Identification of Patulin from Penicillium coprobium as a Toxin for Enteric Neurons


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
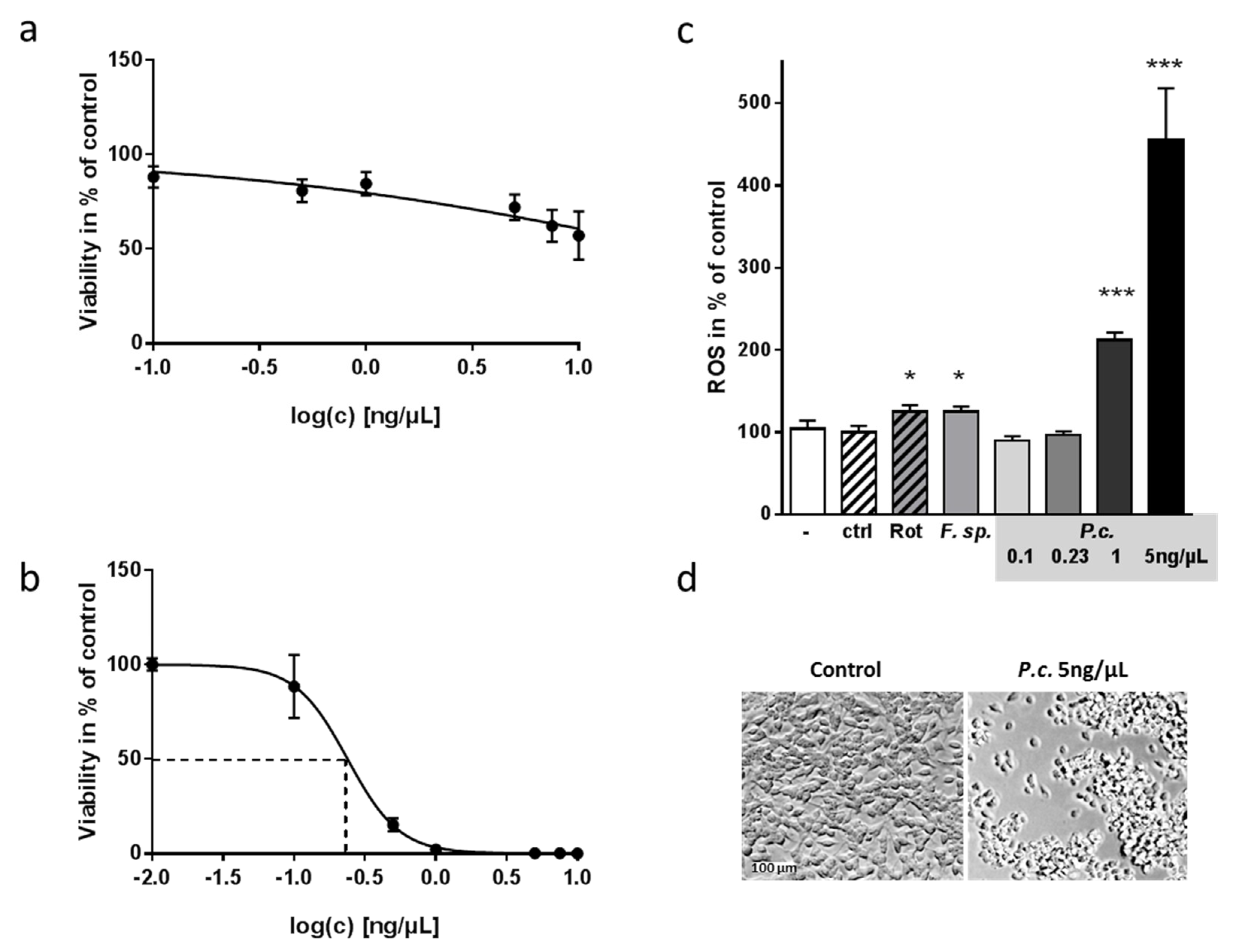
2.1. Toxic Effect of Fungal Extracts on Neuronal Cells
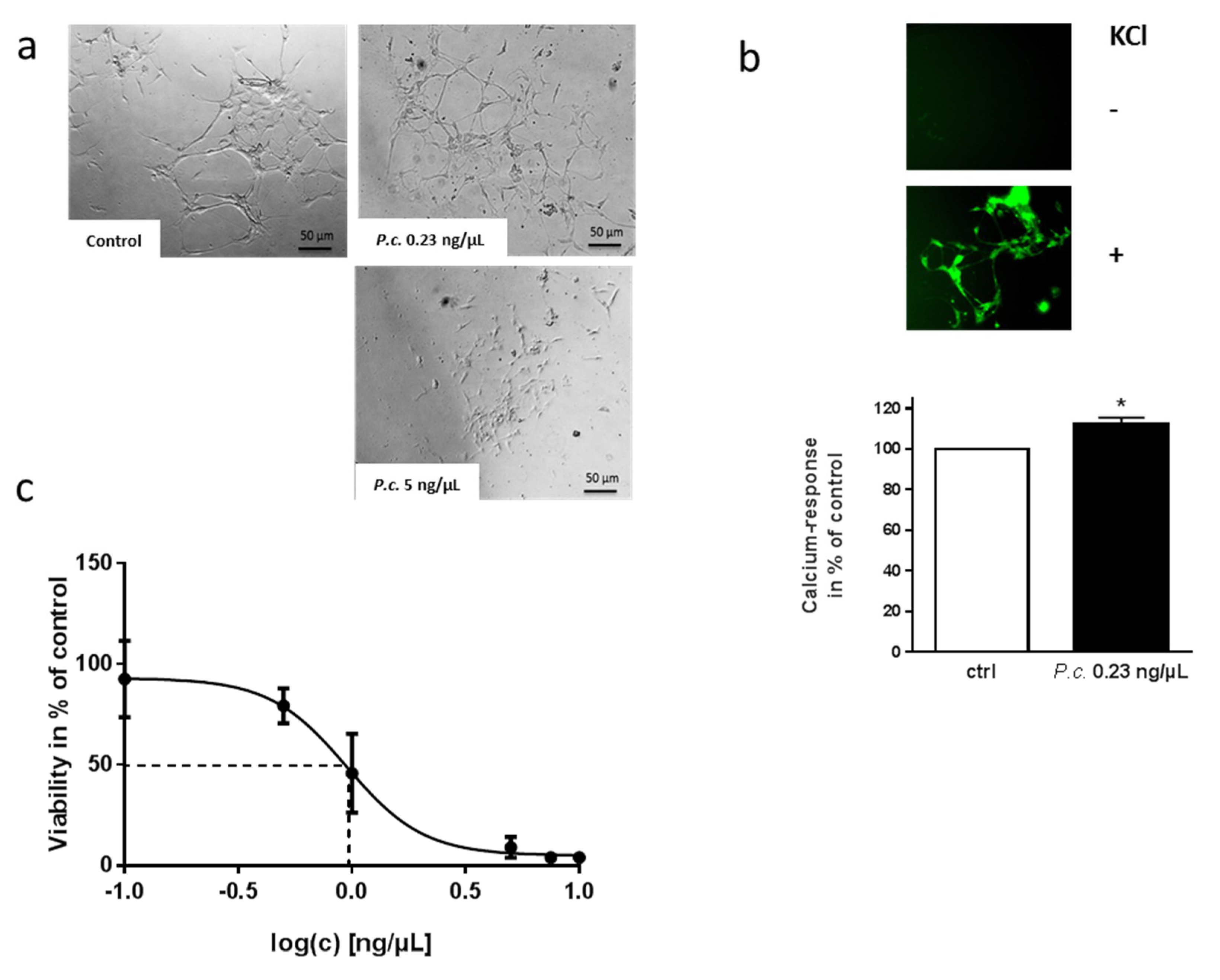
2.2. Impact of P. coprobium Extract on Primary Enteric Neurons
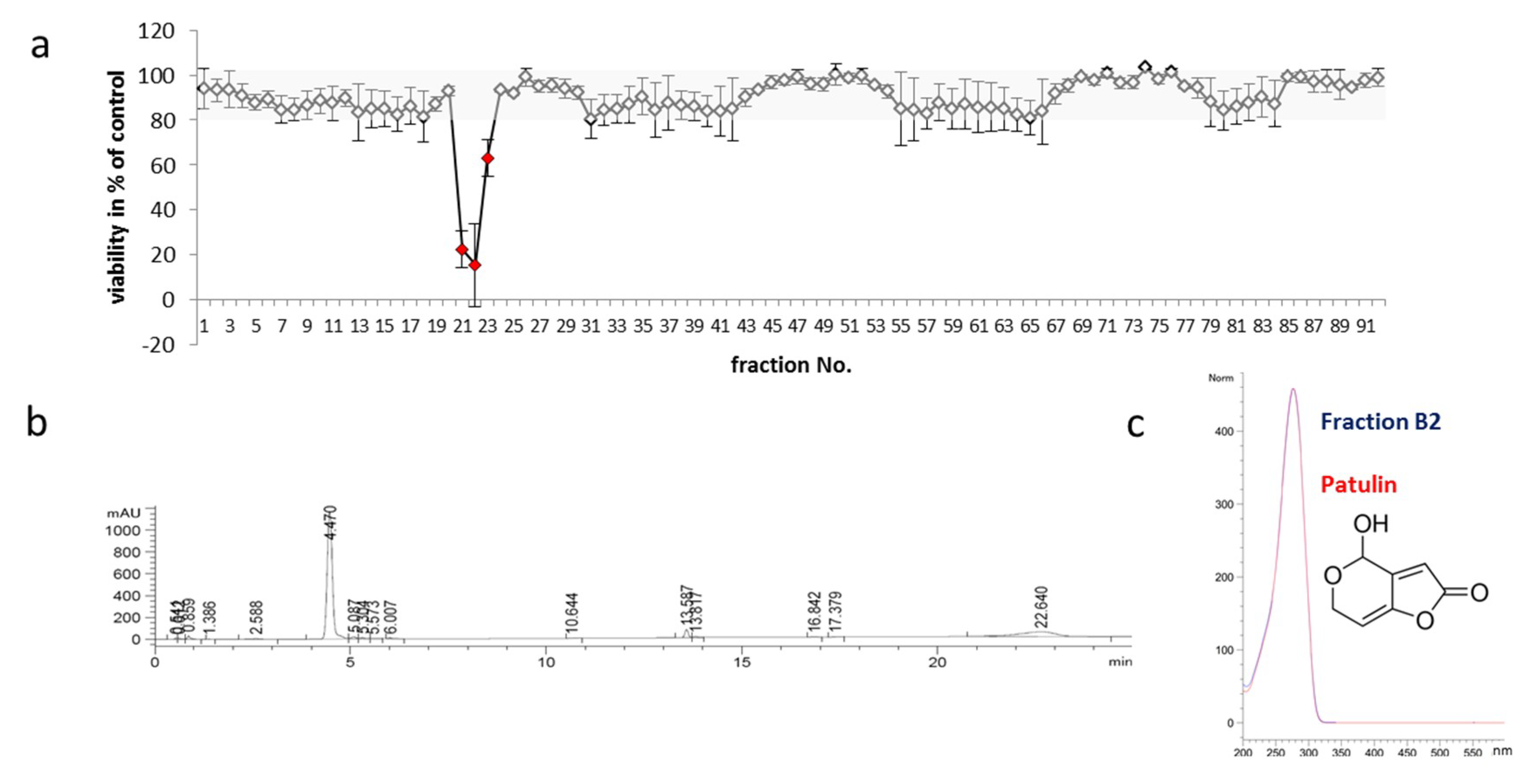
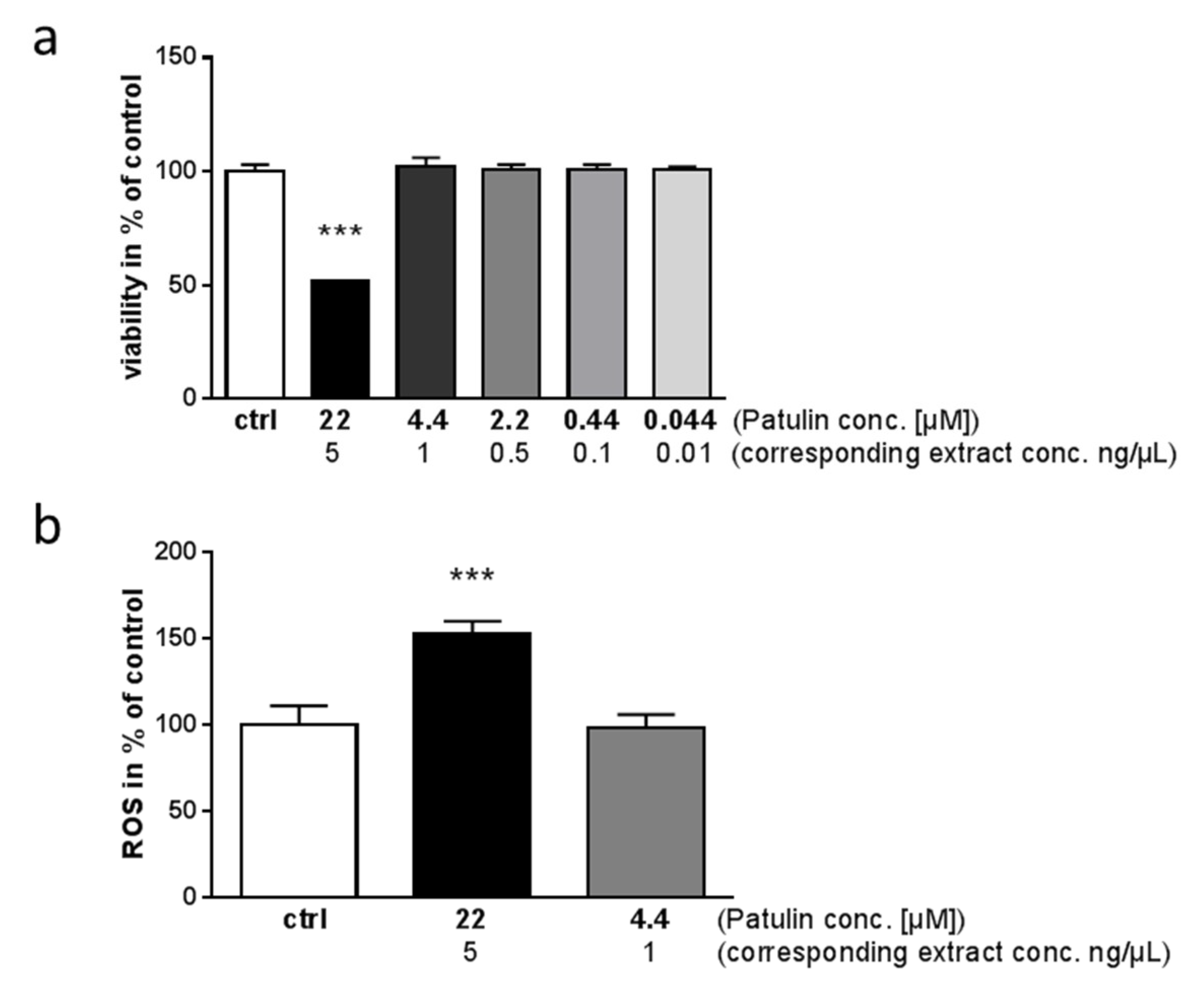
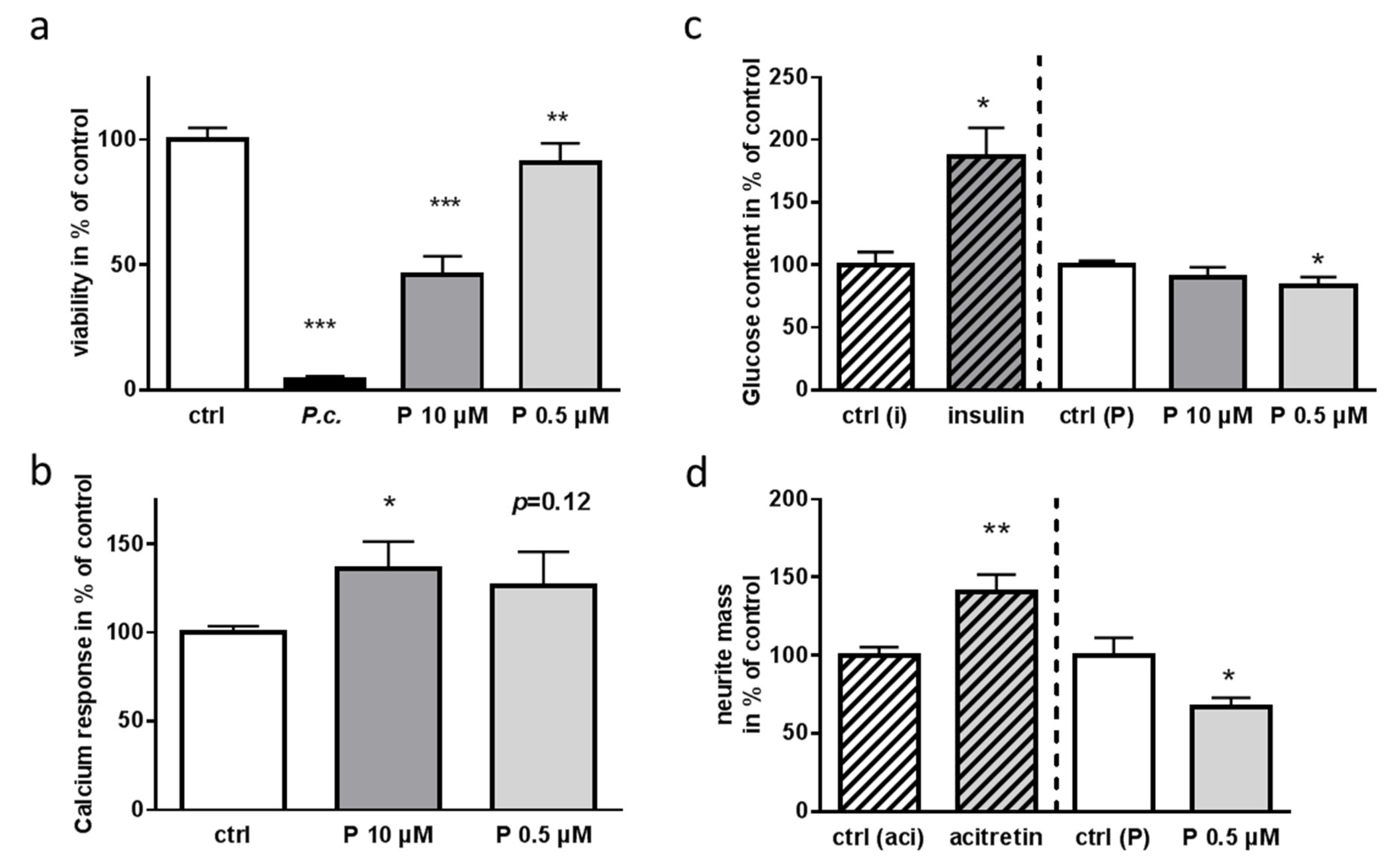
2.3. Identification of Toxic Compounds Derived from P. coprobium Extract
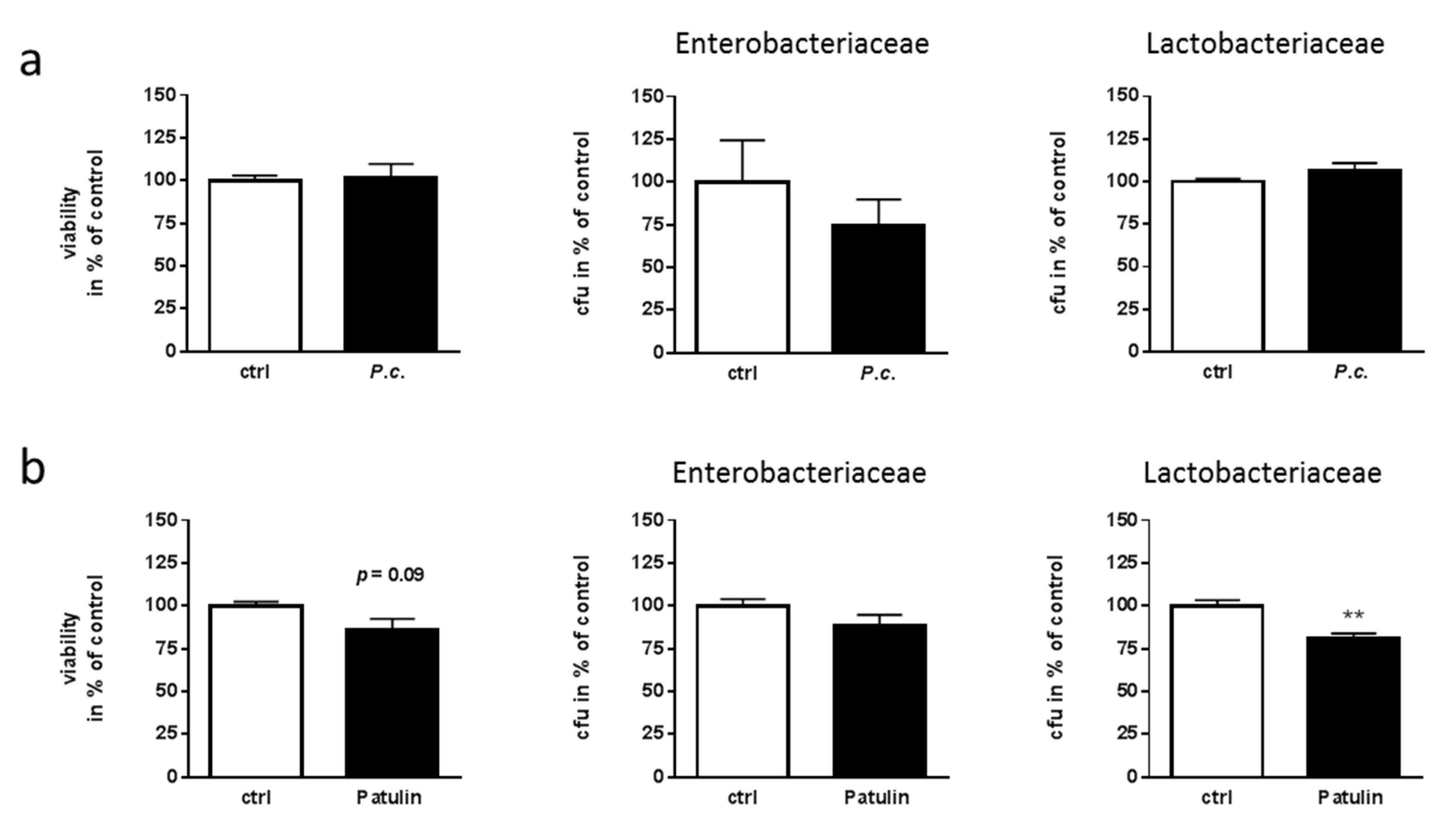
2.4. Lack of Toxicity of P. coprobium Extract on Fecal Bacteria
2.5. Impact of Patulin on Physiological Functions of Enteric Neurons
3. Discussion
4. Materials and Methods
4.1. Fungal Extracts
4.2. Cell Culture
4.3. Cytotoxicity Test
4.4. ROS Test
4.5. Calcium Signaling
4.6. Measurement of Glucose Content
4.7. Neurite Outgrowth Measurement
4.8. Growth and Viability Testing of Exemplary Murine Fecal Bacteria
4.9. Identification of P. coprobium, Fractionation of Its Extract and Verification of Patulin
Author Contributions
Funding
Conflicts of Interest
References
- Mason, K.L.; Huffnagle, G.B.; Noverr, M.C.; Kao, J.Y. Overview of gut immunology. Adv. Exp. Med. Biol. 2008, 635, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bayliss, W.M.; Starling, E.H. The movements and innervation of the small intestine. J. Physiol. 1901, 26, 125–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, R.K.; Hirano, I. The enteric nervous system. N. Engl. J. Med. 1996, 334, 1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. Adv. Exp. Med. Biol. 2014, 817, 39–71. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. Types of neurons in the enteric nervous system. J. Auton. Nerv. Syst. 2000, 81, 87–96. [Google Scholar] [CrossRef]
- Breit, S.; Kupferberg, A.; Rogler, G.; Hasler, G. Vagus Nerve as Modulator of the Brain-Gut Axis in Psychiatric and Inflammatory Disorders. Front Psychiatry 2018, 9, 44. [Google Scholar] [CrossRef]
- Human Microbiome Project, C. A framework for human microbiome research. Nature 2012, 486, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Human Microbiome Project, C. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Mar Rodriguez, M.; Perez, D.; Javier Chaves, F.; Esteve, E.; Marin-Garcia, P.; Xifra, G.; Vendrell, J.; Jove, M.; Pamplona, R.; Ricart, W.; et al. Obesity changes the human gut mycobiome. Sci. Rep. 2015, 5, 14600. [Google Scholar] [CrossRef]
- Underhill, D.M.; Iliev, I.D. The mycobiota: Interactions between commensal fungi and the host immune system. Nat. Rev. Immunol. 2014, 14, 405–416. [Google Scholar] [CrossRef]
- Hallen-Adams, H.E.; Kachman, S.D.; Kim, J.; Legge, R.M.; Martinez, I. Fungi inhabiting the healthy human gastrointestinal tract: A diverse and dynamic community. Fungal. Ecol. 2015, 15, 9–17. [Google Scholar] [CrossRef]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.T.; Shao, T.Y.; Ang, W.X.G.; Kinder, J.M.; Turner, L.H.; Pham, G.; Whitt, J.; Alenghat, T.; Way, S.S. Commensal Fungi Recapitulate the Protective Benefits of Intestinal Bacteria. Cell Host Microbe 2017, 22, 809–816.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.; Metzke, D.; Schmitz, J.; Dorffel, Y.; Baumgart, D.C. Anti-inflammatory effects of Saccharomyces boulardii mediated by myeloid dendritic cells from patients with Crohn’s disease and ulcerative colitis. Am. J. Physiol. Gastrointest Liver Physiol. 2011, 301, G1083–G1092. [Google Scholar] [CrossRef] [PubMed]
- Botschuijver, S.; Roeselers, G.; Levin, E.; Jonkers, D.M.; Welting, O.; Heinsbroek, S.E.M.; de Weerd, H.H.; Boekhout, T.; Fornai, M.; Masclee, A.A.; et al. Intestinal Fungal Dysbiosis Is Associated with Visceral Hypersensitivity in Patients with Irritable Bowel Syndrome and Rats. Gastroenterology 2017, 153, 1026–1039. [Google Scholar] [CrossRef] [PubMed]
- Mckenzie, H.; Main, J.; Pennington, C.R.; Parratt, D. Antibody to Selected Strains of Saccharomyces-Cerevisiae (Bakers and Brewers-Yeast) and Candida-Albicans in Crohns-Disease. Gut 1990, 31, 536–538. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.P.; Jegou, S.; Landman, C.; Cohen, D.; Liguori, G.; Bourrier, A.; Nion-Larmurier, I.; et al. Fungal microbiota dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Standaert-Vitse, A.; Sendid, B.; Joossens, M.; Francois, N.; Vandewalle-El Khoury, P.; Branche, J.; Van Kruiningen, H.; Jouault, T.; Rutgeerts, P.; Gower-Rousseau, C.; et al. Candida albicans Colonization and ASCA in Familial Crohn’s Disease. Am. J. Gastroenterol. 2009, 104, 1745–1753. [Google Scholar] [CrossRef] [PubMed]
- Kovalevich, J.; Langford, D. Considerations for the use of SH-SY5Y neuroblastoma cells in neurobiology. Methods Mol. Biol. 2013, 1078, 9–21. [Google Scholar] [CrossRef]
- Schuck, F.; Schmitt, U.; Reinhardt, S.; Freese, C.; Lee, I.S.; Thines, E.; Efferth, T.; Endres, K. Extract of Caragana sinica as a potential therapeutic option for increasing alpha-secretase gene expression. Phytomedicine 2015, 22, 1027–1036. [Google Scholar] [CrossRef]
- Reinhardt, S.; Stoye, N.; Luderer, M.; Kiefer, F.; Schmitt, U.; Lieb, K.; Endres, K. Identification of disulfiram as a secretase-modulating compound with beneficial effects on Alzheimer’s disease hallmarks. Sci. Rep. 2018, 8, 1329. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, A.; Haney, S.A.; Bowman, D. Designing Metrics for High Content Assays. In Introduction to High Content Screening: Imaging Technology, Assay Development, and Data Analysis in Biology and Drug Discovery; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 115–129. [Google Scholar]
- de Abreu Costa, L.; Henrique Fernandes Ottoni, M.; Dos Santos, M.G.; Meireles, A.B.; Gomes de Almeida, V.; de Fatima Pereira, W.; Alves de Avelar-Freitas, B.; Eustaquio Alvim Brito-Melo, G. Dimethyl Sulfoxide (DMSO) Decreases Cell Proliferation and TNF-alpha, IFN-gamma, and IL-2 Cytokines Production in Cultures of Peripheral Blood Lymphocytes. Molecules 2017, 22, 1789. [Google Scholar] [CrossRef] [PubMed]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.H.; Wu, T.S.; Yu, F.Y.; Su, C.C. Induction of oxidative stress response by the mycotoxin patulin in mammalian cells. Toxicol. Sci. 2007, 95, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Ragheb, K.; Lawler, G.; Sturgis, J.; Rajwa, B.; Melendez, J.A.; Robinson, J.P. Mitochondrial complex I inhibitor rotenone induces apoptosis through enhancing mitochondrial reactive oxygen species production. J. Biol. Chem. 2003, 278, 8516–8525. [Google Scholar] [CrossRef] [PubMed]
- Bassotti, G.; Villanacci, V.; Maurer, C.A.; Fisogni, S.; Di Fabio, F.; Cadei, M.; Morelli, A.; Panagiotis, T.; Cathomas, G.; Salerni, B. The role of glial cells and apoptosis of enteric neurones in the neuropathology of intractable slow transit constipation. Gut 2006, 55, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Lerdkrai, C.; Asavapanumas, N.; Brawek, B.; Kovalchuk, Y.; Mojtahedi, N.; Olmedillas Del Moral, M.; Garaschuk, O. Intracellular Ca(2+) stores control in vivo neuronal hyperactivity in a mouse model of Alzheimer’s disease. Proc. Natl. Acad. Sci. U S A 2018, 115, E1279–E1288. [Google Scholar] [CrossRef]
- Frisvad, J.C. A critical review of producers of small lactone mycotoxins: Patulin, penicillic acid and moniliformin. World Mycotoxin. J. 2018, 11, 73–100. [Google Scholar] [CrossRef]
- Pal, S.; Singh, N.; Ansari, K.M. Toxicological effects of patulin mycotoxin on the mammalian system: An overview. Toxicol Res. (Camb) 2017, 6, 764–771. [Google Scholar] [CrossRef]
- Endres, K.; Schafer, K.H. Influence of Commensal Microbiota on the Enteric Nervous System and Its Role in Neurodegenerative Diseases. J. Innate Immun. 2018, 10, 172–180. [Google Scholar] [CrossRef]
- Kaptchuk, T.J.; Kerr, C.E. Commentary: Unbiased divination, unbiased evidence, and the patulin clinical trial. Int. J. Epidemiol. 2004, 33, 247–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahfoud, R.; Maresca, M.; Garmy, N.; Fantini, J. The mycotoxin patulin alters the barrier function of the intestinal epithelium: Mechanism of action of the toxin and protective effects of glutathione. Toxicol. Appl. Pharmacol. 2002, 181, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Mohan, H.M.; Collins, D.; Maher, S.; Walsh, E.G.; Winter, D.C.; O’Brien, P.J.; Brayden, D.J.; Baird, A.W. The mycotoxin patulin increases colonic epithelial permeability in vitro. Food Chem. Toxicol. 2012, 50, 4097–4102. [Google Scholar] [CrossRef]
- Lupescu, A.; Jilani, K.; Zbidah, M.; Lang, F. Patulin-induced suicidal erythrocyte death. Cell Physiol. Biochem. 2013, 32, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Singh, N. Mechanism of antifungal action of patulin. 1966. [Google Scholar]
- Liu, M.; Seino, S.; Kirchgessner, A.L. Identification and characterization of glucoresponsive neurons in the enteric nervous system. J. Neurosci. 1999, 19, 10305–10317. [Google Scholar] [CrossRef] [PubMed]
- Pfannkuche, H.; Gabel, G. Glucose, epithelium, and enteric nervous system: Dialogue in the dark. J. Anim. Physiol. Anim. Nutr. (Berl) 2009, 93, 277–286. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Sendid, B.; Hoarau, G.; Colombel, J.F.; Poulain, D.; Ghannoum, M.A. Mycobiota in gastrointestinal diseases. Nat. Rev. Gastro Hepat 2015, 12, 77–87. [Google Scholar] [CrossRef]
- Noverr, M.C.; Huffnagle, G.B. Regulation of Candida albicans morphogenesis by fatty acid metabolites. Infect Immun. 2004, 72, 6206–6210. [Google Scholar] [CrossRef]
- Enaud, R.; Vandenborght, L.E.; Coron, N.; Bazin, T.; Prevel, R.; Schaeverbeke, T.; Berger, P.; Fayon, M.; Lamireau, T.; Delhaes, L. The Mycobiome: A Neglected Component in the Microbiota-Gut-Brain Axis. Microorganisms 2018, 6, 22. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 49. [Google Scholar] [CrossRef]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens-the IARC Monographs classification. Mycotoxin. Res. 2017, 33, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.; Wang, L.; Lee, H.B.; Frisvad, J.C. New sections in Penicillium containing novel species producing patulin, pyripyropens or other bioactive compounds. Persoonia 2016, 36, 299–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, M.L.; Dowd, S.E.; Stephenson, C.; Steiner, J.M.; Suchodolski, J.S. Characterization of the fungal microbiome (mycobiome) in fecal samples from dogs. Vet. Med. Int. 2013, 2013, 658373. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Smedsgaard, J.; Larsen, T.O.; Samson, R.A. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud. Mycol. 2004, 49, 201–241. [Google Scholar]
- Ciegler, A.; Detroy, R.W.; Lillehoj, E.B. Patulin, Penicillic Acid, and Other Carcinogenic lactones. Ajl. Samuel. J. Microb. Toxins 1971, 6, 409–434. [Google Scholar]
- Morgavi, D.P.; Boudra, H.; Jouany, J.P.; Graviou, D. Prevention of patulin toxicity on rumen microbial fermentation by SH-containing reducing agents. J. Agric. Food Chem. 2003, 51, 6906–6910. [Google Scholar] [CrossRef] [PubMed]
- Collin, S.; Bodart, E.; Badot, C.; Bouseta, A.; Nizet, S. Identification of the main degradation products of patulin generated through heat detoxication treatments. J. Inst. Brewing 2008, 114, 167–171. [Google Scholar] [CrossRef]
- Boue, S.; Fortgang, I.; Levy, R.J., Jr.; Bhatnagar, D.; Burow, M.; Fahey, G.; Heiman, M.L. A novel gastrointestinal microbiome modulator from soy pods reduces absorption of dietary fat in mice. Obesity (Silver Spring) 2016, 24, 87–95. [Google Scholar] [CrossRef]
- Fliege, R.; Metzler, M. Electrophilic properties of patulin. N-acetylcysteine and glutathione adducts. Chem. Res. Toxicol. 2000, 13, 373–381. [Google Scholar] [CrossRef]
- Assuncao, R.; Alvito, P.; Kleiveland, C.R.; Lea, T.E. Characterization of in vitro effects of patulin on intestinal epithelial and immune cells. Toxicol. Lett. 2016, 250–251, 47–56. [Google Scholar] [CrossRef]
- Mbundi, L. Advances in Molecular Toxicology; Elsevier: Oxford, UK, 2014. [Google Scholar]
- Monteillier, A.; Allard, P.M.; Gindro, K.; Wolfender, J.L.; Cuendet, M. Lung Cancer Chemopreventive Activity of Patulin Isolated from Penicillium vulpinum. Molecules 2018, 23, 636. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.-Y.; Yu, J. Sensory Nerves in Lung and Airways. Wiley Online Library: Compr Physiol. 2014, 4. [Google Scholar]
- Smith, T.H.; Ngwainmbi, J.; Grider, J.R.; Dewey, W.L.; Akbarali, H.I. An in-vitro preparation of isolated enteric neurons and glia from the myenteric plexus of the adult mouse. J. Vis. Exp. 2013. [Google Scholar] [CrossRef] [PubMed]
- Smit, M.; Leng, J.; Klemke, R.L. Assay for neurite outgrowth quantification. Biotechniques 2003, 35, 254–256. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors (AS, ET). |







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brand, B.; Stoye, N.M.; dos Santos Guilherme, M.; Nguyen, V.T.T.; Baumgaertner, J.C.; Schüffler, A.; Thines, E.; Endres, K. Identification of Patulin from Penicillium coprobium as a Toxin for Enteric Neurons. Molecules 2019, 24, 2776. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24152776
Brand B, Stoye NM, dos Santos Guilherme M, Nguyen VTT, Baumgaertner JC, Schüffler A, Thines E, Endres K. Identification of Patulin from Penicillium coprobium as a Toxin for Enteric Neurons. Molecules. 2019; 24(15):2776. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24152776
Chicago/Turabian StyleBrand, Benjamin, Nicolai M. Stoye, Malena dos Santos Guilherme, Vu Thu Thuy Nguyen, Julia C. Baumgaertner, Anja Schüffler, Eckhard Thines, and Kristina Endres. 2019. "Identification of Patulin from Penicillium coprobium as a Toxin for Enteric Neurons" Molecules 24, no. 15: 2776. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24152776