
Profile of Bioactive Compounds in the Morphological Parts of Wild Fallopia japonica (Houtt) and Fallopia sachalinensis (F. Schmidt) and Their Antioxidative Activity



Abstract
:1. Introduction
2. Results and Discussion
2.1. Content of Phenolic Compounds
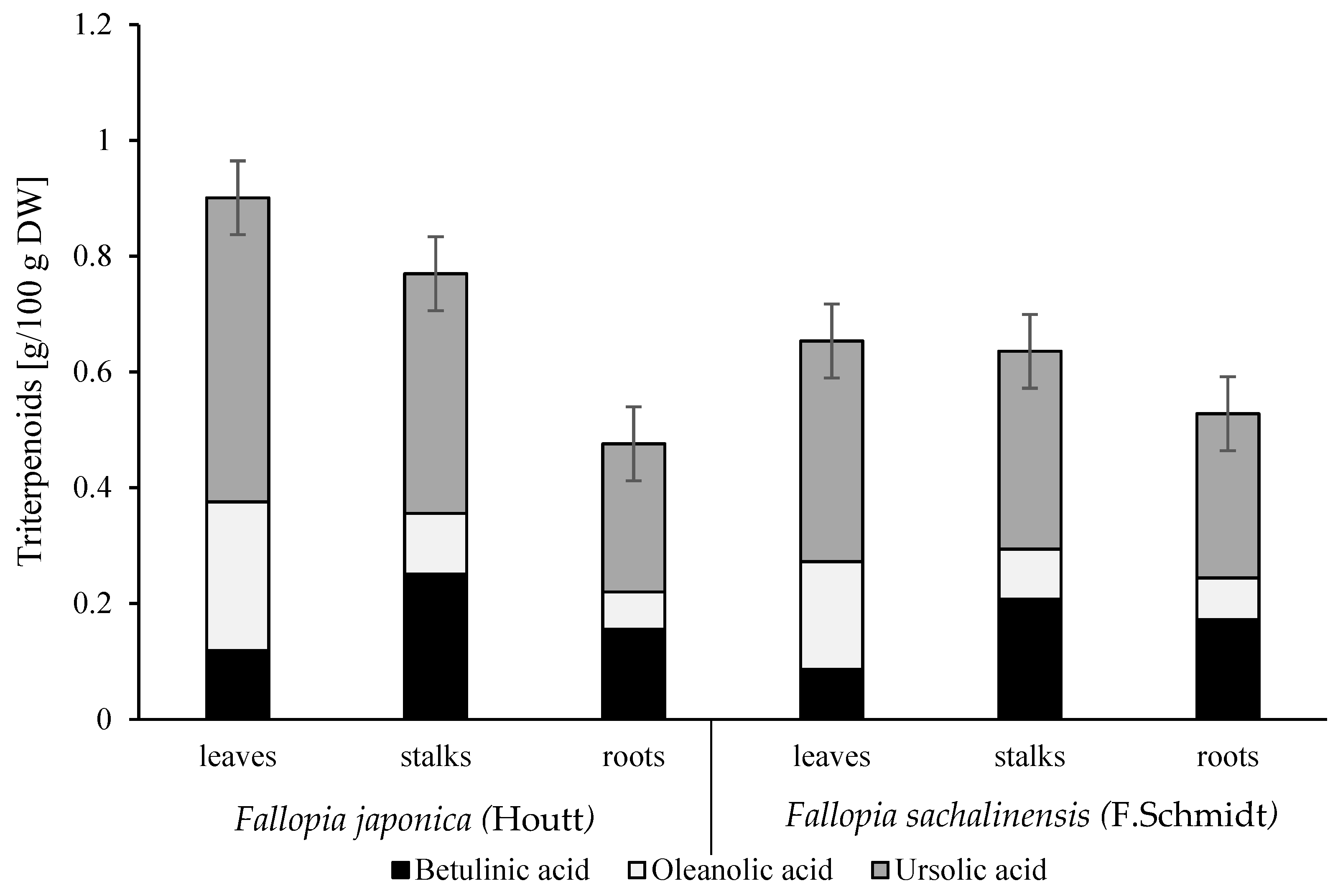
2.2. Determination of Tritepenoids
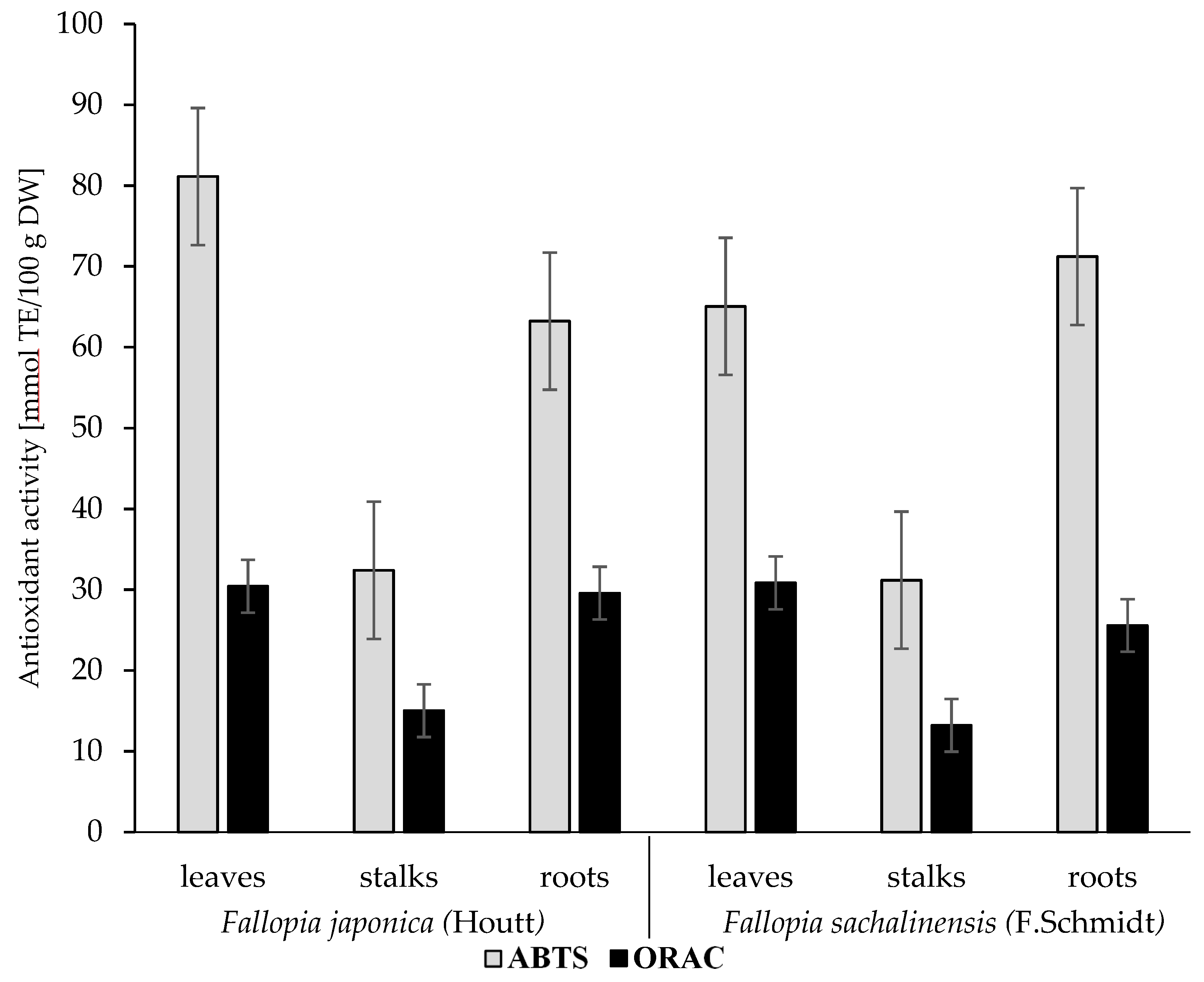
2.3. Antioxidant Activity
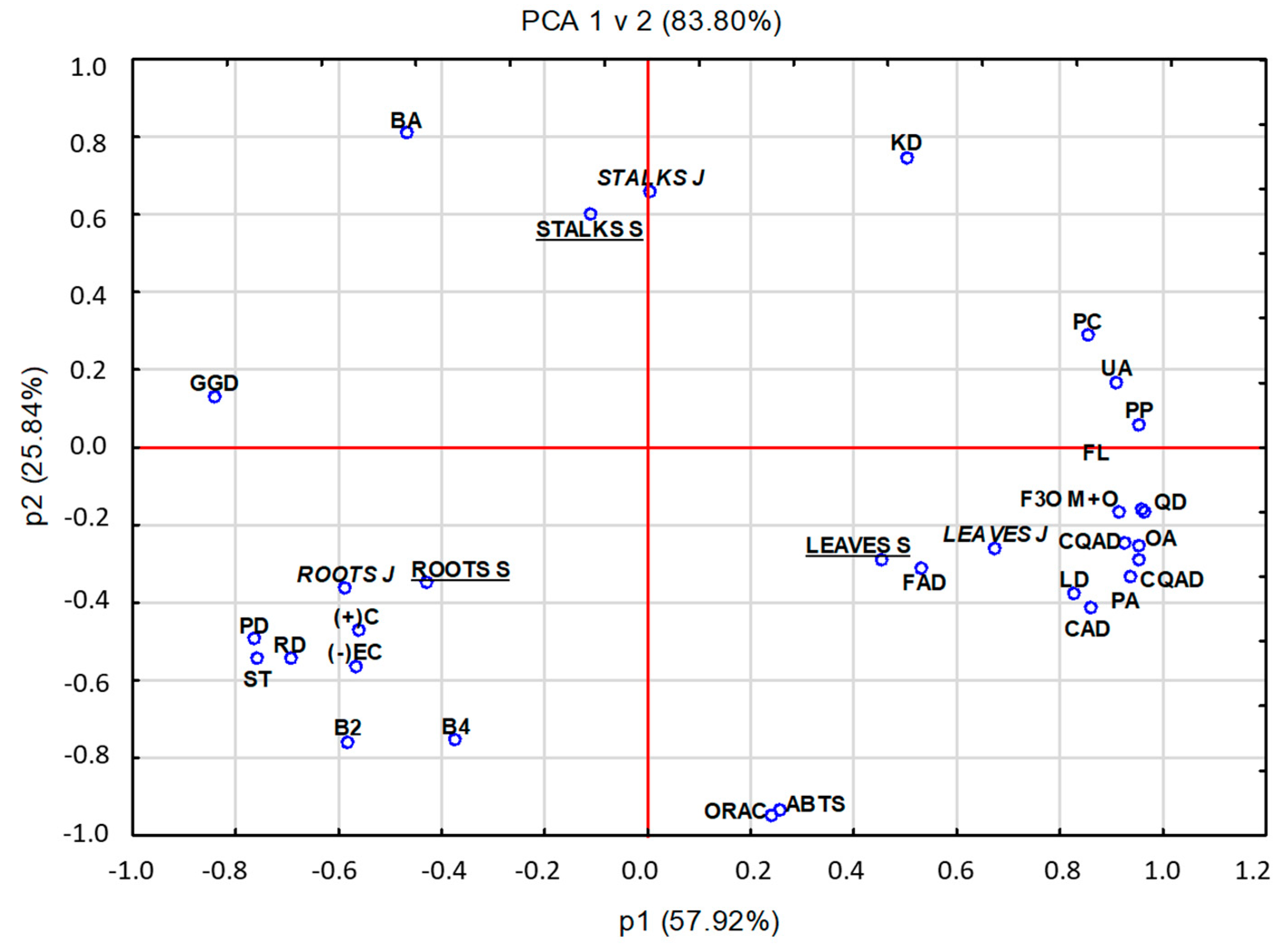
2.4. Principal Component Analysis (PCA)
3. Materials and Methods
3.1. Reagent and Standards
3.2. Plant Materials
3.3. Determination of Polyphenols
3.4. Determination of Proanthocyanidins
3.5. Determination of Triterpenoids
3.6. Determination of Antioxidative Activity
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jeong, E.T.; Jin, M.H.; Kim, M.-S.; Chang, Y.H.; Park, S.G. Inhibition of melanogenesis by piceid isolated from Polygonum cuspidatum. Arch. Pharm. Res. 2010, 33, 1331–1338. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Marston, A.; Hay, A.-E.; Hostettmann, K. Rapid separation of three glucosylated resveratrol analogues from the invasive plant Polygonum cuspidatum by high-speed countercurrent chromatography. J. Sep. Sci. 2009, 32, 2979–2984. [Google Scholar] [CrossRef]
- Kim, H.; Song, M.J. Ethnobotanical analysis for traditional knowledge of wild edible plants in North Jeolla Province (Korea). Genet Resour. Crop Evol. 2013, 60, 1571–1585. [Google Scholar] [CrossRef]
- Bralley, E.E.; Greenspan, P.; Hargrove, J.L.; Wicker, L.; Hartle, D.K. Topical anti-inflammatory activity of Polygonum cuspidatum extract in the TPA model of mouse ear inflammation. J. Inflamm. 2008, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-Y.; Cai, Y.-Z.; Xing, J.; Corke, H.; Sun, M. Comparative Analysis of Bioactivities of Four Polygonum Species. Planta Med. 2008, 74, 43–49. [Google Scholar] [CrossRef]
- Shukla, Y.; Singh, R. Resveratrol and cellular mechanisms of cancer prevention. Ann. N. Y. Acad. Sci. 2011, 1215, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahedi, H.S.; Jazayeri, S.; Ghiasvand, R.; Djalali, M.; Eshraghian, M.R. Effects of Polygonum Cuspidatum Containing Resveratrol on Inflammation in Male Professional Basketball Players. Int. J. Prev. Med. 2013, 4, S1–S4. [Google Scholar]
- Goc, A.; Niedzwiecki, A.; Rath, M. In vitro evaluation of antibacterial activity of phytochemicals and micronutrients against Borrelia burgdorferi and Borrelia garinii. J. Appl. Microbiol. 2015, 119, 1561–1572. [Google Scholar] [CrossRef]
- Cassidy, A.; Hanley, B.; Lamuela-Raventos, R.M. Isoflavones, lignans and stilbenes—origins, metabolism and potential importance to human health. J. Sci. Food Agric. 2000, 80, 1044–1062. [Google Scholar] [CrossRef]
- Ares, A.M.; Soto, M.E.; Nozal, M.J.; Bernal, J.L.; Higes, M.; Bernal, J. Determination of Resveratrol and Piceid Isomers in Bee Pollen by Liquid Chromatography Coupled to Electrospray Ionization-Mass Spectrometry. Food Anal. Methods 2015, 8, 1565–1575. [Google Scholar] [CrossRef]
- Wang, T.-H.; Zhang, J.; Qiu, X.-H.; Bai, J.-Q.; Gao, Y.-H.; Xu, W. Application of Ultra-High-Performance Liquid Chromatography Coupled with LTQ-Orbitrap Mass Spectrometry for the Qualitative and Quantitative Analysis of Polygonum multiflorum Thumb. and Its Processed Products. Molecules 2016, 21, 40. [Google Scholar] [CrossRef]
- Lin, L.; Lin, H.; Zhang, M.; Ni, B.; Yin, X.; Qu, C.; Ni, J. A novel method to analyze hepatotoxic components in Polygonum multiflorum using ultra-performance liquid chromatography-quadrupole time-of-flight mass spectrometry. J. Hazard. Mater. 2015, 299, 249–259. [Google Scholar] [CrossRef]
- Qiu, X.; Zhang, J.; Huang, Z.; Zhu, D.; Xu, W. Profiling of phenolic constituents in Polygonum multiflorum Thunb. by combination of ultra-high-pressure liquid chromatography with linear ion trap-Orbitrap mass spectrometry. J. Chromatogr. A 2013, 1292, 121–131. [Google Scholar] [CrossRef]
- Beňová, B.; Adam, M.; Pavlíková, P.; Fischer, J. Supercritical fluid extraction of piceid, resveratrol and emodin from Japanese knotweed. J. Supercrit. Fluids 2010, 51, 325–330. [Google Scholar] [CrossRef]
- Lachowicz, S.; Oszmiański, J.; Wojdyło, A.; Cebulak, T.; Hirnle, L.; Siewiński, M. UPLC-PDA-Q/TOF-MS identification of bioactive compounds and on-line UPLC-ABTS assay in Fallopia japonica Houtt and Fallopia sachalinensis (F. Schmidt) leaves and rhizomes grown in Poland. Eur. Food Res. Technol. 2019, 245, 691–706. [Google Scholar] [CrossRef]
- Loza-Mejía, M.A.; Salazar, J.R. Sterols and triterpenoids as potential anti-inflammatories: Molecular docking studies for binding to some enzymes involved in inflammatory pathways. J. Mol. Graph. Model. 2015, 62, 18–25. [Google Scholar] [CrossRef]
- Lachowicz, S.; Kolniak-Ostek, J.; Oszmiański, J.; Wiśniewski, R. Comparison of Phenolic Content and Antioxidant Capacity of Bear Garlic (Allium ursinum L.) in Different Maturity Stages. J. Food Process. Preserv. 2017, 41, e12921. [Google Scholar] [CrossRef]
- Lachowicz, S.; Oszmiański, J.; Wiśniewski, R. Determination of triterpenoids, carotenoids, chlorophylls, and antioxidant capacity in Allium ursinum L. at different times of harvesting and morphological parts. Eur. Food Res. Technol. 2018, 244, 1269–1280. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Czemerys, R. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem. 2007, 105, 940–949. [Google Scholar] [CrossRef]
- Katalinic, V.; Milos, M.; Kulisic, T.; Jukic, M. Screening of 70 medicinal plant extracts for antioxidant capacity and total phenols. Food Chem. 2006, 94, 550–557. [Google Scholar] [CrossRef]
- Pourmorad, F.; Hosseinimehr, S.J.; Shahabimajd, N. Antioxidant activity, phenol and flavonoid contents of some selected Iranian medicinal plants. Afr. J. Biotechnol. 2006, 5. [Google Scholar]
- Ivanova, D.; Gerova, D.; Chervenkov, T.; Yankova, T. Polyphenols and antioxidant capacity of Bulgarian medicinal plants. J. Ethnopharmacol. 2005, 96, 145–150. [Google Scholar] [CrossRef]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant activity of some algerian medicinal plants extracts containing phenolic compounds. Food Chem. 2006, 97, 654–660. [Google Scholar] [CrossRef]
- Chen, H.F.; Chen, Y.H.; Liu, C.H.; Wang, L.; Chen, X.; Yu, B.Y.; Qi, J. Integrated chemometric fingerprints of antioxidant activities and HPLC–DAD–CL for assessing the quality of the processed roots of Polygonum multiflorum Thunb. (Heshouwu). Chin. Med. 2016, 11, 18. [Google Scholar] [CrossRef]
- Lavola, A.; Karjalainen, R.; Julkunen-Tiitto, R. Bioactive polyphenols in leaves, stems, and berries of Saskatoon (Amelanchier alnifolia Nutt.) cultivars. J. Agric. Food Chem. 2012, 60. [Google Scholar] [CrossRef]
- Park, Y.-S.; Namiesnik, J.; Vearasilp, K.; Leontowicz, H.; Leontowicz, M.; Barasch, D.; Nemirovski, A.; Trakhtenberg, S.; Gorinstein, S. Bioactive compounds and the antioxidant capacity in new kiwi fruit cultivars. Food Chem. 2014, 165, 354–361. [Google Scholar] [CrossRef]
- Romero-Pérez, A.I.; Ibern-Gómez, M.; Lamuela-Raventós, R.M.; de la Torre-Boronat, M.C. Piceid, the Major Resveratrol Derivative in Grape Juices. J. Agric. Food Chem. 1999, 47, 1533–1536. [Google Scholar] [CrossRef]
- Cacho, J.I.; Campillo, N.; Viñas, P.; Hernández-Córdoba, M. Stir bar sorptive extraction with gas chromatography–mass spectrometry for the determination of resveratrol, piceatannol and oxyresveratrol isomers in wines. J. Chromatogr. A 2013, 1315, 21–27. [Google Scholar] [CrossRef]
- Szakiel, A.; Pączkowski, C.; Pensec, F.; Bertsch, C. Fruit cuticular waxes as a source of biologically active triterpenoids. Phytochem. Rev. 2012, 11, 263–284. [Google Scholar] [CrossRef] [Green Version]
- Sproull, G.J.; Quigley, M.F.; Sher, A.; González, E. Long-term changes in composition, diversity and distribution patterns in four herbaceous plant communities along an elevational gradient. J. Veget. Sci. 2015, 26, 552–563. [Google Scholar] [CrossRef]
- Farneti, B.; Masuero, D.; Costa, F.; Magnago, P.; Malnoy, M.; Costa, G.; Vrhovsek, U.; Mattivi, F. Is there room for improving the nutraceutical composition of apple? J. Agric. Food Chem. 2015, 63, 2750–2759. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Kapusta, I.; Cebulak, T.; Oszmiański, J. Characterization of polish wines produced from the interspecific hybrid grapes grown in south-east Poland. Eur. Food Res. Technol. 2018, 244, 441–455. [Google Scholar] [CrossRef]
Sample Availability: Samples of the phytochemical composition and antioxidative activity of plants of the wild Fallopia species and their morphological parts. |




{kind=link}
{kind=link}
{kind=link}
| Tentative Identification | Retention Time [min] | λ (nm) | Molecular ion MS [H − M]− | Fragments MS/MS (m/z) |
|---|---|---|---|---|
| Galloyl glucose | 1.40 | 277 | 331 | 169 |
| Galloyl glucose | 1.45 | 277 | 331 | 169 |
| Galloyl glucose | 1.57 | 277 | 331 | 169 |
| 3-O-caffeoylquinic acid | 3.25 | 324 | 353 | 191/179 |
| Cis 3-O-caffeoylquinic acid | 3.47 | 326 | 353 | 191 |
| Caftaric acid | 3.58 | 328 | 311 | 179 |
| Procyanidin dimer B | 4.06 | 279 | 577 | 289 |
| Caffeoyl-glucose | 4.16 | 341 | 179 | |
| p-Coumaroylquinic acid | 4.26 | 310 | 337 | 191/163 |
| (+) Catechin | 4.50 | 279 | 289 | |
| 5-O-caffeoylquinic acid | 4.62 | 324 | 353 | 191 |
| Cis 5-O-caffeoylquinic acid | 4.74 | 324 | 353 | 191 |
| Feruloylquinic acid | 5.09 | 320 | 367 | 191 |
| Procyanidin dimer B | 5.19 | 279 | 577 | 289 |
| Procyanidin dimer B | 5.56 | 279 | 577 | 289 |
| p-Coumaroylquinic acid | 5.68 | 310 | 337 | 191/163 |
| Resveratroloside | 5.90 | 303 | 389 | 227 |
| (−)-Epicatechin | 6.01 | 278 | 289 | |
| Astringin | 6.03 | 328 | 405 | 243 |
| Piceatannol 3′-O-glucoside | 6.14 | 305 | 405 | 243 |
| (+)-Catechin glucoside | 6.30 | 277 | 451 | 289 |
| Trans-Piceid | 6.35 | 317 | 389 | 227 |
| Procyanidin B gallate | 6.61 | 279 | 729 | 577/289 |
| Procyanidin tetramer B | 6.72 | 277 | 1153 | 863/575/289 |
| Procyanidin tetramer B | 6.88 | 279 | 1153 | 863/575/289 |
| Luteolin 7-O-galactoside | 7.00 | 349 | 447 | 285 |
| Luteolin 7-O-glucoside | 7.15 | 347 | 447 | 285 |
| Resveratrol-galleoyl-glucoside | 7.22 | 333 | 541 | 227 |
| (+)Catechin gallate | 7.31 | 277 | 441 | 289 |
| Quercetin-3-O-rhamno-glucoside | 7.32 | 352 | 609 | 431/301 |
| Quercetin 3-O-rutinoside | 7.40 | 352 | 609 | 431/301 |
| Procyanidin tetramer B | 7.45 | 277 | 1153 | 863/575/289 |
| Quercetin 3-O-galatcoside | 7.56 | 352 | 463 | 301 |
| Quercetin 3-O-glucoside | 7.67 | 353 | 463 | 301 |
| Quercetin 3-O-pentoside | 7.99 | 355 | 433 | 301 |
| cis-Piceid | 8.10 | 285 | 389 | 227 |
| Quercetin acetylhexoside | 8.20 | 354 | 505 | 463/301 |
| Quercetin 3-O-pentoside | 8.35 | 351 | 433 | 301 |
| Luteolin 7-O-rhamnoside | 8.40 | 342 | 431 | 285 |
| Quercetin-3-O-rhamnoside | 8.51 | 346 | 447 | 301 |
| Kaempferol -3-O-galactoside | 9.18 | 345 | 447 | 285 |
| Kaempferol -3-O-glucoside | 9.25 | 345 | 447 | 285 |
| 3,4-Di-O-caffeoylquinic acid | 9.26 | 324 | 515 | 353/191 |
| 3,5-Di-O-caffeoylquinic acid | 9.37 | 326 | 515 | 353/191 |
| 4,5-Di-O-caffeoylquinic acid | 9.49 | 326 | 515 | 353/191 |
| trans-Resveratrol | 9.51 | 285 | 227 | |
| Kaempferol 3-O-rhamnoside | 9.57 | 340 | 431 | 285 |
| Quercetin | 10.84 | 364 | 301 | |
| Betulinic acid | 6.99 | 455 | ||
| Oleanolic acid | 7.66 | 455 | ||
| Ursolic acid | 8.38 | 455 |
| Group | Compounds | Fallopia japonica (Houtt) | Fallopia sachalinensis (F. Schmidt) | ||||
|---|---|---|---|---|---|---|---|
| leaves | stalks | roots | leaves | stalks | roots | ||
| Flavan-3-ols | Procyanidin dimer B 3 | 0.09 ± 0.01c 1,2 | 0.02 ± 0.00f | 0.06 ± 0.00e | 0.13 ± 0.00b | 0.06 ± 0.00d | 0.17 ± 0.00a |
| (+) Catechin 3 | 0.19 ± 0.01b | 0.08 ± 0.01f | 0.09 ± 0.00e | 0.14 ± 0.01c | 0.09 ± 0.00d | 0.22 ± 0.00a | |
| Procyanidin dimer B 3 | 0.17 ± 0.01e | 0.08 ± 0.01f | 0.28 ± 0.01b | 0.23 ± 0.01d | 0.24 ± 0.00c | 0.37 ± 0.01a | |
| Procyanidin dimer B 3 | 0.26 ± 0.01b | 0.18 ± 0.01d | 0.63 ± 0.02a | 0.09 ± 0.01d | 0.07 ± 0.01f | 0.19 ± 0.00c | |
| (-)Epicatechin 3 | 0.13 ± 0.01d | 0.06 ± 0.00f | 0.27 ± 0.01b | 0.13 ± 0.01c | 0.11 ± 0.00e | 0.62 ± 0.01a | |
| (+)Catechin glucoside 4 | 0.13 ± 0.00b | 0.11 ± 0.01d | 1.48 ± 0.00a | 0.062 ± 0.00f | 0.06 ± 0.00d | 0.09 ± 0.00c | |
| Procyanidin B gallate 4 | 0.07 ± 0.00d | 0.04 ± 0.00e | 0.12 ± 0.01a | 0.09 ± 0.00c | 0.04 ± 0.00f | 0.09 ± 0.00b | |
| Procyanidin tetramer B 3 | 0.04 ± 0.00c | 0.01 ± 0.00f | 0.02 ± 0.00e | 0.04 ± 0.00b | 0.03 ± 0.00d | 0.07 ± 0.00a | |
| Procyanidin tetramer B 3 | 0.06 ± 0.01d | 0.02 ± 0.00f | 0.09 ± 0.01b | 0.07 ± 0.00c | 0.03 ± 0.00e | 0.17 ± 0.01a | |
| (+)Catechin gallate 4 | 0.09 ± 0.01b | 0.05 ± 0.00e | 0.26 ± 0.01a | 0.06 ± 0.00d | 0.02 ± 0.00f | 0.09 ± 0.00c | |
| Procyanidin tetramer B 3 | 0.07 ± 0.00d | 0.04 ± 0.00f | 0.14 ± 0.00b | 0.13 ± 0.00c | 0.07 ± 0.00e | 0.17 ± 0.01a | |
| Procyanidin polymers | 15143.39 ± 4.54a | 10.05 ± 0.12c | 4.24 ± 0.27f | 10.62 ± 0.19b | 7.97 ± 0.39d | 6.72 ± 0.02e | |
| Phenolic acids | Galloyl glucose 3 | 0.01 ± 0.00d1,2 | 0.010.00fa | 0.01 ± 0.00b | 0.01 ± 0.00e | 0.01 ± 0.00a | 0.01 ± 0.00c |
| Galloyl glucose 3 | 0.01 ± 0.00f | 0.01 ± 0.00d | 0.01 ± 0.00 | 0.01 ± 0.00b | 0.01 ± 0.00c | 0.01 ± 0.00e | |
| Galloyl glucose 3 | 0.01 ± 0.00d | 0.01 ± 0.00a | 0.01 ± 0.00c | 0.01 ± 0.00f | 0.01 ± 0.00e | 0.01 ± 0.00b | |
| 3-O-caffeoylquinic acid 3 | 0.03 ± 0.01b | 0.01 ± 0.00c | 0.00 ± 0.00d | 0.04 ± 0.00a | 0.01 ± 0.00c | 0.01 ± 0.00c | |
| Cis 3-O-caffeoylquinic acid 3 | 0.49 ± 0.01a | 0.07 ± 0.00c | 0.01 ± 0.00f | 0.45 ± 0.01b | 0.07 ± 0.01d | 0.01 ± 0.00e | |
| Caftaric acid 3 | 0.44 ± 0.00b | 0.02 ± 0.00d | 0.01 ± 0.00e | 0.58 ± 0.01a | 0.02 ± 0.00c | 0.00 ± 0.00f | |
| Caffeoyl-glucose 4 | 0.02 ± 0.00a | 0.01 ± 0.00c | 0.01 ± 0.00e | 0.04 ± 0.00b | 0.01 ± 0.00d | 0.01 ± 0.00f | |
| p-Coumaroylquinic acid 3 | 0.23 ± 0.01a | 0.04 ± 0.00c | 0.01 ± 0.00f | 0.13 ± 0.01b | 0.02 ± 0.00d | 0.01 ± 0.00e | |
| 5-O-caffeoylquinic acid 4 | 0.30 ± 0.00a | 0.08 ± 0.00c | 0.01 ± 0.00e | 0.25 ± 0.01b | 0.06 ± 0.01d | 0.01 ± 0.00f | |
| Cis 5-O-caffeoylquinic acid 3 | 0.022 ± 0.00a | 0.01 ± 0.00c | 0.01 ± 0.00e | 0.02 ± 0.00b | 0.01 ± 0.00d | 0.01 ± 0.00f | |
| 3-O-p-Coumaroylquinic acid 3 | 0.01 ± 0.00a | 0.00 ± 0.00e | nd | 0.01 ± 0.00b | 0.01 ± 0.00d | 0.01 ± 0.00c | |
| Feruloylquinic acid 3 | 0.01 ± 0.00c | 0.01 ± 0.00e | nd | 0.02 ± 0.00a | 0.01 ± 0.00d | 0.01 ± 0.00b | |
| 3-O-p-Coumaroylquinic acid 3 | 0.09 ± 0.00a | 0.03 ± 0.01c | nd | 0.03 ± 0.00b | 0.01 ± 0.00d | 0.01 ± 0.00e | |
| 3,4-Di-O-caffeoylquinic acid 4 | nd≠ | nd | nd | nd | nd | 0.01 ± 0.00a | |
| 3,5-Di-O-caffeoylquinic acid 4 | nd | nd | nd | nd | nd | 0.01 ± 0.00a | |
| 4,5-Di-O-caffeoylquinic acid 4 | nd | nd | nd | nd | nd | 0.03 ± 0.00a | |
| Flavones and Flavonols | Luteolin 7-O-galactoside 4 | 0.02 ± 0.00b 1,2 | nd | nd | 0.05 ± 0.00a | 0.01 ± 0.00c | nd |
| Luteolin 7-O-glucoside 3 | 0.01 ± 0.00b | nd | nd | 0.03 ± 0.00a | 0.01 ± 0.00c | nd | |
| Luteolin 7-O-rhamnoside 3 | 0.03 ± 0.00a | nd | nd | 0.02 ± 0.00b | nd | nd | |
| Quercetin-3-O-rhamno-glucoside 4 | 0.02 ± 0.00a | 0.02 ± 0.00c | nd | 0.01 ± 0.00d | 0.02 ± 0.00b | nd | |
| Quercetin 3-O-rutinoside 3 | 0.03 ± 0.00a | 0.02 ± 0.00d | 0.01 ± 0.00e | 0.02 ± 0.00c | 0.02 ± 0.00b | nd | |
| Quercetin 3-O-galactoside 3 | 0.05 ± 0.00a | 0.02 ± 0.00c | 0.01 ± 0.00e | 0.03 ± 0.00b | 0.02 ± 0.00d | 0.01 ± 0.00f | |
| Quercetin glucoside 3 | 0.105 ± 0.01b | 0.07 ± 0.00d | nd | 0.12 ± 0.01a | 0.09 ± 0.00c | nd | |
| Quercetin pentoside 4 | 0.09 ± 0.00a | 0.02 ± 0.00d | 0.01 ± 0.00e | 0.05 ± 0.00b | 0.03 ± 0.00c | nd | |
| Quercetin acetylhexoside 4 | 0.08 ± 0.00a | 0.04 ± 0.00d | 0.01 ± 0.00e | 0.05 ± 0.00c | 0.06 ± 0.00b | nd | |
| Quercetin pentoside 4 | 0.25 ± 0.08a | 0.08 ± 0.00d | 0.01 ± 0.00 | 0.17 ± 0.01b | 0.09 ± 0.00c | nd | |
| Quercetin rhamnoside 4 | 1.71 ± 0.01a | 0.28 ± 0.00c | 0.01 ± 0.00e | 0.89 ± 0.02b | 0.16 ± 0.01d | nd | |
| Kaempferol -3-O-galactoside 3 | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.01 ± 0.00a | nd | |
| Kaempferol -3-O-glucoside 3 | 0.01 ± 0.00a | 0.01 ± 0.00a | nd | 0.01 ± 0.00a | 0.01 ± 0.00a | nd | |
| Kaempferol -3-O-rhamnoside 4 | nd | nd | nd | 0.01 ± 0.00a | 0.01 ± 0.00b | nd | |
| Quercetin 4 | nd | nd | nd | 0.04 ± 0.00a | 0.02 ± 0.00b | nd | |
| Stilbene | Resveratroloside 4 | nd | nd | 0.02 ± 0.00b | nd | nd | 0.03 ± 0.00a |
| Astringin 3 | nd | nd | 0.03 ± 0.00b | nd | nd | 0.05 ± 0.00a | |
| Piceatannol 3′-O-glucoside 4 | 0.02 ± 0.00c 1,2 | 0.01 ± 0.00d | 0.22 ± 0.01a | 0.02 ± 0.00c | 0.01 ± 0.00d | 0.05 ± 0.00b | |
| Trans-Piceid 3 | 0.03 ± 0.00e | 0.04 ± 0.01d | 0.50 ± 0.01a | 0.02 ± 0.00f | 0.06 ± 0.00c | 0.22 ± 0.00b | |
| Resveratrol-galleoyl-glucoside 4 | nd | nd | nd | nd | nd | 0.02 ± 0.00a | |
| Cis-Piceid 3 | 0.01 ± 0.00c | 0.01 ± 0.00c | 0.02 ± 0.00b | 0.01 ± 0.00c | 0.01 ± 0.00b | 0.05 ± 0.00a | |
| Trans-Resveratrol 3 | 0.01 ± 0.00b | 0.01 ± 0.00b | 0.02 ± 0.00a | 0.01 ± 0.00b | 0.01 ± 0.00b | 0.02 ± 0.00a | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lachowicz, S.; Oszmiański, J. Profile of Bioactive Compounds in the Morphological Parts of Wild Fallopia japonica (Houtt) and Fallopia sachalinensis (F. Schmidt) and Their Antioxidative Activity. Molecules 2019, 24, 1436. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24071436
Lachowicz S, Oszmiański J. Profile of Bioactive Compounds in the Morphological Parts of Wild Fallopia japonica (Houtt) and Fallopia sachalinensis (F. Schmidt) and Their Antioxidative Activity. Molecules. 2019; 24(7):1436. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24071436
Chicago/Turabian StyleLachowicz, Sabina, and Jan Oszmiański. 2019. "Profile of Bioactive Compounds in the Morphological Parts of Wild Fallopia japonica (Houtt) and Fallopia sachalinensis (F. Schmidt) and Their Antioxidative Activity" Molecules 24, no. 7: 1436. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24071436