Myrtus Polyphenols, from Antioxidants to Anti-Inflammatory Molecules: Exploring a Network Involving Cytochromes P450 and Vitamin D

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
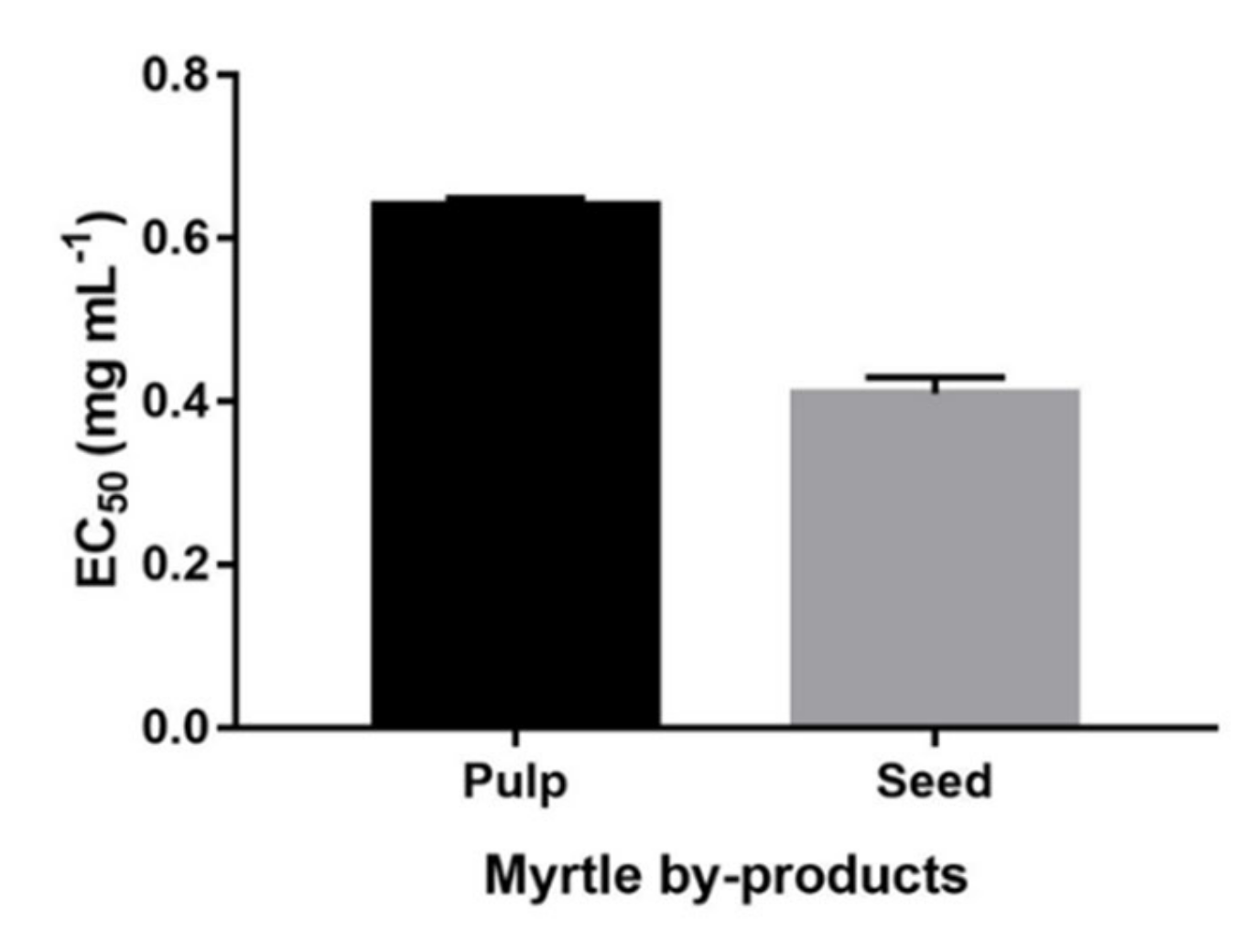
2.1. Quantitative Data Analysis of Main Phenolic Compounds
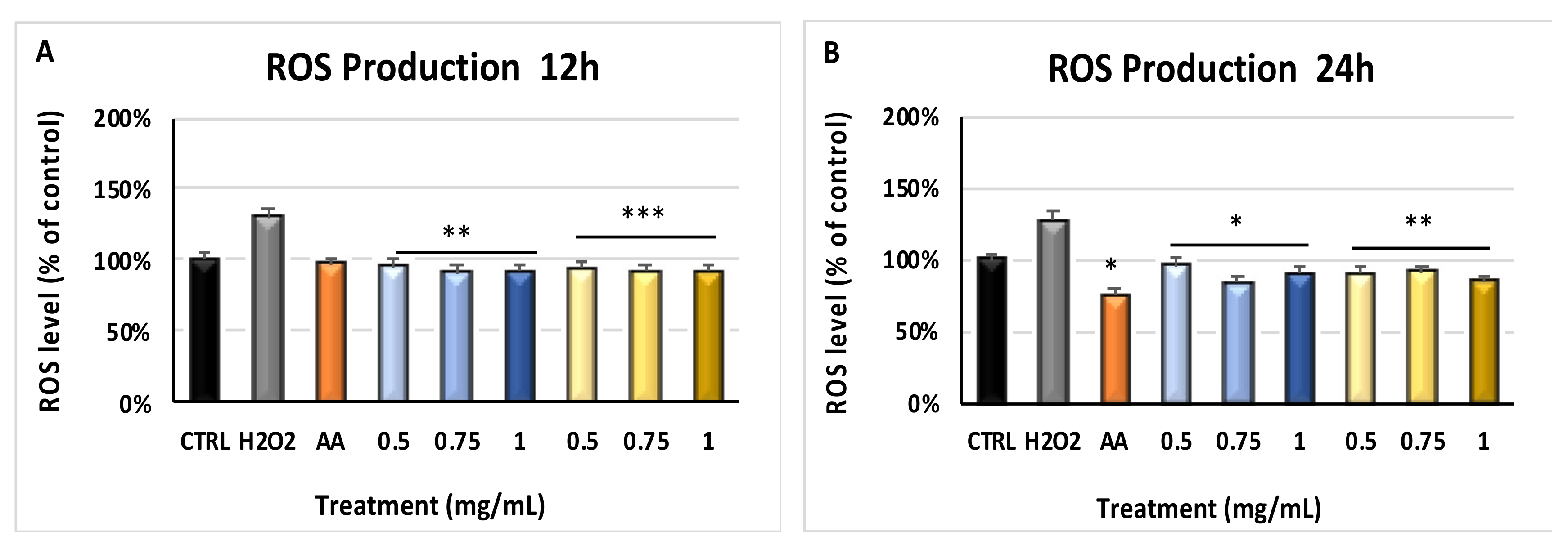
2.2. Myrtus Reduces ROS Production by H2O2
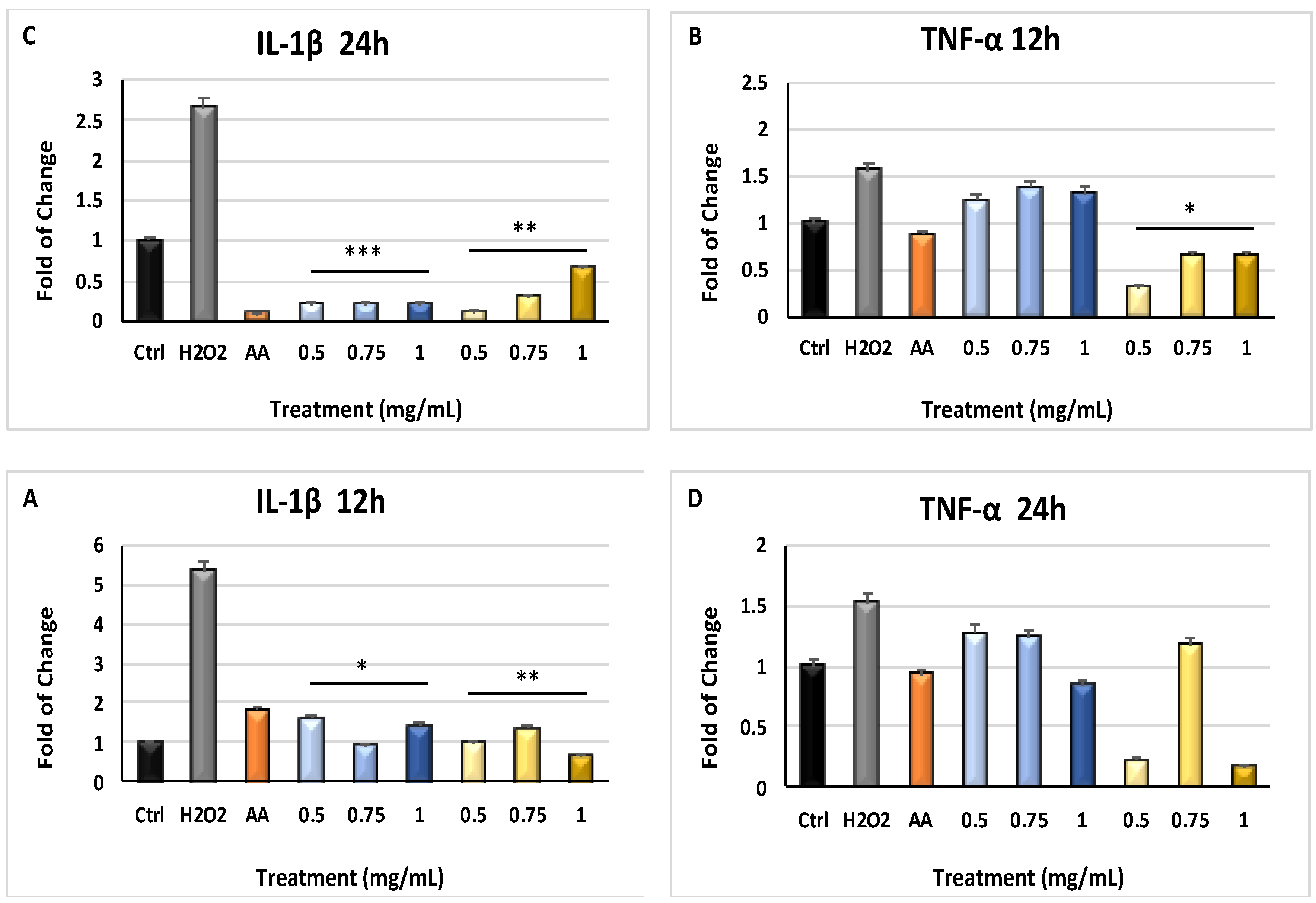
2.3. Inflammatory-Related Gene Modulation by Myrtus Products
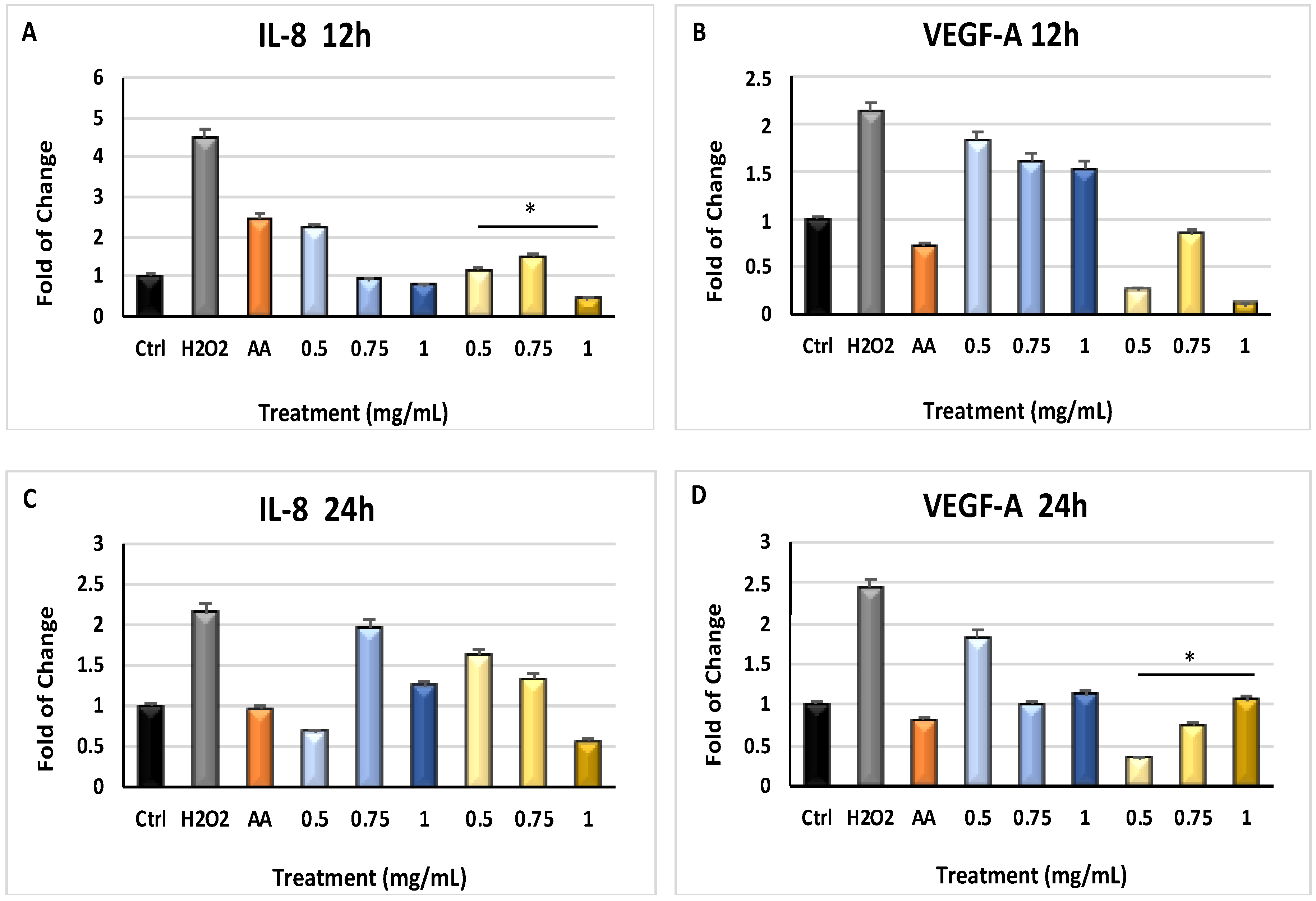
2.4. Myrtus Extracts Act on Angiogenesis by Modulating Inflammatory Mediating Genes
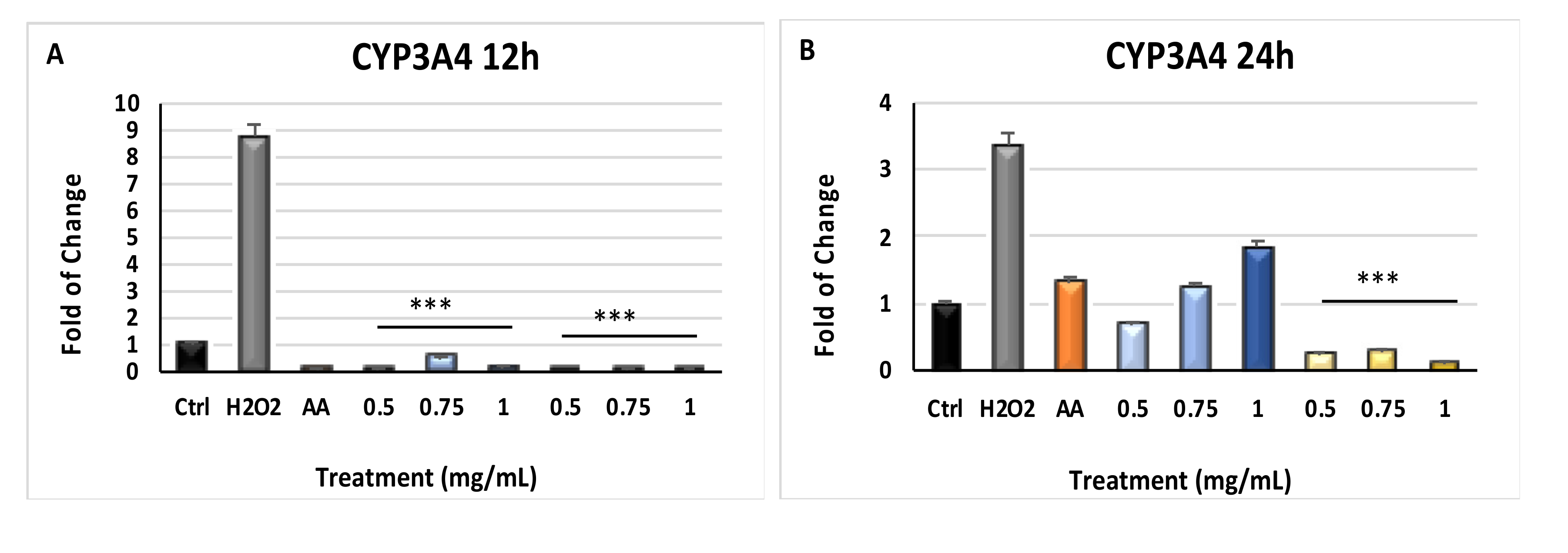
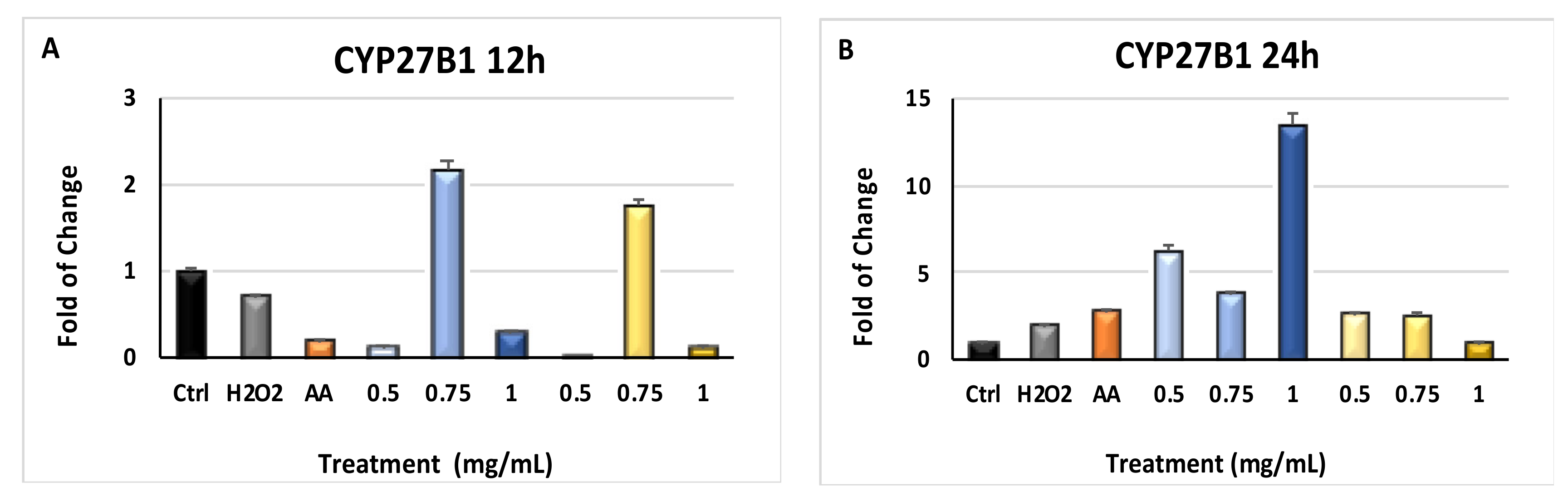
2.5. Modulation of Cytochrome p450 CYP3A4 and CYP27B1 Gene Expression
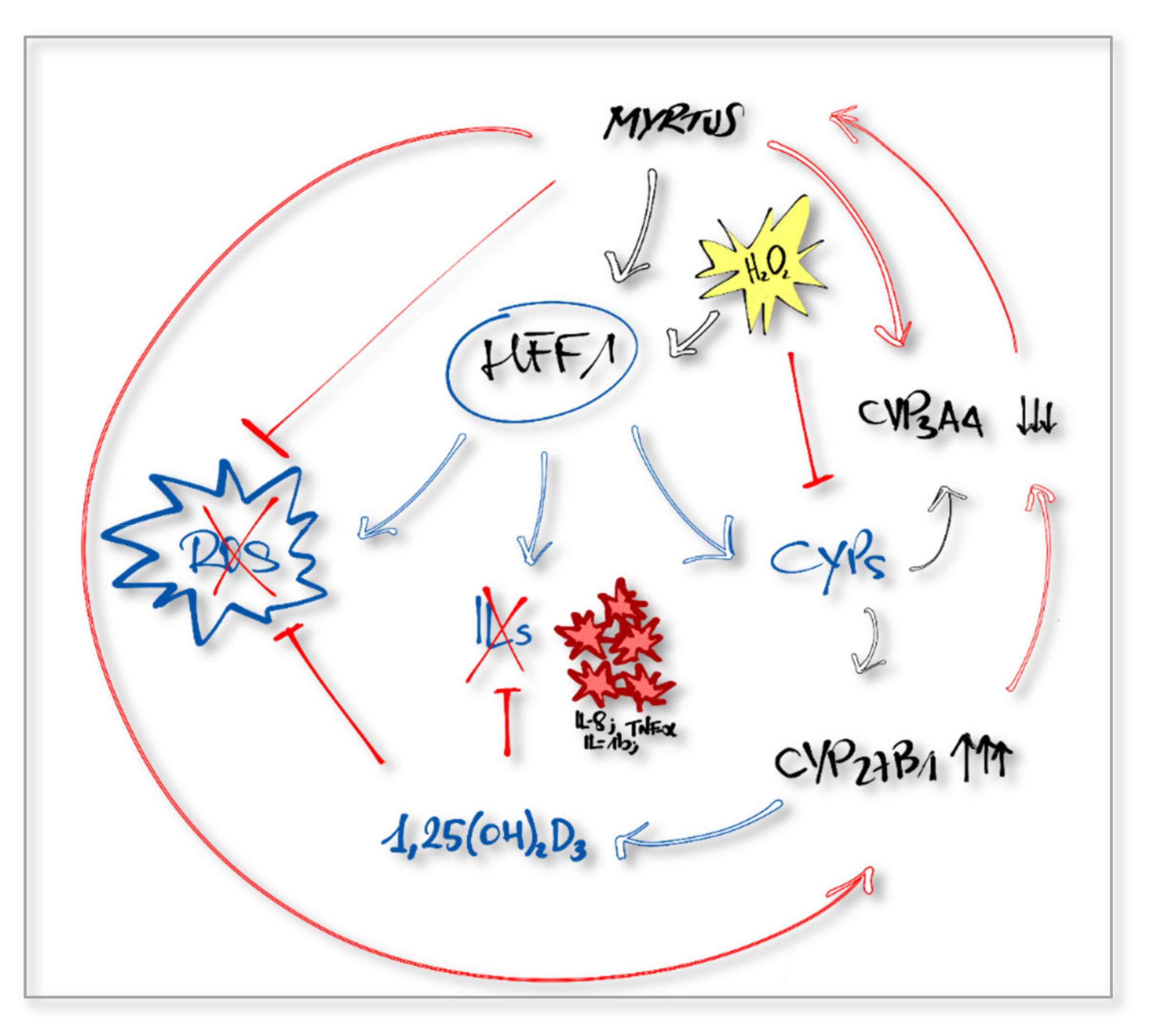
3. Discussion
4. Materials and Methods
4.1. Reagents and Solvents
4.1.1. Preparation of the Biomass Extracts
4.1.2. Stock Standard Solutions of Polyphenols
4.1.3. HPLC Analysis
4.1.4. Hydroxyl Radical Scavenging Activity
4.2. Cell Culturing Conditions
4.3. MTT Viability Assay
4.4. Measuring of ROS Production
4.5. Gene Expression Analysis by Real Time-PCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Kotas, M.E.; Medzhitov, R. Homeostasis, Inflammation, and Disease Susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Neri, M.; Fineschi, V.; Paolo, M.; Pomara, C.; Riezzo, I.; Turillazzi, E.; Cerretani, D. Cardiac Oxidative Stress and Inflammatory Cytokines Response after Myocardial Infarction. Curr. Vasc. Pharmacol. 2015, 13, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P. The inflammation theory of disease. the growing realization that chronic inflammation is crucial in many diseases opens new avenues for treatment. EMBO Rep. 2012, 13, 968–970. [Google Scholar] [CrossRef]
- Wojdasiewicz, P.; Poniatowski Łukasz, A.; Szukiewicz, D. The Role of Inflammatory and Anti-Inflammatory Cytokines in the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2014, 2014, 1–19. [Google Scholar] [CrossRef] [PubMed]
- McArdle, M.A.; Finucane, O.M.; Connaughton, R.M.; McMorrow, A.M.; Roche, H.M. Mechanisms of Obesity-Induced Inflammation and Insulin Resistance: Insights into the Emerging Role of Nutritional Strategies. Front. Endocrinol. 2013, 4. [Google Scholar] [CrossRef]
- Ponath, G.; Park, C.; Pitt, D. The Role of Astrocytes in Multiple Sclerosis. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Sand, J.; Haertel, E.; Biedermann, T.; Contassot, E.; Reichmann, E.; French, L.E.; Werner, S.; Beer, H.-D. Expression of inflammasome proteins and inflammasome activation occurs in human, but not in murine keratinocytes. Cell Death 2018, 9, 24. [Google Scholar] [CrossRef]
- Eckmann, L.; Kagnoff, M.F.; Fierer, J. Epithelial cells secrete the chemokine interleukin-8 in response to bacterial entry. Infect. Immun. 1993, 61, 4569–4574. [Google Scholar] [PubMed]
- Gresnigt, M.S.; Joosten, L.A.B.; Verschueren, I.; Van Der Meer, J.W.M.; Netea, M.G.; Dinarello, C.A.; Van De Veerdonk, F.L. Neutrophil-Mediated Inhibition of Proinflammatory Cytokine Responses. J. Immunol. 2012, 189, 4806–4815. [Google Scholar] [CrossRef]
- Wright, T.M. Cytokines in acute and chronic inflammation. Front. Biosci. 1997, 2, d12–d26. [Google Scholar] [CrossRef]
- Zhang, J.-M.; An, J. Cytokines, Inflammation, and Pain. Int. Anesthesiol. Clin. 2007, 45, 27. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Scandalios, J. Oxidative stress: Molecular perception and transduction of signals triggering antioxidant gene defenses. Braz. J. Med. Boil. Res. 2005, 38, 995–1014. [Google Scholar] [CrossRef]
- Anannarukan, N.; Niwattisaiwong, N.; Warisnoicharoen, W.; Winitthana, T.; Pramyothin, P.; Chaichantipyuth, C.; Lawanprasert, S. Inhibition of human cytochrome P450 in vitro by Phyllanthus amarus and Phyllanthus emblica aqueous extracts. Thai J. Pharm. Sci. 2012, 36, 135–143. [Google Scholar]
- Hrycay, E.G.; Bandiera, S.M. Involvement of Cytochrome P450 in Reactive Oxygen Species Formation and Cancer. Adv. Pharmacol. 2015, 74, 35–84. [Google Scholar]
- Nagai, F.; Kato, E.; Tamura, H.-O. Oxidative Stress Induces GSTP1 and CYP3A4 Expression in the Human Erythroleukemia Cell Line, K562. Boil. Pharm. Bull. 2004, 27, 492–495. [Google Scholar] [CrossRef]
- Gröschel, C.; Tennakoon, S.; Kállay, E. Cytochrome P450 Function and Pharmacological Roles in Inflammation and Cancer. Adv. Pharmacol. (San Diego, Calif.) 2015, 74, 413–458. [Google Scholar] [CrossRef]
- Banerjee, S.; Sil, J.G.A.P.C. Drug Metabolism and Oxidative Stress: Cellular Mechanism and New Therapeutic Insights. Biochem. Anal. Biochem. 2019, 5, 255. [Google Scholar] [CrossRef]
- Sakaki, T.; Kagawa, N.; Yamamoto, K.; Inouye, K. Metabolism of vitamin D3 by cytochromes P450. Front Biosci 2005, 10, 119–134. [Google Scholar]
- Eichelbaum, M.; Burk, O. CYP3A genetics in drug metabolism. Nat. Med. 2001, 7, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.C.; Hines, R.N.; Gu, C.; Koukouritaki, S.B.; Manro, J.R.; Tandler, P.J.; Zaya, M.J. Developmental Expression of the Major Human Hepatic CYP3A Enzymes. J. Pharmacol. Exp. Ther. 2003, 307, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Basheer, L.; Kerem, Z. Interactions between CYP3A4 and Dietary Polyphenols. Oxid. Med. Cell. Longev. 2015, 2015, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.L.; Edson, K.Z.; Totah, R.A.; Rettie, A.E. Cytochrome P450 ω-Hydroxylases in Inflammation and Cancer. In HIV-1: Molecular Biology and Pathogenesis; Elsevier BV: Amsterdam, The Netherlands, 2015; Volume 74, pp. 223–262. [Google Scholar] [CrossRef]
- Korobkova, E.A. Effect of Natural Polyphenols on CYP Metabolism: Implications for Diseases. Chem. Res. Toxicol. 2015, 28. [Google Scholar] [CrossRef]
- Singh, U.; Jialal, I. Oxidative stress and atherosclerosis. Pathophysiology 2006, 13, 129–142. [Google Scholar] [CrossRef]
- Wang, X.; Wang, W.; Li, L.; Perry, G.; Lee, H.-G.; Zhu, X. Oxidative stress and mitochondrial dysfunction in Alzheimer’s disease. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2014, 1842, 1240–1247. [Google Scholar] [CrossRef]
- Lenaz, G.; Bovina, C.; D’Aurelio, M.; Fato, R.; Formiggini, G.; Genova, M.L.; Giuliano, G.; Pich, M.M.; Paolucci, U.; Castelli, G.P.; et al. Role of Mitochondria in Oxidative Stress and Aging. Ann. N. Y. Acad. Sci. 2002, 959, 199–213. [Google Scholar] [CrossRef]
- Pohl, F.; Lin, P.K.T.; Ferreira, I.C. The Potential Use of Plant Natural Products and Plant Extracts with Antioxidant Properties for the Prevention/Treatment of Neurodegenerative Diseases: In Vitro, In Vivo and Clinical Trials. Molecules 2018, 23, 3283. [Google Scholar] [CrossRef]
- Maioli, M.; Basoli, V.; Carta, P.; Fabbri, D.; Dettori, M.A.; Cruciani, S.; Serra, P.A.; Delogu, G. Synthesis of magnolol and honokiol derivatives and their effect against hepatocarcinoma cells. PLoS ONE 2018, 13, e0192178. [Google Scholar] [CrossRef]
- Ferhi, S.; Santaniello, S.; Zerizer, S.; Cruciani, S.; Fadda, A.; Sanna, D.; Dore, A.; Maioli, M.; D’Hallewin, G.; Fabiani, R. Total Phenols from Grape Leaves Counteract Cell Proliferation and Modulate Apoptosis-Related Gene Expression in MCF-7 and HepG2 Human Cancer Cell Lines. Molecules 2019, 24, 612. [Google Scholar] [CrossRef]
- Coșarcă, S.-L.; Moacă, E.-A.; Tanase, C.; Muntean, D.L.; Pavel, I.Z.; Dehelean, C.A. Spruce and beech bark aqueous extracts: Source of polyphenols, tannins and antioxidants correlated to in vitro antitumor potential on two different cell lines. Wood Sci. Technol. 2018, 53, 313–333. [Google Scholar] [CrossRef]
- Dróżdż, P.; Šėžienė, V.; Pyrzynska, K. Phytochemical Properties and Antioxidant Activities of Extracts from Wild Blueberries and Lingonberries. Foods Hum. Nutr. 2017, 72, 360–364. [Google Scholar] [CrossRef]
- Stankovic, M.S.; Niciforovic, N.; Topuzovic, M.; Solujic, S. Total phenolic content, flavonoid concentrations and antioxidant activity, of the whole plant and plant parts extracts from Teucrium montanum L. var. montanum, f. supinum (L.) reichenb. Biotechnol. Biotechnol. Equip. 2011, 25, 2222–2227. [Google Scholar] [CrossRef]
- Rebaya, A.; Belghith, S.I.; Baghdikian, B.; Leddet, V.M.; Mabrouki, F.; Olivier, E.; Cherif, J.K.; Ayadi, M.T. Total Phenolic, Total Flavonoid, Tannin Content, and Antioxidant Capacity of Halimium halimifolium (Cistaceae). J. Appl. Pharm. Sci. 2014, 5, 52–57. [Google Scholar] [CrossRef]
- Li, H.; Deng, Z.; Liu, R.; Zhu, H.; Draves, J.; Marcone, M.; Sun, Y.; Tsao, R. Characterization of phenolics, betacyanins and antioxidant activities of the seed, leaf, sprout, flower and stalk extracts of three Amaranthus species. J. Compos. Anal. 2015, 37, 75–81. [Google Scholar] [CrossRef]
- Hennia, A.; Miguel, M.; Nemmiche, S. Antioxidant Activity of Myrtus communis L. and Myrtus nivellei Batt. & Trab. Extracts: A Brief Review. Medicines 2018, 5, 89. [Google Scholar] [CrossRef]
- Romani, A.; Coinu, R.; Carta, S.; Pinelli, P.; Galardi, C.; Vincieri, F.F.; Franconi, F. Evaluation of Antioxidant Effect of Different Extracts ofMyrtus communisL. Free. Radic. Res. 2004, 38, 97–103. [Google Scholar] [CrossRef]
- Sisay, M.; Gashaw, T. Ethnobotanical, Ethnopharmacological, and Phytochemical Studies of Myrtus communis Linn: A Popular Herb in Unani System of Medicine. J. Evid.-Based Complement. Altern. Med. 2017, 22, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxid. Med. Cell. Longev. 2016, 2016, 1–9. [Google Scholar] [CrossRef]
- Sorice, A.; Guerriero, E.; Capone, F.; Colonna, G.; Castello, G.; Costantini, S. Ascorbic Acid: Its Role in Immune System and Chronic Inflammation Diseases. Mini-Rev. Med. Chem. 2014, 14, 444–452. [Google Scholar] [CrossRef]
- Artis, D. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat. Rev. Immunol. 2008, 8, 411–420. [Google Scholar] [CrossRef]
- Nichols, J.A.; Katiyar, S.K. Skin photoprotection by natural polyphenols: Anti-inflammatory, antioxidant and DNA repair mechanisms. Arch. Dermatol. Res. 2010, 302, 71–83. [Google Scholar] [CrossRef]
- Bickers, D.R.; Athar, M. Oxidative Stress in the Pathogenesis of Skin Disease. J. Investig. Dermatol. 2006, 126, 2565–2575. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta (BBA)—Bioenerg. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Semak, I.; Zbytek, B.; Pisarchik, A.; Li, W.; Zjawiony, J.; Tuckey, R.C. Cytochromes P450 and Skin Cancer: Role of Local Endocrine Pathways. Anti-Cancer Agents Med. Chem. 2014, 14, 77–96. [Google Scholar] [CrossRef]
- Gašperlin, M.; Gosenca, M. Main approaches for delivering antioxidant vitamins through the skin to prevent skin ageing. Opin. Drug Deliv. 2011, 8, 905–919. [Google Scholar] [CrossRef] [PubMed]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S.; Esa, N.M. Role of Antioxidants and Natural Products in Inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 1–15. [Google Scholar] [CrossRef]
- Valério, D.A.; Georgetti, S.R.; Magro, D.A.; Casagrande, R.; Cunha, T.M.; Vicentini, F.T.M.C.; Vieira, S.M.; Fonseca, M.J.V.; Ferreira, S.H.; Cunha, F.Q.; et al. Quercetin Reduces Inflammatory Pain: Inhibition of Oxidative Stress and Cytokine Production. J. Nat. Prod. 2009, 72, 1975–1979. [Google Scholar] [CrossRef]
- Kang, S.R.; Park, K.I.; Park, H.S.; Lee, D.H.; A Kim, J.; Nagappan, A.; Kim, E.H.; Lee, W.S.; Shin, S.C.; Park, M.K.; et al. Anti-inflammatory effect of flavonoids isolated from Korea Citrus aurantium L. on lipopolysaccharide-induced mouse macrophage RAW 264.7 cells by blocking of nuclear factor-kappa B (NF-κB) and mitogen-activated protein kinase (MAPK) signalling pathways. Food Chem. 2011, 129, 1721–1728. [Google Scholar] [CrossRef]
- Melito, S.; Chessa, I.; Erre, P.; Podani, J.; Mulas, M. The genetic diversity of Sardinian myrtle (Myrtus communis L.) populations. Electron. J. Biotechnol. 2013, 16. [Google Scholar] [CrossRef]
- Montoro, P.; Tuberoso, C.I.; Piacente, S.; Perrone, A.; De Feo, V.; Cabras, P.; Pizza, C. Stability and antioxidant activity of polyphenols in extracts of Myrtus communis L. berries used for the preparation of myrtle liqueur. J. Pharm. Biomed. Anal. 2006, 41, 1614–1619. [Google Scholar] [CrossRef] [PubMed]
- Zilkah, S.; Goldschdmidt, E.E. Myrtle (Myrtus communis L.)—A Native Mediterranean and Cultured Crop Species. In Medicinal and Aromatic Plants of the Middle-East; Springer: Dordrecht, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Ghorbani, A.; Esmaeilizadeh, M. Pharmacological properties of Salvia officinalis and its components. J. Tradit. Complement. Med. 2017, 7, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.-N.; Zhang, Y.; Cui, Y.-L.; Akinyi, O.M. Comparison of Paeoniflorin and Albiflorin on Human CYP3A4 and CYP2D6. Evid.-Based Complement. Altern. Med. 2015, 2015, 1–7. [Google Scholar] [CrossRef]
- Sieniawska, E.; Baj, T.; Dudka, J.; Gieroba, R.; Swiatek, L.; Rajtar, B.; Glowniak, K.; Polz-Dacewicz, M. Cytotoxicity, antioxidant activity and an effect on CYP3A4 and CYP2D6 of Mutellina purpurea L. extracts. Chem. Toxicol. 2013, 52, 188–192. [Google Scholar] [CrossRef]
- Karakurt, S. Modulatory effects of rutin on the expression of cytochrome P450s and antioxidant enzymes in human hepatoma cells. Acta Pharm. 2016, 66, 491–502. [Google Scholar] [CrossRef]
- Sumantran, V.N.; Mishra, P.; Bera, R.; Sudhakar, N.; Hong, H. Microarray Analysis of Differentially-Expressed Genes Encoding CYP450 and Phase II Drug Metabolizing Enzymes in Psoriasis and Melanoma. Pharmaceutics 2016, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Cytochrome P450s and other enzymes in drug metabolism and toxicity. AAPS J. 2006, 8, E101–E111. [Google Scholar] [CrossRef] [PubMed]
- Schuster, I. Cytochromes P450 are essential players in the vitamin D signaling system. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2011, 1814, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Chirumbolo, S.; Bjørklund, G.; Sboarina, A.; Vella, A. The Role of Vitamin D in the Immune System as a Pro-survival Molecule. Clin. Ther. 2017, 39, 894–916. [Google Scholar] [CrossRef]
- Peng, X.; Vaishnav, A.; Murillo, G.; Alimirah, F.; Torres, K.E.; Mehta, R.G. Protection against cellular stress by 25-hydroxyvitamin D3 in breast epithelial cells. J. Cell. Biochem. 2010, 110, 1324–1333. [Google Scholar] [CrossRef] [PubMed]
- Brozyna, A.A.; Józwicki, W.; Jochymski, C.; Slominski, A.T. Decreased expression of CYP27B1 correlates with the increased aggressiveness of ovarian carcinomas. Oncol. Rep. 2015, 33, 599–606. [Google Scholar] [CrossRef]
- Bikle, D.; Chang, S.; Crumrine, D.; Elalieh, H.; Man, M.-Q.; Choi, E.; Dardenne, O.; Xie, Z.; Arnaud, R.S.; Feingold, K.; et al. 25 Hydroxyvitamin D 1 α-Hydroxylase Is Required for Optimal Epidermal Differentiation and Permeability Barrier Homeostasis. J. Investig. Dermatol. 2004, 122, 984–992. [Google Scholar] [CrossRef]
- Piotrowska, A.; Wierzbicka, J.; Ślebioda, T.; Woźniak, M.; Tuckey, R.C.; Slominski, A.T.; Żmijewski, M.A.; Anna, P.; Justyna, W.; Tomasz, Ś.; et al. Vitamin D derivatives enhance cytotoxic effects of H2O2 or cisplatin on human keratinocytes. Steroids 2016, 110, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.-L.; Yeh, J.K.; Cao, J.J.; Tatum, O.L.; Dagda, R.Y.; Wang, J.-S. Synergistic effects of green tea polyphenols and alphacalcidol on chronic inflammation-induced bone loss in female rats. Osteoporos. Int. 2010, 21, 1841–1852. [Google Scholar] [CrossRef] [PubMed]
- Zargar, S.; Al-Majed, A.-R.A.; Wani, T.A. Potentiating and synergistic effect of grapefruit juice on the antioxidant and anti-inflammatory activity of aripiprazole against hydrogen peroxide induced oxidative stress in mice. BMC Complement. Altern. Med. 2018, 18, 106. [Google Scholar] [CrossRef]
- Sarais, G.; D’Urso, G.; Lai, C.; Pirisi, F.M.; Pizza, C.; Montoro, P. Targeted and Untargeted Mass Spectrometric Approaches in Discrimination between Myrtus Communis Cultivars from Sardinia Region. J. Mass Spectrom. 2016, 51, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Fadda, A.; Barberis, A.; Sanna, D. Influence of pH, buffers and role of quinolinic acid, a novel iron chelating agent, in the determination of hydroxyl radical scavenging activity of plant extracts by Electron Paramagnetic Resonance (EPR). Food Chem. 2018, 240, 174–182. [Google Scholar] [CrossRef]
Sample Availability: Extracts of the compounds are available from the authors upon request. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | Concentration mg/kg | |
|---|---|---|
| Ethanol/Water | ||
| Pulp | Seeds | |
| Gallic acid | 52.2 ± 0.9 | 137.0 ± 6.8 |
| Hydrolysable tannins a | 498.0 ± 20.5 | 11989.8 ± 205.2 |
| Ellagic acid | 350.5 ± 15.0 | 726.9 ± 28.3 |
| Flavonols | ||
| Quercetin-3-O-galactoside | 191.0 ± 6.7 | 104.9 ± 9.3 |
| Quercetin-3-O-rhamnosid | 66.6 ± 3.0 | 62.0 ± 2.9 |
| Anthocyanins | ||
| Cyanidin-3-glucoside | 1.8 ± 0.2 | nd |
| Petunidin-3-glucoside b | 3.6 ± 0.3 | nd |
| Peonidin-3-glucoside | 13.5 ± 0.3 | nd |
| Malvidin-3-glucoside | 42.0 ± 2.4 | nd |
| Primers | Forward | Reverse |
|---|---|---|
| hGAPDH | GAGTCAACGGAATTTGGTCGT | GACAAGCTTCCCGTTCTCAG |
| IL-1β | GCTACGAATCTCCGACCACC | ATCGTGCACATAAGCCTCGT |
| IL-8 | CTTCTCCACAACCCTCTG | GAACTGAGAGTGATTGAGAGT |
| TNF-α | CCTCAGACGCCACAT | GAGGGCTGATTAGAGAGA |
| VEGF-A | GCCAAGTGGTCCCAGGCTGC | TCGTCATTGCAGGCAGCCCCC |
| CYP3A4 | TAGCCCAGCAAAGAGCAACA | CAAAAGGCCTCCGGTTTGTG |
| CYP27B1 | CCTGAACCAGACCATGACCC | GAGCCTTTGCCATTCTTCGC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruciani, S.; Santaniello, S.; Garroni, G.; Fadda, A.; Balzano, F.; Bellu, E.; Sarais, G.; Fais, G.; Mulas, M.; Maioli, M. Myrtus Polyphenols, from Antioxidants to Anti-Inflammatory Molecules: Exploring a Network Involving Cytochromes P450 and Vitamin D. Molecules 2019, 24, 1515. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24081515
Cruciani S, Santaniello S, Garroni G, Fadda A, Balzano F, Bellu E, Sarais G, Fais G, Mulas M, Maioli M. Myrtus Polyphenols, from Antioxidants to Anti-Inflammatory Molecules: Exploring a Network Involving Cytochromes P450 and Vitamin D. Molecules. 2019; 24(8):1515. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24081515
Chicago/Turabian StyleCruciani, Sara, Sara Santaniello, Giuseppe Garroni, Angela Fadda, Francesca Balzano, Emanuela Bellu, Giorgia Sarais, Giacomo Fais, Maurizio Mulas, and Margherita Maioli. 2019. "Myrtus Polyphenols, from Antioxidants to Anti-Inflammatory Molecules: Exploring a Network Involving Cytochromes P450 and Vitamin D" Molecules 24, no. 8: 1515. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24081515