; Smith, T.K.; Mandell, I.B.; Yannikouris, A.; Swanson, K.C. Effects of feeding corn naturally contaminated with Fusarium mycotoxins and/or a modified yeast cell wall extract on the performance, immunity and carcass characteristics of grain-fed veal calves. Anim. Feed Sci. Technol. 2010, 159, 27–34. [Google Scholar] [CrossRef]
- Dunbar, B.; Patel, M.; Fahey, J.; Wira, C. Endocrine control of mucosal immunity in the female reproductive tract: Impact of environmental disruptors. Mol. Cell. Endocrinol. 2012, 354, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Alm, H.; Brüssow, K.-P.; Torner, H.; Vanselow, J.; Tomek, W.; Dänicke, S.; Tiemann, U. Influence of Fusarium-toxin contaminated feed on initial quality and meiotic competence of gilt oocytes. Reprod. Toxicol. 2006, 22, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. The effect of low monotonic doses of zearalenone on selected reproductive tissues in pre-pubertal female dogs—A review. Molecules 2015, 20, 20669–20687. [Google Scholar] [CrossRef] [PubMed]
- Gajęcki, M. The Effect of Experimentally Induced Fusarium Mycotoxicosis on Selected Diagnostic and Morphological Parameters of the Porcine Digestive Tract; Final Report; Development Project NR12-0080-10; National Centre for Research and Development: Warsaw, Poland, 2013; pp. 1–180.
- Escrivá, L.; Font, G.; Manyes, L. In vivo toxicity studies of fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowski, M.; Obremski, K.; Gajęcka, M.; Gajęcki, M.; Zielonka, Ł. Changes in the subpopulations of porcine peripheral blood lymphocytes induced by exposure to low doses of zearalenone (ZEN) and deoxynivalenol (DON). Molecules 2016, 21, 557. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of mycotoxin on immune response and consequences for pig health. Anim. Nutr. 2016, 2, 63–68. [Google Scholar] [CrossRef]
- Taranu, I.; Braicu, C.; Marin, D.E.; Pistol, G.C.; Motiu, M.; Balacescu, L.; Neagoe, I.B.; Burlacu, R. Exposure to zearalenone mycotoxin alters in vitro porcine intestinal epithelial cells by differential gene expression. Toxicol. Lett. 2015, 232, 310–325. [Google Scholar] [CrossRef] [PubMed]
- Przybylska-Gornowicz, B.; Tarasiuk, M.; Lewczuk, B.; Prusik, M.; Ziółkowska, N.; Zielonka, Ł.; Gajęcki, M.; Gajęcka, M. The effects of low doses of two Fusarium toxins, zearalenone and deoxynivalenol, on the pig jejunum. A light and electron microscopic study. Toxins 2015, 7, 4684–4705. [Google Scholar] [CrossRef] [PubMed]
- Lewczuk, B.; Przybylska-Gornowicz, B.; Gajęcka, M.; Targońska, K.; Ziółkowska, N.; Prusik, M.; Gajęcki, M. Histological structure of duodenum in gilts receiving low doses of zearalenone and deoxynivalenol in feed. Exp. Toxicol. Pathol. 2016, 68, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M.; Fantini, J. Some food-associated mycotoxins as potential risk factors in humans predisposed to chronic intestinal inflammatory diseases. Toxicon 2010, 56, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Joa, H.; Konga, C.; Song, M.; Kim, B.G. Effects of dietary deoxynivalenol and zearalenone on apparent ileal digestibility of amino acids in growing pigs. Anim. Feed Sci. Technol. 2016, 219, 77–82. [Google Scholar] [CrossRef]
- Tarasiuk, M. The Effect of Low Doses of Zearalenone and Deoxynivalenol on the Jejunal Mucosa, Metabolic Profile and Body Weight of Pre-Pubertal Gilts. Ph.D. Thesis, University of Warmia and Mazury in Olsztyn, Olsztyn, Poland, 2015. [Google Scholar]
- Hartung, T.; McBride, M. Food for thought … on mapping the human toxome. ALTEX 2011, 28, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Embry, M.R.; Bachman, A.N.; Bell, D.R.; Boobis, A.R.; Cohen, S.M.; Dellarco, M.; Dewhurst, I.C.; Doerrer, N.G.; Hines, R.N.; Moretto, A.; et al. Risk assessment in the 21st century: Roadmap and matrix. Crit. Rev. Toxicol. 2014, 44, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Pastoor, T.P.; Bachman, A.N.; Bell, D.R.; Cohen, S.M.; Dellarco, M.; Dewhurst, I.C.; Doe, J.E.; Doerrer, N.G.; Embry, M.R.; Hines, R.N.; et al. A 21st century roadmap for human health risk assessment. Crit. Rev. Toxicol. 2014, 44, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Simon, T.W.; Simons, S.S., Jr.; Preston, R.J.; Boobis, A.R.; Cohen, S.M.; Doerrer, N.G.; Fenner-Crisp, P.A.; McMullin, T.S.; McQueen, C.A.; Rowlands, J.C.; et al. The use of mode of action information in risk assessment: Quantitative key events/dose-response framework for modeling the dose-response for key events. Crit. Rev. Toxicol. 2014, 44 (Suppl. 3), 17–43. [Google Scholar] [CrossRef] [PubMed]
- Fleck, S.C.; Churchwell, M.I.; Doerge, D.R.; Teeguarden, J.G. Urine and serum biomonitoring of exposure to environmental estrogens II: Soy isoflavones and zearalenone in pregnant women. Food Chem. Toxicol. 2016, 95, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, L.M.; Moeser, A.J.; Blikslager, A.T. Porcine models of digestive disease: The future of large animal translational research. Transl. Res. 2015, 166, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Demaegdt, H.; Daminet, B.; Evrard, A.; Scippo, M.-L.; Muller, M.; Pussemier, L.; Callebaut, A.; Vandermeiren, K. Endocrine activity of mycotoxins and mycotoxin mixtures. Food Chem. Toxicol. 2016, 96, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Patlewicz, G.; Simon, T.; Goyak, K.; Phillips, R.D.; Rowlands, J.C.; Seidel, S.; Becker, R.A. Use and validation of HT/HC assays to support 21st century toxicity evaluations. Regul. Toxicol. Pharmacol. 2013, 65, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Rybarczyk, L.; Zwierzchowski, W.; Jakimiuk, E.; Zielonka, Ł.; Obremski, K.; Gajęcki, M. The effect of experimental, long-term exposure to low-dose zearalenone mycotoxicosis on the histological condition of ovaries in sexually immature gilts. Theriogenology 2011, 75, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Zachariasova, M.; Dzumana, Z.; Veprikova, Z.; Hajkovaa, K.; Jiru, M.; Vaclavikova, M.; Zachariasova, A.; Pospichalova, M.; Florian, M.; Hajslova, J. Occurrence of multiple mycotoxins in European feeding stuffs, assessment of dietary intake by farm animals. Anim. Feed Sci. Technol. 2014, 193, 124–140. [Google Scholar] [CrossRef]
- Calabrese, E.J. Paradigm lost, paradigm found: The re-emergence of hormesis as a fundamental dose response model in the toxicological sciences. Environ. Pollut. 2005, 138, 378–411. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyński, L.; Fornalski, K.W. Hormesis—Natural phenomenon of answer of organism on stress. In Proceeding of the 7th International Scientific Conference on Veterinary Feed Hygiene—The Effects of Mycotoxins on Gastrointestinal Function, Olsztyn, Poland, 23–24 September 2011; pp. 6–14.
- Kramer, H.J.; van den Ham, W.A.; Slob, W.; Pieters, M.N. Conversion factors estimating indicative chronic no-observed-adverse-effect levels from short-term toxicity data. Regul. Toxicol. Pharmacol. 1996, 23, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Stopa, E.; Babińska, I.; Zielonka, Ł.; Gajęcki, M.; Gajęcka, M. Immunohistochemical evaluation of apoptosis and proliferation in the mucous membrane of selected uterine regions in pre-pubertal bitches exposed to low doses of zearalenone. Pol. J. Vet. Sci. 2016, 19, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, Ł.; Jakimiuk, E.; Obremski, K.; Gajęcka, M.; Dąbrowski, M.; Gajęcki, M. An evaluation of the proliferative activity of immunocompetent cells in the jejunal and iliac lymph nodes of prepubertal female wild boars diagnosed with mixed mycotoxicosis. Bull. Vet. Inst. Pulawy 2015, 59, 197–203. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.; Lee, D.-H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed]
- Hickey, G.L.; Craig, P.S.; Luttik, R.; de Zwart, D. On the quantification of intertest variability in ecotoxicity data with application to species sensitivity distributions. Environ. Toxicol. Chem. 2012, 31, 1903–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waśkiewicz, A.; Beszterda, M.; Kostecki, M.; Zielonka, Ł.; Goliński, P.; Gajęcki, M. Deoxynivalenol in the gastrointestinal tract of immature gilts under per os toxin application. Toxins 2014, 6, 973–987. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M.; Śliżewska, K.; Nowak, A.; Zielonka, Ł.; Żakowska, Z.; Gajęcka, M.; Gajęcki, M. The effect of experimental Fusarium mycotoxicosis on microbiota diversity in porcine ascending colon contents. Toxins 2014, 6, 2064–2081. [Google Scholar] [CrossRef] [PubMed]
- Nowak, A.; Śliżewska, K.; Gajęcka, M.; Piotrowska, M.; Żakowska, Z.; Zielonka, Ł.; Gajęcki, M. The genotoxicity of caecal water from gilts following experimentally induced Fusarium mycotoxicosis. Vet. Med. 2015, 60, 133–140. [Google Scholar] [CrossRef]
- Zielonka, Ł.; Waśkiewicz, A.; Beszterda, M.; Kostecki, M.; Dąbrowski, M.; Obremski, K.; Goliński, P.; Gajęcki, M. Zearalenone in the intestinal tissues of immature gilts exposed per os to mycotoxins. Toxins 2015, 7, 3210–3223. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.Y.M.; Turner, P.C.; El-Nezami, H. Individual and combined cytotoxic effects of Fusarium toxins (deoxynivalenol, nivalenol, zearalenone and fumonisins B1) on swine jejunal epithelial cells. Food Chem. Toxicol. 2013, 57, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Gajęcki, M.; Gajęcka, M.; Zielonka, Ł.; Jakimiuk, E.; Obremski, K. Zearalenone as a potential allergen in the alimentary tract—A review. Pol. J. Food Nutr. Sci. 2006, 15, 263–268. [Google Scholar]
- McLaughlin, J.; Lambert, D.; Padfield, P.J.; Burt, J.P.; O’Neill, C.A. The mycotoxin patulin, modulates tight junctions in caco-2 cells. Toxicol. In Vitro 2009, 23, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Bakhru, S.H.; Furtado, S.; Morello, A.P.; Mathiowitz, E. Oral delivery of proteins by biodegradable nanoparticles. Adv. Drug Deliv. Rev. 2013, 65, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Tulayakul, P.; Li, J.Y.; Dong, K.S.; Manabe, N.; Kumagai, S. Metabolic conversion of zearalenone to α-zearalenol by goat tissues. J. Vet. Med. Sci. 2010, 72, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Cavret, S.; Lecoeur, S. Fusariotoxin transfer in animal. Food Chem. Toxicol. 2006, 44, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Bracarense, A.P.; Lucioli, J.; Grenier, B.; Pacheco, G.D.; Moll, W.D.; Schatzmayr, G.; Oswald, I.P. Chronic ingestion of deoxynivalenol and fumonisin, alone or in interaction, induces morphological and immunological changes in the intestine of piglets. Br. J. Nutr. 2012, 107, 1776–1786. [Google Scholar] [CrossRef] [PubMed]
- Diczfalusy, U.; Bjorkhem, I. Still another activity by the highly promiscuous enzyme CYP3A4: 25-hydroxylation of cholesterol. J. Lipid Res. 2011, 52, 1447–1449. [Google Scholar] [CrossRef] [PubMed]
- Hulce, J.J.; Cognetta, A.B.; Niphakis, M.J.; Tully, S.E.; Cravatt, B.F. Proteome-wide mapping of cholesterol-interacting proteins in mammalian cells. Nat. Methods 2013, 10, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Lathe, R.; Kotelevtsev, Y. Steroid signaling: Ligand-binding promiscuity molecular symmetry, and the need for gating. Steroids 2014, 82, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Otrocka-Domagała, I. Immunocytochemical expression of 3β- and 17β-hydroxysteroid dehydrogenase in bitch ovaries exposed to low doses of zearalenone. Pol. J. Vet. Sci. 2013, 16, 55–62. [Google Scholar] [PubMed]
- Maćkowiak, B.; Wang, H. Mechanisms of xenobiotic receptor activation: Direct vs. indirect. BBA Gene Regul. Mech. 2016, 1859, 1130–1140. [Google Scholar] [CrossRef] [PubMed]
- Brzuzan, P.; Woźny, M.; Wolińska, L.; Piasecka, A.; Florczyk, M.; Jakimiuk, E.; Góra, M.; Łuczyński, M.K.; Gajęcki, M. MicroRNA expression profiles in liver and colon of sexually immature gilts after exposure to Fusarium mycotoxins. Pol. J. Vet. Sci. 2015, 18, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Tawfik, D.S. Messy biology and the origins of evolutionary innovations. Nat. Chem. Biol. 2010, 6, 692–696. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.K.; Noel, J.P. The remarkable pliability and promiscuity of specialized metabolism. Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, Ł.; Gajęcka, M.; Rozicka, A.; Dąbrowski, M.; Żmudzki, J.; Gajęcki, M. The Effect of environmental mycotoxins on selected ovarian tissue fragments of multiparous female wild boars at the beginning of astronomical winter. Toxicon 2014, 89, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Kradolfer, D.; Flöter, V.L.; Bick, J.T.; Fürst, R.W.; Rode, K.; Brehm, R.; Henning, H.; Waberski, D.; Bauersachs, S.; Ulbrich, S.E. Epigenetic effects of prenatal estradiol-17b exposure on the reproductive system of pigs. Mol. Cell. Endocrinol. 2016, 430, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Luu-The, V. Assessment of steroidogenesis and steroidogenic enzyme functions. J. Steroid Biochem. 2013, 137, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, J.V.; Sundaramoorthy, M.; Zhao, B.; Waterman, M.R.; Stegeman, J.J.; Lamb, D.C. Genetic and structural analyses of cytochrome P450 hydroxylases in sex hormone biosynthesis: Sequential origin and subsequent coevolution. Mol. Phylogenet. Evol. 2016, 94, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Warmerdam, E.G.; Visser, M.; Coelingh Bennink, H.J.; Groen, M. A new route of synthesis of estetrol. Climacteric 2008, 11, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Aidoo-Gyamfi, K.; Cartledge, T.; Shah, K.; Ahmed, S. Estrone sulfatase and its inhibitors. Anticancer Agents Med. Chem. 2009, 9, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Bondesson, M.; Hao, R.; Lin, C.Y.; Williams, C.; Gustafsson, J.-Å. Estrogen receptor signaling during vertebrate development. BBA Gene Regul. Mech. 2015, 1849, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Arlt, W.; Martens, J.W.; Song, M.; Wang, J.T.; Auchus, R.J.; Miller, W.L. Molecular evolution of adrenarche: Structural and functional analysis of p450c17 from four primate species. Endocrinology 2002, 143, 4665–4672. [Google Scholar] [CrossRef] [PubMed]
- Greaves, R.F.; Jevalikar, G.; Hewitt, J.K.; Zacharin, M.R. A guide to understanding the steroid pathway: New insights and diagnostic implications. Clin. Biochem. 2014, 47, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Snawder, J.E.; Lipscomb, J.C. Interindividual variance of cytochrome P450 forms in human hepatic microsomes: Correlation of individual forms with xenobiotics metabolism and implications in risk assessment. Regul. Toxicol. Pharmacol. 2000, 32, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Jakimiuk, E.; Zielonka, Ł.; Obremski, K.; Gajęcki, M. The biotransformation of chosen mycotoxins. Pol. J. Vet. Sci. 2009, 12, 293–303. [Google Scholar] [PubMed]
- Sayers, E.W.; Barrett, T.; Benson, D.A.; Bolton, E.; Bryant, S.H.; Canese, K.; Chetvernin, V.; Church, D.M.; Dicuccio, M.; Federhen, S.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2012, 40, D13–D25. [Google Scholar] [CrossRef] [PubMed]
- Kisiela, M.; Skarka, A.; Ebert, B.; Maser, E. Hydroxysteroid dehydrogenases (HSDs) in bacteria—A bioinformatic perspective. J. Steroid Biochem. 2012, 129, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Payne, D.W.; Talalay, P. Isolation of novel microbial 3α-, 3β-, and 17-β hydroxysteroid dehydorgenases. J. Biol. Chem. 1985, 260, 13648–13655. [Google Scholar] [PubMed]
- Drasar, B.S.; Hill, M.J. Intestinal bacteria and cancer. Am. J. Clin. Nutr. 1972, 25, 1399–1404. [Google Scholar] [PubMed]
- Clark, D.T.; Soory, M. The metabolism of cholesterol and certain hormonal steroids by Treponema denticola. Steroids 2006, 71, 352–363. [Google Scholar] [CrossRef] [PubMed]
- Frizzell, C.; Ndossi, D.; Verhaegen, S.; Dahl, E.; Eriksen, G.; Sørlie, M.; Ropstad, E.; Muller, M.; Elliott, C.T.; Connolly, L. Endocrine disrupting effects of zearalenone, alpha- and beta-zearalenol at the level of nuclear receptor binding and steroidogenesis. Toxicol. Lett. 2011, 206, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Endo, S.; Miyagi, N.; Matsunaga, T.; Hara, A.; Ikari, A. Human dehydrogenase/reductase (SDR family) member 11 is a novel type of 17β-hydroxysteroid dehydrogenase. Biochem. Biophys. Res. Commun. 2016, 472, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Hueza, I.M.; Raspantini, P.C.F.; Raspantini, L.E.R.; Latorre, A.O.; Górniak, S.L. Zearalenone, an estrogenic mycotoxin, is an immunotoxic compound. Toxins 2014, 6, 1080–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchais-Oberwinkler, S.; Henn, C.; Moller, G.; Klein, T.; Negri, M.; Oster, A.; Spadaro, A.; Werth, R.; Wetzel, M.; Xu, K.; et al. 17β-Hydroxysteroid dehydrogenases (17β-HSDs) as therapeutic targets: Protein structures, functions, and recent progress in inhibitor development. J. Steroid Biochem. 2011, 125, 66–82. [Google Scholar] [CrossRef] [PubMed]
- Huhtinen, K.; Stahle, M.; Perheentupa, A.; Poutanen, M. Estrogen biosynthesis and signaling in endometriosis. Mol. Cell Endocrinol. 2012, 358, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, R.D.; Brisken, C. Paracrine signaling by progesterone. Mol. Cell Endocrinol. 2012, 357, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Gajęcki, M.; Gajęcka, M.; Jakimiuk, E.; Zielonka, L.; Obremski, K. Zearalenone—Undesirable substance. In Mycotoxins in Food, Feed and Bioweapons; Rai, M., Ajit, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 131–144. [Google Scholar]
- Gajęcka, M. The effect of low-dose experimental zearalenone intoxication on the immunoexpression of estrogen receptors in the ovaries of pre-pubertal bitches. Pol. J. Vet. Sci. 2012, 15, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Warnmark, A.; Treuter, E.; Wright, A.P.; Gustafsson, J.Å. Activation functions 1 and 2 of nuclear receptors: Molecular strategies for transcriptional activation. Mol. Endocrinol. 2003, 17, 1901–1909. [Google Scholar] [CrossRef] [PubMed]
- López-Calderero, I.; Carnero, A.; Astudillo, A.; Palacios, J.; Chaves, M.; Benavent, M.; Limón, M.L.; Garcia-Carbonero, R. Prognostic relevance of estrogen receptor-α Ser167 phosphorylation in stage II-III colon cancer patients. Hum. Pathol. 2014, 45, 2437–2446. [Google Scholar] [CrossRef] [PubMed]
- Zuloaga, D.G.; Zuloaga, K.L.; Hinds, L.R.; Carbone, D.L.; Handa, R.J. Estrogen receptor β expression in the mouse forebrain: Age and sex differences. J. Comp. Neurol. 2014, 522, 358–371. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.F.; Tan, X.J.; Dai, Y.B.; Krishnan, V.; Warner, M.; Gustafsson, J.Å. Targeting estrogen receptor b in microglia and T cells to treat experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. USA 2013, 110, 3543–3548. [Google Scholar] [CrossRef] [PubMed]
- Warner, M.; Gustafsson, J.-A. DHEA—A precursor of ERβ ligands. J. Steroid Biochem. 2015, 145, 245–247. [Google Scholar] [CrossRef] [PubMed]
- Wada-Hiraike, O.; Imamov, O.; Hiraike, H.; Hultenby, K.; Schwend, T.; Omoto, Y.; Warner, M.; Gustafsson, J.Å. Role of estrogen receptor beta in colonic Epithelium. Proc. Natl. Acad. Sci. USA 2006, 103, 2959–2964. [Google Scholar] [CrossRef] [PubMed]
- Papoutsi, Z.; Zhao, C.; Putnik, M.; Gustafsson, J.; Dahlman-Wright, K. Binding of estrogen receptor alpha/beta heterodimers to chromatin in MCF-7 cells. J. Mol. Endocrinol. 2009, 43, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Takemura, H.; Shim, J.Y.; Sayama, K.; Tsubura, A.; Zhu, B.T.; Shimoi, K. Characterization of the estrogenic activities of zearalenone and zeranol in vivo and in vitro. J. Steroid Biochem. 2007, 103, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.; Simon, S.; Chae, K.; Metzler, M.; Korach, K.S. Phytoestrogens and their human metabolites show distinct agonistic and antagonistic properties on estrogen receptor α (ERa) and ERβ in human cells. Toxicol. Sci. 2004, 80, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.R.; Pietras, R.J. Estrogen receptors outside the nucleus in breast cancer. Breast Cancer Res. Treat. 2008, 108, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Barton, M. Position paper: The membrane estrogen receptor GPER—Clues and questi. Steroids 2012, 77, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Barton, M. Not lost in translation: Emerging clinical importance of the G protein-coupled estrogen receptor GPER. Review Article. Steroids 2016, 111, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Bodiga, V.L.; Boindala, S.; Putcha, U.; Subramaniam, K.; Manchala, R. Chronic low intake of protein or vitamins increases the intestinal epithelial cell apoptosis in Wistar/NIN rats. Nutrition 2005, 21, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Taranu, I.; Burlacu, R.; Manda, G.; Motiu, M.; Neagoe, I.; Dragomir, C.; Stancu, M.; Calin, L. Effects of zearalenone and its derivatives on porcine immune response. Toxicol. In Vitro 2011, 25, 1981–1988. [Google Scholar] [CrossRef] [PubMed]
- Solhaug, A.; Karlsøen, L.M.; Holme, J.A.; Kristoffersen, A.B.; Eriksen, G.S. Immunomodulatory effects of individual and combined mycotoxins in the THP-1 cell line. Toxicol. In Vitro 2016, 36, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Obremski, K. The effect of in vivo exposure to zearalenone on cytokine secretion by Th1 and Th2 lymphocytes in porcine Peyer’s patches after in vitro stimulation with LPS. Pol. J. Vet. Sci. 2014, 17, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Knudsen, K.E.B.; Lærke, H.N.; Ingerslev, A.K.; Hedemann, M.S.; Nielsen, T.S.; Theil, P.K. Carbohydrates in pig nutrition—Recent advances. J. Anim. Sci. 2016, 94, 1–11. [Google Scholar] [CrossRef]
- Lupescu, A.; Bissinger, R.; Jilani, K.; Lang, F. In vitro induction of erythrocyte phosphatidyloserine translocation by the natural Naphthoquinone Shikonin. Toxins 2014, 6, 1559–1574. [Google Scholar] [CrossRef] [PubMed]
- Avantaggiato, G.; Havenaar, R.; Visconti, A. Evaluation of the intestinal absorption of deoxynivalenol and nivalenol by an in vitro gastrointestinal model, and the binding efficacy of activated carbon and other adsorbent materials. Food Chem. Toxicol. 2004, 42, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M. From the gut to the brain: Journey and pathophysiological effects of the food-associated trichothecene mycotoxin deoxynivalenol. Toxins 2013, 5, 784–820. [Google Scholar] [CrossRef] [PubMed]
- Kollarczik, B.; Gareis, M.; Hanelt, M. In vitro transformation of the Fusarium mycotoxins deoxynivalenol and zearalenone by the normal gut microflora of pigs. Nat. Toxins 1994, 2, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Stopa, E.; Tarasiuk, M.; Zielonka, Ł.; Gajęcki, M. The expression of type-1 and type-2 nitric oxide synthase in selected tissues of the gastrointestinal tract during mixed mycotoxicosis. Toxins 2013, 5, 2281–2292. [Google Scholar] [CrossRef] [PubMed]
- Lucioli, J.; Pinton, P.; Callu, P.; Laffitte, J.; Grosjean, F.; Kolf-Clauw, M.; Oswald, I.P.; Bracarense, A.P. The food contaminant deoxynivalenol activates the mitogen activated protein kinases in the intestine: Interest of ex vivo models as an alternative to in vivo experiments. Toxicon 2013, 66, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Sergent, T.; Ribonnet, L.; Kolosova, A.; Garsou, S.; Schaut, A.; de Saeger, S.; van Peteghem, C.; Larondelle, Y.; Pussemier, L.; Schneider, Y.J. Molecular and cellular effects of food contaminants and secondary plant components and their plausible interactions at the intestinal level. Food Chem.Toxicol. 2008, 46, 813–841. [Google Scholar] [CrossRef] [PubMed]
- Cutolo, M.; Sulli, A.; Straub, R.H. Estrogen metabolism and autoimmunity. Autoimmun. Rev. 2012, 11, A460–A464. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Lin, P.; Wang, N.; Yang, D.; Wen, X.; Zhou, D.; Wang, A.; Jin, Y. Herp depletion inhibits zearalenone-induced cell death in RAW 264.7 macrophages. Toxicol. In Vitro 2016, 32, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Silva-Campa, E.; Mata-Haro, V.; Mateu, E.; Hernández, J. Porcine reproductive and respiratory syndrome virus induces CD4+CD8+CD25+Foxp3+ regulatory T cells (Tregs). Virology 2012, 430, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.-J.; Feng, C.-C.; Liu, Q.; Zhang, L.-Y.; Dong, X.; Liu, Z.-L.; Cao, Z.-J.; Mo, J.-Z.; Li, Y.; Fang, J.-Y.; et al. Vagal afferents mediate antinociception of estrogen in a rat model of visceral pain: The involvement of intestinal mucosal mast cells and 5-hydroxytryptamine 3 signaling. J. Pain 2014, 15, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.E.; Martin-Hirsch, P.L.; Martin, F.L. Oestrogen receptor splice variants in the pathogenesis of disease. Cancer Lett. 2010, 288, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Gustafsson, J.Å. The different roles of ER subtypes in cancer biology and therapy. Nat. Rev. Cancer 2011, 11, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Oduwole, O.O.; Isomaa, V.V.; Nokelainen, P.A.; Stenback, F.; Vihko, P.T. Down regulation of estrogen-metabolizing 17 beta-hydroxysteroid dehydrogenase type 2 expression correlates inversely with Ki67 proliferation marker in colon-cancer development. Int. J. Cancer 2002, 97, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Woźny, M.; Brzuzan, P.; Zielonka, Ł.; Gajęcki, M. Expression of CYPscc and 3β-HSD mRNA in bitches ovary after long-term exposure to zearalenone. Bull. Vet. Inst. Pulawy 2011, 55, 777–780. [Google Scholar]
- Juengel, J.L.; Heath, D.A.; Quirke, L.D.; McNatty, K.P. Oestrogen receptor α and β, androgen receptor and progesterone receptor mRNA and protein localization within the developing ovary and in small growing follicles of sheep. Reproduction 2006, 131, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Bishop, C.V.; Stormshak, F. Non-genomic actions of progesterone and estrogens in regulating reproductive events in domestic animals. Vet. J. 2008, 176, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.D.; Gong, J.; de Lange, C.F.M. The gastrointestinal microbiota and its role in monogastric nutrition and health with an emphasis on pigs: Current understanding, possible modulations, and new technologies for ecological studies. Can. J. Anim. Sci. 2005, 85, 421–435. [Google Scholar] [CrossRef]
- Culpepper, T.; Mai, V. Evidence for contributions of gut microbiota to colorectal carcinogenesis. Curr. Nutr. Rep. 2013, 2, 10–18. [Google Scholar] [CrossRef]
- Broom, L. Mycotoxins and the intestine. Anim. Nutr. 2015, 1, 262–265. [Google Scholar] [CrossRef]
- Pedersen, K.; Tannock, G.W. Colonization of the porcine gastrointestinal-tract by lactobacilli. Appl. Environ. Microbiol. 1989, 55, 279–283. [Google Scholar] [PubMed]
- Giang, H.H.; Viet, T.Q.; Ogle, B.; Lindberg, J.E. Growth performance, digestibility, gut environment and health status in weaned piglets fed a diet supplemented with potentially probiotic complexes of lactic acid bacteria. Livest. Sci. 2010, 129, 95–103. [Google Scholar] [CrossRef]
- Rovers, M. Healthy pigs with less use of antibiotics—A nutritional approach in three steps. Int. Pigs Top. 2012, 27, 15–17. [Google Scholar]
- Franco, T.S.; Garcia, S.; Hirooka, E.Y.; Ono, Y.S.; dos Santos, J.S. Lactic acid bacteria in the inhibition of Fusarium graminearum and deoxynivalenol detoxification. J. Appl. Microbiol. 2011, 111, 739–748. [Google Scholar] [CrossRef] [PubMed]
- El-Nezami, H.S.; Chrevatidis, A.; Auriola, S.; Salminen, S.; Mykkanen, H. Removal of common Fusarium toxins in vitro by strains of Lactobacillus and Propionibacterium. Food Addit. Contam. 2002, 19, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Burel, C.; Tanguy, M.; Guerre, P.; Boilletot, E.; Cariolet, R.; Queguiner, M.; Postollec, G.; Pinton, P.; Salvat, G.; Oswald, I.P.; et al. Effect of low dose of fumonisins on pig health: Immune status, intestinal microbiota and sensitivity to Salmonella. Toxins 2013, 5, 841–864. [Google Scholar] [CrossRef] [PubMed]
- Drew, M.D.; van Kessel, A.G.; Estrada, A.E.; Ekpe, E.D.; Zijlstra, R.T. Effect of dietary cereal on intestinal bacterial populations in weaned pigs. Can. J. Anim. Sci. 2002, 82, 607–609. [Google Scholar] [CrossRef]
- Hughes, R.; Rowland, I.R. Metabolic activities of the gut microflora in relation to cancer. Microb. Ecol. Health Dis. 2000, 12, 179–185. [Google Scholar] [CrossRef]
- De Moreno de LeBlanc, A.; Perdigón, G. Reduction of β-glukuronidase and reductase activity by yoghurt in a murine colon cancer model. Biocell 2005, 29, 15–24. [Google Scholar] [PubMed]
- Macfarlane, G.T.; Macfarlane, S. Models for intestinal fermentation: Association between food components, delivery systems, bioavailability and functional interactions in the gut. Curr. Opin. Biotechnol. 2007, 18, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Zinedine, A.; Soriano, J.M.; Molto, J.C.; Manes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kuciel-Lisieska, G.; Obremski, K.; Stelmachów, J.; Gajęcka, M.; Zielonka, Ł.; Jakimiuk, E.; Gajęcki, M. Presence of zearalenone in blood plasma in women with neoplastic lesions in the mammary gland. Bull. Vet. Inst. Pulawy 2008, 52, 671–674. [Google Scholar]
- Klinder, A.; Karlsson, P.C.; Clune, Y.; Hughes, R.; Glei, M.; Rafter, J.; Rowland, I.; Collins, J.K.; Pool-Zobel, B.L. Faecal water as a non-invasive biomarker in nutritional intervention: Comparison of preparation methods and refinement of different endpoints. Nutr. Cancer 2007, 57, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.R.; Gill, C.I.R.; Rowland, I.R. Diet, faecal water, and colon cancer—Development of a biomarker. Nutr. Rev. 2009, 6, 509–526. [Google Scholar] [CrossRef] [PubMed]
- De Ruyck, K.; de Bovre, M.; Huybrechts, I.; de Saeger, S. Dietary mycotoxins, co-exposure, and carcinogenesis in humans: Short review. Mutat. Res. Rev. Mutat. Res. 2015, 766, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Van der Aar, P.J.; Molist, F.; van der Klis, J.D. The central role of intestinal health on the effectof feed additives on feed intake in swine and poultry. Anim. Feed 2016. [Google Scholar] [CrossRef]
- Antonissen, G.; Martel, A.; Pasmans, F.; Ducatelle, R.; Verbrugghe, E.; Vandenbroucke, V.; Li, S.; Haesebrouck, F.; Immerseel, F.V.; Croubels, S. The impact of Fusarium mycotoxins on human and animal host susceptibility to infectious diseases. Toxins 2014, 6, 430–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grześk, E.; Grześk, G.; Koziński, M.; Stolarek, W.; Zieliński, M.; Kubica, J. Nitric oxide as a cause and a potential place therapeutic intervention in hyporesponsiveness vascular in early sepsis. Folia Cardiol. 2011, 6, 36–43. [Google Scholar]
- Bennett, M.R. Non-adrenergic non-cholinergic (NANC) transmission to smooth muscle: 35 years on. Prog. Neurobiol. 1997, 52, 159–195. [Google Scholar] [CrossRef]
- Carlson, S.J.; Chang, M.I.; Nandivada, P.; Cowan, E.; Puder, M. Neonatal intestinal physiology and failure. Semin. Pediatr. Surg. 2013, 22, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, G.; van Goor, H.; Jansen, P.L.; Moshage, H. Targeting nitric oxide in the gastrointestinal tract. Curr. Opin. Investig. Drugs 2004, 5, 529–536. [Google Scholar] [PubMed]
- Davila, A.-M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.-H.; Sanz, Y.; Tomé, D. Re-print of “Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host”. Pharmacol. Res. 2013, 69, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.J.; Yim, E.Y.; Song, G.; Kim, G.O.; Hyun, C.G. Inhibition of nitric oxide production in lipopolysaccharide-activated RAW 264.7 macrophages by Jeju plant extracts. Interdiscip. Toxicol. 2009, 2, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Prodanov-Radulović, J.; Došenz, R.; Stojanov, I.; Polaček, V.; Živkov-Baloš, M.; Marčić, D.; Pušić, I. The interaction between the swine infectious diseases agents and low levels of mycotoxins in swine feed. Biotechnol. Anim. Husb. 2014, 30, 433–444. [Google Scholar] [CrossRef]
- Maresca, M.; Yahi, N.; Younès-Sakr, L.; Boyron, M.; Caporiccio, B.; Fantini, J. Both direct and indirect effects account for the proinflammatory activity of enteropathogenic mycotoxins on the human intestinal epithelium: Stimulation of interleukin-8 secretion, potentiation of interleukin-1beta effect and increase in the transepithelial passage of commensal bacteria. Toxicol. Appl. Pharmacol. 2008, 228, 84–92. [Google Scholar] [PubMed]
- Levkut, M.; Revajova, V.; Slaminkova, Z.; Levkutova, M.; Borutova, R.; Gresakova, L.; Leng, L. Lymphocyte subpopulations in blood and duodenal epithelium of broilers fed diets contaminated with deoxynivalenol and zearalenone. Anim. Feed Sci. Technol. 2011, 165, 210–217. [Google Scholar] [CrossRef]
- Prims, S.; Tambuyzer, B.; Vergauwen, H.; Huygelen, V.; van Cruchten, S.; van Ginneken, C.; Casteleyn, C. Intestinal immune cell quantification and gram type classification of the adherent microbiota in conventionally and artificially reared, normal and low birth weight piglets. Livest. Sci. 2016, 185, 1–7. [Google Scholar] [CrossRef]
- Gajęcka, M.; Tarasiuk, M.; Zielonka, Ł.; Dąbrowski, M.; Gajęcki, M. Risk assessment for changes in metabolic profile and body weight of pre-pubertal gilts during long-term monotonic exposure to low doses of zearalenone (ZEN). Res. Vet. Sci. 2016, 109, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Jilani, K.; Lang, F. Ca2+-dependent suicidal erythrocyte death following zearalenone exposure. Arch. Toxicol. 2013, 87, 1821–1828. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.Z.; Yang, Z.B.; Yang, W.R.; Gao, J.; Liu, F.X.; Broomhead, J.; Chi, F. Effects of purified zearalenone on growth performance, organ size, serum metabolites, and oxidative stress in postweaning gilts. J. Anim. Sci. 2011, 89, 3008–3015. [Google Scholar] [CrossRef] [PubMed]
- Ingawale, D.K.; Mandlikb, S.K.; Naik, S.R. Models of hepatotoxicity and the underlying cellular, biochemical and immunological mechanism(s): A critical discussion. Environ. Toxicol. Pharmacol. 2014, 37, 118–133. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Rybarczyk, L.; Jakimiuk, E.; Zielonka, Ł.; Obremski, K.; Zwierzchowski, W.; Gajęcki, M. The effect of experimental long-term exposure to low-dose zearalenone on uterine histology in sexually immature gilts. Exp. Toxicol. Pathol. 2012, 64, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, F.M.; de Sousa, F.R.; Barbosa, A.L.; Martins, S.C.; Araújo, R.L.; Soares, R.; Abreu, C. Metabolic syndrome and risk of cancer: Which link? Metabolism 2015, 64, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.Z.; Yang, Z.B.; Yang, W.R.; Wang, S.J.; Liu, F.X.; Johnston, L.A.; Chi, F.; Wang, Y. Effect of purified zearalenone with or without modified montmorillonite on nutrient availability, genital organs and serum hormones in post-weaning piglets. Livest. Sci. 2012, 144, 110–118. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, Y.; Gu, J.; Yuan, Y.; Liu, X.; Zheng, W.; Huang, Q.; Liu, Z.; Bian, J. Zearalenone inhibits testosterone biosynthesis in mouse Leydig cells via the crosstalk of estrogen receptor signaling and orphan nuclear receptor Nur77 expression. Toxicol. In Vitro 2014, 28, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Tiemann, U.; Brüssow, K.-P.; Küchenmeister, U.; Jonas, L.; Kohlschein, P.; Pöhland, R.; Dänicke, S. Influence of diets with cereal grains contaminated by graded levels of two Fusarium toxins on selected enzymatic and histological parameters of liver in gilts. Food Chem. Toxicol. 2006, 44, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.B.; Meca, G.; Font, G.; Ferrer, E. Risk assessment of beauvericin, enniatins and fusaproliferin present in follow-up infant formula by in vitro evaluation of the duodenal and colonic bioaccessibility. Food Control 2014, 42, 234–241. [Google Scholar] [CrossRef]
- Wang, J.; Ji, H.F.; Hou, C.L.; Wang, S.X.; Zhang, D.Y.; Liu, H.; Shan, D.C.; Wang, Y.M. Effects of Lactobacillus johnsonii XS4 supplementation on reproductive performance, gut environment, and blood biochemical and immunological index in lactating sows. Livest. Sci. 2014, 164, 96–101. [Google Scholar] [CrossRef]
- De Angelis, E.; Monaci, L.; Visconti, A. Investigation on the stability of deoxynivalenol and DON-3 glucoside during gastro-duodenal in vitro digestion of a naturally contaminated bread model food. Food Control 2014, 43, 270–275. [Google Scholar] [CrossRef]
- Etzel, R.A. What the primary care pediatrician should know about syndromes associated with exposures to mycotoxins. Curr. Probl. Pediatr. Adolesc. Health Care 2006, 36, 282–305. [Google Scholar] [CrossRef] [PubMed]
- Fleck, S.C.; Hildebrand, A.A.; Müller, E.; Pfeiffer, E.; Metzler, M. Genotoxicity and inactivation of catechol metabolites of the mycotoxin zearalenone. Mycotoxin Res. 2012, 28, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Büning, C.; Geissler, N.; Prager, M.; Sturm, A.; Baumgart, D.C.; Büttner, J.; Bühner, S.; Haas, V.; Lochs, H. Increased small intestinal permeability in ulcerative colitis: Rather genetic than environmental and a risk factor for extensive disease? Inflamm. Bowel Dis. 2012, 18, 1932–1939. [Google Scholar] [CrossRef] [PubMed]
- Gerez, J.R.; Pinton, P.; Callu, P.; Grosjean, F.; Oswald, I.P.; Bracarense, A.P.F.L. Deoxynivalenol alone or in combination with nivalenol and zearalenone induce systemic histological changes in pigs. Exp. Toxicol. Pathol. 2015, 67, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Speranda, M.; Liker, B.; Speranda, T.; Seric, V.; Antunovic, Z.; Grabarevic, Z.; Sencic, D.; Grguric, D.; Steiner, Z. Haematological and biochemical parameters of weaned piglets fed on fodder mixture contaminated by zearalenone with addition of clinoptilolite. Acta Vet. Beograd. 2006, 56, 121–136. [Google Scholar]
- Jakimiuk, E.; Kuciel-Lisiecka, G.; Zwierzchowski, W.; Gajęcka, M.; Obremski, K.; Zielonka, Ł.; Skorska-Wyszyńska, E.; Gajęcki, M. Morphometric changes of the reproductive system in gilts during zearalenone mycotoxicosis. Med. Weter. 2006, 62, 99–102. [Google Scholar]
- De Boevre, M.; Graniczkowska, K.; de Saeger, S. Metabolism of modified mycotoxins studied through in vitro and in vivo models: An overview. Toxicol. Lett. 2015, 233, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Parr, T.; Mareko, M.H.D.; Ryan, K.J.P.; Hemmings, K.M.; Brown, D.M.; Brameld, J.M. The impact of growth promoters on muscle growth and the potential consequences for meat quality. Meat Sci. 2016, 120, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Zhabinskii, V.N.; Khripach, N.B.; Khripach, V.A. Steroid plant hormones: Effects outside plant kingdom. Steroids 2015, 97, 87–97. [Google Scholar] [CrossRef] [PubMed]




© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. Activity of Zearalenone in the Porcine Intestinal Tract. Molecules 2017, 22, 18. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22010018
Gajęcka M, Zielonka Ł, Gajęcki M. Activity of Zearalenone in the Porcine Intestinal Tract. Molecules. 2017; 22(1):18. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22010018
Chicago/Turabian StyleGajęcka, Magdalena, Łukasz Zielonka, and Maciej Gajęcki. 2017. "Activity of Zearalenone in the Porcine Intestinal Tract" Molecules 22, no. 1: 18. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22010018