Long-Noncoding RNA (lncRNA) in the Regulation of Hypoxia-Inducible Factor (HIF) in Cancer

 , , and
, , and
Abstract
:1. Introduction
2. Methods
3. lncRNAs in the Regulation of HIF-1α
3.1. lncRNAs as Enhancers of HIF-1α Expression, Activation and Stability
3.1.1. PVT1
3.1.2. H19
3.1.3. HOTAIR
3.1.4. UCA1
3.1.5. LINK-A
3.1.6. lincRNA-p21
3.1.7. HISLA
3.1.8. GHET1
3.1.9. MIR31HG/HIFCAR
3.1.10. DANCR
3.1.11. CASC9
3.1.12. MALAT1
3.1.13. MTA2TR
3.1.14. Other lncRNAs
3.2. lncRNAs as Inhibitors of HIF-1α Expression, Activation and Stability
3.2.1. LET
3.2.2. ENST00000480739
3.2.3. CPS1-IT1
3.2.4. HITT
3.2.5. MEG3
3.2.6. IDH1-AS1
3.2.7. PIN1-v2
3.2.8. HOTAIRM1
4. lncRNA and Regulation of HIF-2α
4.1. HIF2PUT
4.2. SARCC
4.3. MALAT1
4.4. NEAT1
4.5. lincRNA-p21
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schofield, C.J.; Ratcliffe, P.J. Oxygen Sensing by HIF Hydroxylases. Nat. Rev. Mol. Cell Biol. 2004, 5, 343–354. [Google Scholar] [CrossRef]
- Loboda, A.; Jozkowicz, A.; Dulak, J. HIF-1 and HIF-2 Transcription Factors--Similar but Not Identical. Mol. Cells 2010, 29, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Kaelin, W.G., Jr.; Ratcliffe, P.J. Oxygen Sensing by Metazoans: The Central Role of the HIF Hydroxylase Pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, Q. VHL and Hypoxia Signaling: Beyond HIF in Cancer. Biomedicines 2018, 6, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L. Hypoxia-Inducible Factor 1 (HIF-1) Pathway. Sci. STKE 2007, 2007, cm8. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.L.; Wu, C.; Xiong, Z.F.; Fang, X. Progress on Hypoxia-Inducible Factor-3: Its Structure, Gene Regulation and Biological Function (Review). Mol. Med. Rep. 2015, 12, 2411–2416. [Google Scholar] [CrossRef] [Green Version]
- Duan, C. Hypoxia-Inducible Factor 3 Biology: Complexities and Emerging Themes. Am. J. Physiol. Cell. Physiol. 2016, 310, C260–C269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, H.; Natsume, A.; Iwami, K.; Ohka, F.; Kuchimaru, T.; Kizaka-Kondoh, S.; Ito, K.; Saito, K.; Sugita, S.; Hoshino, T.; et al. A Hypoxia-Inducible Factor (HIF)-3α Splicing Variant, HIF-3α4 Impairs Angiogenesis in Hypervascular Malignant Meningiomas with Epigenetically Silenced HIF-3α4. Biochem. Biophys. Res. Commun. 2013, 433, 139–144. [Google Scholar] [CrossRef]
- Jing, X.; Yang, F.; Shao, C.; Wei, K.; Xie, M.; Shen, H.; Shu, Y. Role of Hypoxia in Cancer Therapy by Regulating the Tumor Microenvironment. Mol. Cancer 2019, 18, 157. [Google Scholar] [CrossRef] [Green Version]
- Manoochehri Khoshinani, H.; Afshar, S.; Najafi, R. Hypoxia: A Double-Edged Sword in Cancer Therapy. Cancer Investig. 2016, 34, 536–545. [Google Scholar] [CrossRef]
- Courtnay, R.; Ngo, D.C.; Malik, N.; Ververis, K.; Tortorella, S.M.; Karagiannis, T.C. Cancer Metabolism and the Warburg Effect: The Role of HIF-1 and PI3K. Mol. Biol. Rep. 2015, 42, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Nagao, A.; Kobayashi, M.; Koyasu, S.; Chow, C.C.T.; Harada, H. HIF-1-Dependent Reprogramming of Glucose Metabolic Pathway of Cancer Cells and its Therapeutic Significance. Int. J. Mol. Sci. 2019, 20, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, Z.; Lu, Y.; Qiu, S.; Fan, Z. Overcoming Cisplatin Resistance of Ovarian Cancer Cells by Targeting HIF-1-Regulated Cancer Metabolism. Cancer Lett. 2016, 373, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Zhan, Y.; Yuan, Z.; Qiu, Y.; Wang, H.; Fan, G.; Wang, J.; Li, W.; Cao, Y.; Shen, X.; et al. Hypoxia Induces Drug Resistance in Colorectal Cancer through the HIF-1alpha/miR-338-5p/IL-6 Feedback Loop. Mol. Ther. 2019, 27, 1810–1824. [Google Scholar] [CrossRef] [PubMed]
- Fallah, J.; Rini, B.I. HIF Inhibitors: Status of Current Clinical Development. Curr. Oncol. Rep. 2019, 21, 6. [Google Scholar] [CrossRef]
- Chen, W.; Hill, H.; Christie, A.; Kim, M.S.; Holloman, E.; Pavia-Jimenez, A.; Homayoun, F.; Ma, Y.; Patel, N.; Yell, P.; et al. Targeting Renal Cell Carcinoma with a HIF-2 Antagonist. Nature 2016, 539, 112–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhry, H.; Harris, A.L. Advances in Hypoxia-Inducible Factor Biology. Cell. Metab. 2018, 27, 281–298. [Google Scholar] [CrossRef]
- Li, H.; Jia, Y.; Wang, Y. Targeting HIF-1α Signaling Pathway for Gastric Cancer Treatment. Pharmazie 2019, 74, 3–7. [Google Scholar]
- Schanza, L.M.; Seles, M.; Stotz, M.; Fosselteder, J.; Hutterer, G.C.; Pichler, M.; Stiegelbauer, V. MicroRNAs Associated with Von Hippel-Lindau Pathway in Renal Cell Carcinoma: A Comprehensive Review. Int. J. Mol. Sci. 2017, 18, 2495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, H.; Vincent, K.; Pichler, M.; Fodde, R.; Berindan-Neagoe, I.; Slack, F.J.; Calin, G.A. Junk DNA and the Long Non-Coding RNA Twist in Cancer Genetics. Oncogene 2015, 34, 5003–5011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichler, M.; Rodriguez-Aguayo, C.; Nam, S.Y.; Dragomir, M.P.; Bayraktar, R.; Anfossi, S.; Knutsen, E.; Ivan, C.; Fuentes-Mattei, E.; Lee, S.K.; et al. Therapeutic Potential of FLANC, a Novel Primate-Specific Long Non-Coding RNA in Colorectal Cancer. Gut 2020. [Google Scholar] [CrossRef] [PubMed]
- Fosselteder, J.; Calin, G.A.; Pichler, M. Long Non-Coding RNA CCAT2 as a Therapeutic Target in Colorectal Cancer. Expert Opin. Ther. Targets 2018, 22, 973–976. [Google Scholar] [CrossRef]
- Cerk, S.; Schwarzenbacher, D.; Adiprasito, J.B.; Stotz, M.; Hutterer, G.C.; Gerger, A.; Ling, H.; Calin, G.A.; Pichler, M. Current Status of Long Non-Coding RNAs in Human Breast Cancer. Int. J. Mol. Sci. 2016, 17, 1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seles, M.; Hutterer, G.C.; Kiesslich, T.; Pummer, K.; Berindan-Neagoe, I.; Perakis, S.; Schwarzenbacher, D.; Stotz, M.; Gerger, A.; Pichler, M. Current Insights into Long Non-Coding RNAs in Renal Cell Carcinoma. Int. J. Mol. Sci. 2016, 17, 573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolle, M.A.; Pichler, M. The Role of Long Non-Coding RNAs in Osteosarcoma. Noncoding RNA 2018, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Barth, D.A.; Slaby, O.; Klec, C.; Juracek, J.; Drula, R.; Calin, G.A.; Pichler, M. Current Concepts of Non-Coding RNAs in the Pathogenesis of Non-Clear Cell Renal Cell Carcinoma. Cancers (Basel) 2019, 11, 1580. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the Classification of Long Non-Coding RNAs. RNA Biol. 2013, 10, 925–933. [Google Scholar] [CrossRef]
- Wang, K.; Chang, H. Molecular Mechanisms of Long Noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wang, W.; Liu, G.; Xie, S.; Li, Q.; Li, Y.; Lin, Z. Long Non-Coding RNA HOTTIP Promotes Hypoxia-Induced Epithelial-Mesenchymal Transition of Malignant Glioma by Regulating the miR-101/ZEB1 Axis. Biomed. Pharm. 2017, 95, 711–720. [Google Scholar] [CrossRef]
- Zhang, J.; Jin, H.Y.; Wu, Y.; Zheng, Z.C.; Guo, S.; Wang, Y.; Yang, D.; Meng, X.Y.; Xu, X.; Zhao, Y. Hypoxia-Induced LncRNA PCGEM1 Promotes Invasion and Metastasis of Gastric Cancer through Regulating SNAI1. Clin. Transl. Oncol. 2019, 21, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhao, X.; Zou, H.; Bai, R.; Yang, K.; Tian, Z. Hypoxia Promotes Gastric Cancer Malignancy Partly through the HIF-1alpha Dependent Transcriptional Activation of the Long Non-Coding RNA GAPLINC. Front. Physiol. 2016, 7, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Wang, C.; Ma, C.; Sun, W.; Li, H.; Cai, Z. Long Noncoding RNA ANRIL is Activated by Hypoxia-Inducible Factor-1alpha and Promotes Osteosarcoma Cell Invasion and Suppresses Cell Apoptosis upon Hypoxia. Cancer Cell. Int. 2016, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhry, H.; Harris, A.L.; McIntyre, A. The Tumour Hypoxia Induced Non-Coding Transcriptome. Mol. Asp. Med. 2016, 47, 35–53. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Li, L.; Xue, X.; Xie, H.; Shi, H.; Hu, Y. A lncRNA Coordinates with Ezh2 to Inhibit HIF-1alpha Transcription and Suppress Cancer Cell Adaption to Hypoxia. Oncogene 2020, 39, 1860–1874. [Google Scholar] [CrossRef]
- Wang, X.; Li, L.; Zhao, K.; Lin, Q.; Li, H.; Xue, X.; Ge, W.; He, H.; Liu, D.; Xie, H.; et al. A Novel LncRNA HITT Forms a Regulatory Loop with HIF-1alpha to Modulate Angiogenesis and Tumor Growth. Cell Death Differ. 2020, 27, 1431–1446. [Google Scholar] [CrossRef]
- Xiang, S.; Gu, H.; Jin, L.; Thorne, R.F.; Zhang, X.D.; Wu, M. LncRNA IDH1-AS1 Links the Functions of C-Myc and HIF1alpha Via IDH1 to Regulate the Warburg Effect. Proc. Natl. Acad. Sci. USA 2018, 115, E1465–E1474. [Google Scholar] [CrossRef] [Green Version]
- Shih, J.W.; Chiang, W.F.; Wu, A.T.H.; Wu, M.H.; Wang, L.Y.; Yu, Y.L.; Hung, Y.W.; Wang, W.C.; Chu, C.Y.; Hung, C.L.; et al. Long Noncoding RNA LncHIFCAR/MIR31HG is a HIF-1alpha Co-Activator Driving Oral Cancer Progression. Nat. Commun. 2017, 8, 15874. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Li, H. Long Non-Coding RNA GEHT1 Promoted the Proliferation of Ovarian Cancer Cells Via Modulating the Protein Stability of HIF1alpha. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Zhang, H.; Mei, Y.; Wu, M. Reciprocal Regulation of HIF-1alpha and lincRNA-p21 Modulates the Warburg Effect. Mol. Cell 2014, 53, 88–100. [Google Scholar]
- Wang, Y.; Chen, W.; Lian, J.; Zhang, H.; Yu, B.; Zhang, M.; Wei, F.; Wu, J.; Jiang, J.; Jia, Y.; et al. The lncRNA PVT1 Regulates Nasopharyngeal Carcinoma Cell Proliferation Via Activating the KAT2A Acetyltransferase and Stabilizing HIF-1alpha. Cell Death Differ. 2020, 27, 695–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Han, C.; Zhang, Y.; Liu, F. LncRNA PVT1 Regulate Expression of HIF1alpha Via Functioning as ceRNA for miR199a5p in Nonsmall Cell Lung Cancer Under Hypoxia. Mol. Med. Rep. 2018, 17, 1105–1110. [Google Scholar] [PubMed]
- Huang, T.; Liu, H.W.; Chen, J.Q.; Wang, S.H.; Hao, L.Q.; Liu, M.; Wang, B. The Long Noncoding RNA PVT1 Functions as a Competing Endogenous RNA by Sponging miR-186 in Gastric Cancer. Biomed. Pharmacother. 2017, 88, 302–308. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, K.; Cai, L. LINK-A lncRNA Functions in the Metastasis of Osteosarcoma by Upregulating HIF1alpha. Oncol. Lett. 2019, 17, 5005–5011. [Google Scholar] [PubMed] [Green Version]
- Zhang, H.; Yao, B.; Tang, S.; Chen, Y. LINK-A Long Non-Coding RNA (lncRNA) Participates in Metastasis of Ovarian Carcinoma and Upregulates Hypoxia-Inducible Factor 1 (HIF1alpha). Med. Sci. Monit. 2019, 25, 2221–2227. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Li, C.; Xing, Z.; Hu, Q.; Liang, K.; Han, L.; Wang, C.; Hawke, D.H.; Wang, S.; Zhang, Y.; et al. The LINK-A lncRNA Activates Normoxic HIF1alpha Signalling in Triple-Negative Breast Cancer. Nat. Cell Biol. 2016, 18, 213–224. [Google Scholar] [CrossRef]
- Chen, F.; Chen, J.; Yang, L.; Liu, J.; Zhang, X.; Zhang, Y.; Tu, Q.; Yin, D.; Lin, D.; Wong, P.P.; et al. Extracellular Vesicle-Packaged HIF-1alpha-Stabilizing lncRNA from Tumour-Associated Macrophages Regulates Aerobic Glycolysis of Breast Cancer Cells. Nat. Cell Biol. 2019, 21, 498–510. [Google Scholar] [CrossRef]
- Zhu, Y.; Tong, Y.; Wu, J.; Liu, Y.; Zhao, M. Knockdown of LncRNA GHET1 Suppresses Prostate Cancer Cell Proliferation by Inhibiting HIF-1alpha/Notch-1 Signaling Pathway Via KLF2. Biofactors 2019, 45, 364–373. [Google Scholar] [CrossRef]
- Wang, R.; Ma, Z.; Feng, L.; Yang, Y.; Tan, C.; Shi, Q.; Lian, M.; He, S.; Ma, H.; Fang, J. LncRNA MIR31HG Targets HIF1A and P21 to Facilitate Head and Neck Cancer Cell Proliferation and Tumorigenesis by Promoting Cell-Cycle Progression. Mol. Cancer 2018, 17, 162. [Google Scholar] [CrossRef]
- Wen, X.; Liu, X.; Mao, Y.P.; Yang, X.J.; Wang, Y.Q.; Zhang, P.P.; Lei, Y.; Hong, X.H.; He, Q.M.; Ma, J.; et al. Long Non-Coding RNA DANCR Stabilizes HIF-1alpha and Promotes Metastasis by Interacting with NF90/NF45 Complex in Nasopharyngeal Carcinoma. Theranostics 2018, 8, 5676–5689. [Google Scholar] [CrossRef]
- Jin, Y.; Xie, H.; Duan, L.; Zhao, D.; Ding, J.; Jiang, G. Long Non-Coding RNA CASC9 and HIF-1α Form A Positive Feedback Loop to Facilitate Cell Proliferation and Metastasis in Lung Cancer. OncoTargets Ther. 2019, 12, 9017–9027. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Li, G.; Liu, W. The Long Noncoding RNA Cancer Susceptibility Candidate 9 Promotes Nasopharyngeal Carcinogenesis Via Stabilizing HIF1alpha. DNA Cell Biol. 2017, 36, 394–400. [Google Scholar] [CrossRef]
- Luo, F.; Liu, X.; Ling, M.; Lu, L.; Shi, L.; Lu, X.; Li, J.; Zhang, A.; Liu, Q. The lncRNA MALAT1, Acting through HIF-1alpha Stabilization, Enhances Arsenite-Induced Glycolysis in Human Hepatic L-02 Cells. Biochim. Biophys. Acta 2016, 1862, 1685–1695. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Xu, F.Y.; Zheng, H.; Cheng, P.; Chen, Q.Y.; Ye, Z.; Zhong, J.X.; Deng, S.J.; Liu, M.L.; Huang, K.; et al. LncRNA-MTA2TR Functions as a Promoter in Pancreatic Cancer Via Driving Deacetylation-Dependent Accumulation of HIF-1alpha. Theranostics 2019, 9, 5298–5314. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Cao, S.; Wang, Y.; Hu, Y.; Liu, H.; Li, J.; Chen, J.; Li, P.; Liu, J.; Wang, Q.; et al. Long Non-Coding RNA UBE2CP3 Enhances HCC Cell Secretion of VEGFA and Promotes Angiogenesis by Activating ERK1/2/HIF-1alpha/VEGFA Signalling in Hepatocellular Carcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, F.; Liao, Y.; Yuan, L.; Zhang, B. LncRNA AWPPH Promotes the Invasion and Migration of Glioma Cells through the Upregulation of HIF1alpha. Oncol. Lett. 2019, 18, 6781–6786. [Google Scholar] [PubMed] [Green Version]
- Yang, F.; Huo, X.S.; Yuan, S.X.; Zhang, L.; Zhou, W.P.; Wang, F.; Sun, S.H. Repression of the Long Noncoding RNA-LET by Histone Deacetylase 3 Contributes to Hypoxia-Mediated Metastasis. Mol. Cell 2013, 49, 1083–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.W.; Chen, Y.F.; Li, J.; Huo, Y.M.; Liu, D.J.; Hua, R.; Zhang, J.F.; Liu, W.; Yang, J.Y.; Fu, X.L.; et al. A Novel Long Non-Coding RNA ENST00000480739 Suppresses Tumour Cell Invasion by Regulating OS-9 and HIF-1alpha in Pancreatic Ductal Adenocarcinoma. Br. J. Cancer 2014, 111, 2131–2141. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yuan, W.; Song, J.; Wang, S.; Gu, X. LncRNA CPS1-IT1 Suppresses EMT and Metastasis of Colorectal Cancer by Inhibiting Hypoxia-Induced Autophagy through Inactivation of HIF-1alpha. Biochimie 2018, 144, 21–27. [Google Scholar] [CrossRef]
- Wang, T.H.; Wu, C.H.; Yeh, C.T.; Su, S.C.; Hsia, S.M.; Liang, K.H.; Chen, C.C.; Hsueh, C.; Chen, C.Y. Melatonin Suppresses Hepatocellular Carcinoma Progression Via lncRNA-CPS1-IT-Mediated HIF-1alpha Inactivation. Oncotarget 2017, 8, 82280–82293. [Google Scholar] [CrossRef]
- Wang, T.H.; Yu, C.C.; Lin, Y.S.; Chen, T.C.; Yeh, C.T.; Liang, K.H.; Shieh, T.M.; Chen, C.Y.; Hsueh, C. Long Noncoding RNA CPS1-IT1 Suppresses the Metastasis of Hepatocellular Carcinoma by Regulating HIF-1alpha Activity and Inhibiting Epithelial-Mesenchymal Transition. Oncotarget 2016, 7, 43588–43603. [Google Scholar] [CrossRef]
- Zhou, C.; Huang, C.; Wang, J.; Huang, H.; Li, J.; Xie, Q.; Liu, Y.; Zhu, J.; Li, Y.; Zhang, D.; et al. LncRNA MEG3 Downregulation Mediated by DNMT3b Contributes to Nickel Malignant Transformation of Human Bronchial Epithelial Cells Via Modulating PHLPP1 Transcription and HIF-1α Translation. Oncogene 2017, 36, 3878–3889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.J.; Kim, I.; Lee, J.E.; Park, J.W. PIN1 Transcript Variant 2 Acts as a Long Non-Coding RNA that Controls the HIF-1-Driven Hypoxic Response. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.J.; Young, M.; Jang, K.; Sauer, S.; Neang, V.E.; King, A.T.; Girke, T.; Martinez, E. HOTAIRM1 lncRNA is Downregulated in Clear Cell Renal Cell Carcinoma and Inhibits the Hypoxia Pathway. Cancer Lett. 2020, 472, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Wang, S.; Chu, X.; Han, D. LncRNA HIF2PUT Inhibited Osteosarcoma Stem Cells Proliferation, Migration and Invasion by Regulating HIF2 Expression. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1342–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Li, J.; Geng, P.; Li, Y.; Chen, H.; Zhu, Y. Knockdown of a HIF-2alpha Promoter Upstream Long Noncoding RNA Impairs Colorectal Cancer Stem Cell Properties in Vitro through HIF-2alpha Downregulation. OncoTargets Ther. 2015, 8, 3467–3474. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yao, J.; Meng, H.; Yu, Z.; Wang, Z.; Yuan, X.; Chen, H.; Wang, A. A Novel Long Non-Coding RNA, Hypoxia-Inducible Factor-2alpha Promoter Upstream Transcript, Functions as an Inhibitor of Osteosarcoma Stem Cells in Vitro. Mol. Med. Rep. 2015, 11, 2534–2540. [Google Scholar] [CrossRef] [Green Version]
- Zhai, W.; Sun, Y.; Jiang, M.; Wang, M.; Gasiewicz, T.A.; Zheng, J.; Chang, C. Differential Regulation of LncRNA-SARCC Suppresses VHL-Mutant RCC Cell Proliferation Yet Promotes VHL-Normal RCC Cell Proliferation Via Modulating Androgen Receptor/HIF-2alpha/C-MYC Axis Under Hypoxia. Oncogene 2016, 35, 4866–4880. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Sun, B.; Li, H.; Xu, Y.; Liu, Y.; Liu, X.; Lu, L.; Li, J.; Wang, Q.; Wei, S.; et al. A MALAT1/HIF-A Feedback Loop Contributes to Arsenite Carcinogenesis. Oncotarget 2016, 7, 5769–5787. [Google Scholar] [CrossRef] [Green Version]
- Derderian, C.; Orunmuyi, A.T.; Olapade-Olaopa, E.O.; Ogunwobi, O.O. PVT1 Signaling is a Mediator of Cancer Progression. Front. Oncol. 2019, 9, 502. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Zheng, G.; Gao, C. LncRNA PVT1: A Novel Therapeutic Target for Cancers. Clin. Lab. 2018, 64, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Gao, H.; Liu, K.; Gao, B.; Ren, H.; Li, Z.; Liu, F. The lncRNA ZEB2-AS1 is Upregulated in Gastric Cancer and Affects Cell Proliferation and Invasion Via miR-143-5p/HIF-1α Axis. OncoTargets Ther. 2019, 12, 657–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.G.; Cao, M.Z.; Zhang, J.; Li, X.Y.; Sun, Q.L. LncRNA XIST Modulates HIF-1A/AXL Signaling Pathway by Inhibiting miR-93-5p in Colorectal Cancer. Mol. Genet. Genom. Med. 2020, e1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Hu, L.; Li, S.; Shen, J.; Wang, D.; Xu, R.; Yang, H. Long Non-Coding RNA Taurine Upregulated Gene 1 Promotes Osteosarcoma Cell Metastasis by Mediating HIF-1alpha Via miR-143-5p. Cell. Death Dis. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Peng, X.; Yan, J.; Cheng, F. LncRNA TMPO-AS1 Up-Regulates the Expression of HIF-1alpha and Promotes the Malignant Phenotypes of Retinoblastoma Cells Via Sponging miR-199a-5p. Pathol. Res. Pract. 2020, 216, 152853. [Google Scholar] [CrossRef]
- Du, F.; Guo, T.; Cao, C. Silencing of Long Noncoding RNA SNHG6 Inhibits Esophageal Squamous Cell Carcinoma Progression Via miR-186-5p/HIF1alpha Axis. Dig. Dis. Sci. 2019, 18, 1–9. [Google Scholar] [CrossRef]
- Takahashi, K.; Yan, I.K.; Haga, H.; Patel, T. Modulation of Hypoxia-Signaling Pathways by Extracellular Linc-RoR. J. Cell. Sci. 2014, 127, 1585–1594. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Wang, Z.; Wang, S.; Weng, M.; Zhou, D.; Li, C.; Wang, J.; Chen, E.; Quan, Z. Long Non-Coding RNA LINC00152 Promotes Gallbladder Cancer Metastasis and Epithelial-Mesenchymal Transition by Regulating HIF-1alpha Via miR-138. Open Biol. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Ou, Z.; Zhang, M.; Ji, L.; Luo, Z.; Han, T.; Lu, Y.; Li, Y. Long Noncoding RNA FEZF1-AS1 Predicts Poor Prognosis and Modulates Pancreatic Cancer Cell Proliferation and Invasion through miR-142/HIF-1α and miR-133a/EGFR upon Hypoxia/Normoxia. J. Cell. Physiol. 2019, 234, 15407–15419. [Google Scholar] [CrossRef]
- Yang, B.; Jia, L.; Ren, H.; Jin, C.; Ren, Q.; Zhang, H.; Hu, D.; Zhang, H.; Hu, L.; Xie, T. LncRNA DLX6-AS1 Increases the Expression of HIF-1alpha and Promotes the Malignant Phenotypes of Nasopharyngeal Carcinoma Cells Via Targeting MiR-199a-5p. Mol. Genet. Genom. Med. 2020, 8, e1017. [Google Scholar]
- Wang, Y.; Huang, Y.; Liu, H.; Su, D.; Luo, F.; Zhou, F. Long Noncoding RNA CDKN2B-AS1 Interacts with miR-411-3p to Regulate Ovarian Cancer in Vitro and in Vivo through HIF-1a/VEGF/P38 Pathway. Biochem. Biophys. Res. Commun. 2019, 514, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, Y.; Liu, A.; Tang, X. Long Non-Coding RNA UCA1 Enhances Tamoxifen Resistance in Breast Cancer Cells through a miR-18a-HIF1alpha Feedback Regulatory Loop. Tumour Biol. 2016, 37, 14733–14743. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Lee, S.; Cao, P. The Inhibitive Effect of Sh-HIF1A-AS2 on the Proliferation, Invasion, and Pathological Damage of Breast Cancer Via Targeting miR-548c-3p through Regulating HIF-1alpha/VEGF Pathway in Vitro and Vivo. OncoTargets Ther. 2019, 12, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Hong, Q.; Li, O.; Zheng, W.; Xiao, W.Z.; Zhang, L.; Wu, D.; Cai, G.Y.; He, J.C.; Chen, X.M. LncRNA HOTAIR Regulates HIF-1alpha/AXL Signaling through Inhibition of miR-217 in Renal Cell Carcinoma. Cell. Death Dis. 2017, 8, e2772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Ye, L.; Jiang, C.; Bai, J.; Chi, Y.; Zhang, H. Long Noncoding RNA HOTAIR, a Hypoxia-Inducible Factor-1α activated Driver of Malignancy, Enhances Hypoxic Cancer Cell Proliferation, Migration, and Invasion in Non-Small Cell Lung Cancer. Tumor Biol. 2015, 36, 9179–9188. [Google Scholar] [CrossRef]
- Li, N.; Meng, D.D.; Gao, L.; Xu, Y.; Liu, P.J.; Tian, Y.W.; Yi, Z.Y.; Zhang, Y.; Tie, X.J.; Xu, Z.Q. Overexpression of HOTAIR Leads to Radioresistance of Human Cervical Cancer Via Promoting HIF-1α expression. Radiat. Oncol. 2018, 13, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.; Fu, Q.; Jing, C.; Zhang, X.; Qin, T.; Pan, Y. LncRNA HOTAIR Knockdown Inhibits Glycolysis by Regulating miR-130a-3p/HIF1A in Hepatocellular Carcinoma Under Hypoxia. Biomed. Pharm. 2020, 125, 109703. [Google Scholar] [CrossRef]
- Peng, F.; Wang, J.; Fan, W.; Meng, Y.; Li, M.; Li, T.; Cui, B.; Wang, H.; Zhao, Y.; An, F.; et al. Glycolysis Gatekeeper PDK1 Reprograms Breast Cancer Stem Cells Under Hypoxia. Oncogene 2018, 37, 1062–1074. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Jin, Y.; Lyu, X.; Fan, L.; Wu, F. Long Noncoding RNA H19 Regulates HIF-1α/AXL Signaling through Inhibiting miR-20b-5p in Endometrial Cancer. Cell Cycle 2019, 18, 2454–2464. [Google Scholar] [CrossRef]
- Corrado, C.; Costa, V.; Giavaresi, G.; Calabrese, A.; Conigliaro, A.; Alessandro, R. Long Non Coding RNA H19: A New Player in Hypoxia-Induced Multiple Myeloma Cell Dissemination. Int. J. Mol. Sci. 2019, 20, 801. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.; Zhao, L. lncRNA Nuclear-Enriched Abundant Transcript 1 Promotes Cell Proliferation and Invasion by Targeting miR-186-5p/HIF-1alpha in Osteosarcoma. J. Cell. Biochem. 2019, 120, 6502–6514. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Esmaeili, M.; Taheri, M. H19 lncRNA: Roles in Tumorigenesis. Biomed. Pharm. 2020, 123, 109774. [Google Scholar] [CrossRef]
- Yoshimura, H.; Matsuda, Y.; Yamamoto, M.; Kamiya, S.; Ishiwata, T. Expression and Role of Long Non-Coding RNA H19 in Carcinogenesis. Front. Biosci. (Landmark Ed.) 2018, 23, 614–625. [Google Scholar]
- Matouk, I.J.; Mezan, S.; Mizrahi, A.; Ohana, P.; Abu-Lail, R.; Fellig, Y.; Degroot, N.; Galun, E.; Hochberg, A. The Oncofetal H19 RNA Connection: Hypoxia, p53 and Cancer. Biochim. Biophys. Acta 2010, 1803, 443–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Hu, Q.; Nie, E.; Yu, T.; Wu, Y.; Zhi, T.; Jiang, K.; Shen, F.; Wang, Y.; Zhang, J.; et al. Hypoxia Induces H19 Expression through Direct and Indirect Hif-1α Activity, Promoting Oncogenic Effects in Glioblastoma. Sci. Rep. 2017, 7, 45029. [Google Scholar] [CrossRef] [PubMed]
- Papandreou, I.; Cairns, R.A.; Fontana, L.; Lim, A.L.; Denko, N.C. HIF-1 Mediates Adaptation to Hypoxia by Actively Downregulating Mitochondrial Oxygen Consumption. Cell. Metab. 2006, 3, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Dico, A.; Costa, V.; Martelli, C.; Diceglie, C.; Rajata, F.; Rizzo, A.; Mancone, C.; Tripodi, M.; Ottobrini, L.; Alessandro, R.; et al. MiR675-5p Acts on HIF-1alpha to Sustain Hypoxic Responses: A New Therapeutic Strategy for Glioma. Theranostics 2016, 6, 1105–1118. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Alsager, S.; Zhuo, Y.; Shan, B. HOX Transcript Antisense RNA (HOTAIR) in Cancer. Cancer Lett. 2019, 454, 90–97. [Google Scholar] [CrossRef]
- Li, T.; Xiao, Y.; Huang, T. HIF1alphainduced Upregulation of lncRNA UCA1 Promotes Cell Growth in Osteosarcoma by Inactivating the PTEN/AKT Signaling Pathway. Oncol. Rep. 2018, 39, 1072–1080. [Google Scholar]
- Xue, M.; Li, X.; Li, Z.; Chen, W. Urothelial Carcinoma Associated 1 is a Hypoxia-Inducible Factor-1alpha-Targeted Long Noncoding RNA that Enhances Hypoxic Bladder Cancer Cell Proliferation, Migration, and Invasion. Tumour Biol. 2014, 35, 6901–6912. [Google Scholar] [CrossRef]
- Yang, J.; Li, L.; Hong, S.; Zhou, Z.; Fan, W. LINK-A lncRNA Activates HIF1alpha Signaling and Inhibits Podocyte Cell Apoptosis in Diabetic Nephropathy. Exp. Ther. Med. 2019, 18, 119–124. [Google Scholar] [PubMed]
- Ye, Y.; Peng, Y.; Li, Y.; Liu, C.; Xu, Y.; Li, W. Effect of lincRNA-p21 Targeting HIF-1alpha on Biological Functions of Liver Cancer Cells. Oncol. Lett. 2019, 17, 4964–4968. [Google Scholar]
- Shen, Y.; Liu, Y.; Sun, T.; Yang, W. LincRNA-p21 Knockdown Enhances Radiosensitivity of Hypoxic Tumor Cells by Reducing Autophagy through HIF-1/Akt/mTOR/P70S6K Pathway. Exp. Cell Res. 2017, 358, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Maolake, A.; Izumi, K.; Shigehara, K.; Natsagdorj, A.; Iwamoto, H.; Kadomoto, S.; Takezawa, Y.; Machioka, K.; Narimoto, K.; Namiki, M.; et al. Tumor-Associated Macrophages Promote Prostate Cancer Migration through Activation of the CCL22-CCR4 Axis. Oncotarget 2017, 8, 9739–9751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Palma, M.; Biziato, D.; Petrova, T.V. Microenvironmental Regulation of Tumour Angiogenesis. Nat. Rev. Cancer 2017, 17, 457–474. [Google Scholar] [CrossRef] [PubMed]
- Goswami, S.; Sahai, E.; Wyckoff, J.B.; Cammer, M.; Cox, D.; Pixley, F.J.; Stanley, E.R.; Segall, J.E.; Condeelis, J.S. Macrophages Promote the Invasion of Breast Carcinoma Cells Via a Colony-Stimulating Factor-1/Epidermal Growth Factor Paracrine Loop. Cancer Res. 2005, 65, 5278–5283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.G.; Cao, B.; Zhang, L.X.; Song, N.; Li, H.; Zhao, W.Z.; Li, Y.S.; Ma, S.M.; Yin, D.J. KLF2 Inhibits Cell Growth Via Regulating HIF-1alpha/Notch-1 Signal Pathway in Human Colorectal Cancer HCT116 Cells. Oncol. Rep. 2017, 38, 584–590. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; He, Y.; Tang, S.; Huang, S. LncRNA GHET1 Predicts Poor Prognosis in Hepatocellular Carcinoma and Promotes Cell Proliferation by Silencing KLF2. J. Cell. Physiol. 2018, 233, 4726–4734. [Google Scholar] [CrossRef]
- Augoff, K.; McCue, B.; Plow, E.F.; Sossey-Alaoui, K. miR-31 and its Host Gene lncRNA LOC554202 are Regulated by Promoter Hypermethylation in Triple-Negative Breast Cancer. Mol. Cancer 2012, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Nie, F.Q.; Ma, S.; Xie, M.; Liu, Y.W.; De, W.; Liu, X.H. Decreased Long Noncoding RNA MIR31HG is Correlated with Poor Prognosis and Contributes to Cell Proliferation in Gastric Cancer. Tumour Biol. 2016, 37, 7693–7701. [Google Scholar] [CrossRef]
- Wang, B.; Jiang, H.; Wang, L.; Chen, X.; Wu, K.; Zhang, S.; Ma, S.; Xia, B. Increased MIR31HG lncRNA Expression Increases Gefitinib Resistance in Non-Small Cell Lung Cancer Cell Lines through the EGFR/PI3K/AKT Signaling Pathway. Oncol. Lett. 2017, 13, 3494–3500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Liu, P.; Zhang, J.; Peng, X.; Lu, Z.; Yu, S.; Meng, Y.; Tong, W.M.; Chen, J. Long Noncoding RNA MIR31HG Exhibits Oncogenic Property in Pancreatic Ductal Adenocarcinoma and is Negatively Regulated by miR-193b. Oncogene 2016, 35, 3647–3657. [Google Scholar] [CrossRef] [PubMed]
- Dandan, W.; Jianliang, C.; Haiyan, H.; Hang, M.; Xuedong, L. Long Noncoding RNA MIR31HG is Activated by SP1 and Promotes Cell Migration and Invasion by Sponging miR-214 in NSCLC. Gene 2019, 692, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Zhang, X.; Wang, X.; Li, J. MIR31HG Promotes Cell Proliferation and Invasion by Activating the Wnt/Beta-Catenin Signaling Pathway in Non-Small Cell Lung Cancer. Oncol. Lett. 2019, 17, 221–229. [Google Scholar]
- Castella, S.; Bernard, R.; Corno, M.; Fradin, A.; Larcher, J.C. Ilf3 and NF90 Functions in RNA Biology. Wiley Interdiscip. Rev. RNA 2015, 6, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Friedrich, S.; Golbik, R.P.; Behrens, S.E. NF90-NF45 is a Selective RNA Chaperone that Rearranges Viral and Cellular Riboswitches: Biochemical Analysis of a Virus Host Factor Activity. Nucleic Acids Res. 2017, 45, 12441–12454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Ma, L. New Insights into Long Non-Coding RNA MALAT1 in Cancer and Metastasis. Cancers (Basel) 2019, 11, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Tang, J.; Huang, X.; Zhang, T.; Feng, X. Hypoxia Exposure Upregulates MALAT-1 and Regulates the Transcriptional Activity of PTB-Associated Splicing Factor in A549 Lung Adenocarcinoma Cells. Oncol. Lett. 2018, 16, 294–300. [Google Scholar] [CrossRef]
- Ikeda, S.; Kitadate, A.; Abe, F.; Takahashi, N.; Tagawa, H. Hypoxia-Inducible KDM3A Addiction in Multiple Myeloma. Blood Adv. 2018, 2, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Deng, S.; He, C.; Liu, M.; Chen, H.; Zeng, Z.; Zhong, J.; Ye, Z.; Deng, S.; Wu, H.; et al. Reciprocal Loop of Hypoxia-Inducible Factor-1α (HIF-1α) and Metastasis-Associated Protein 2 (MTA2) Contributes to the Progression of Pancreatic Carcinoma by Suppressing E-Cadherin Transcription. J. Pathol. 2018, 245, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.N.; Qu, H.J.; Gong, W.J.; Xiang, J.Y.; Yang, M.M.; Zhang, W. LncRNA AWPPH and miRNA-21 Regulates Cancer Cell Proliferation and Chemosensitivity in Triple-Negative Breast Cancer by Interacting with each Other. J. Cell. Biochem. 2019, 120, 14860–14866. [Google Scholar] [CrossRef] [PubMed]
- Huo, Y.; Li, A.; Wang, Z. LncRNA AWPPH Participates in the Metastasis of Non-Small Cell Lung Cancer by Upregulating TGF-Beta1 Expression. Oncol. Lett. 2019, 18, 4246–4252. [Google Scholar] [PubMed] [Green Version]
- Li, C.; Wang, F.; Wei, B.; Wang, L.; Kong, D. LncRNA AWPPH Promotes Osteosarcoma Progression Via Activation of Wnt/Beta-Catenin Pathway through Modulating miR-93-3p/FZD7 Axis. Biochem. Biophys. Res. Commun. 2019, 514, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Weidle, U.H.; Birzele, F.; Kollmorgen, G.; Ruger, R. Long Non-Coding RNAs and their Role in Metastasis. Cancer. Genom. Proteom. 2017, 14, 143–160. [Google Scholar]
- Zhuang, J.; Shen, L.; Yang, L.; Huang, X.; Lu, Q.; Cui, Y.; Zheng, X.; Zhao, X.; Zhang, D.; Huang, R.; et al. TGFbeta1 Promotes Gemcitabine Resistance through Regulating the LncRNA-LET/NF90/miR-145 Signaling Axis in Bladder Cancer. Theranostics 2017, 7, 3053–3067. [Google Scholar] [CrossRef]
- Mao, Z.; Li, H.; Du, B.; Cui, K.; Xing, Y.; Zhao, X.; Zai, S. LncRNA DANCR Promotes Migration and Invasion through Suppression of lncRNA-LET in Gastric Cancer Cells. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [Green Version]
- Piunti, A.; Shilatifard, A. Epigenetic Balance of Gene Expression by Polycomb and COMPASS Families. Science 2016, 352, aad9780. [Google Scholar] [CrossRef] [Green Version]
- Newton, A.C.; Trotman, L.C. Turning off AKT: PHLPP as a Drug Target. Annu. Rev. Pharm. Toxicol. 2014, 54, 537–558. [Google Scholar] [CrossRef] [Green Version]
- Dang, C.V. MYC on the Path to Cancer. Cell 2012, 149, 22–35. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, E.D.; Selak, M.A.; Tennant, D.A.; Payne, L.J.; Crosby, S.; Frederiksen, C.M.; Watson, D.G.; Gottlieb, E. Cell-Permeating Alpha-Ketoglutarate Derivatives Alleviate Pseudohypoxia in Succinate Dehydrogenase-Deficient Cells. Mol. Cell. Biol. 2007, 27, 3282–3289. [Google Scholar] [CrossRef] [Green Version]
- Lonati, E.; Brambilla, A.; Milani, C.; Masserini, M.; Palestini, P.; Bulbarelli, A. Pin1, a New Player in the Fate of HIF-1alpha Degradation: An Hypothetical Mechanism Inside Vascular Damage as Alzheimer’s Disease Risk Factor. Front. Cell. Neurosci. 2014, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalouli, M.; Dery, M.A.; Lafleur, V.N.; Lamalice, L.; Zhou, X.Z.; Lu, K.P.; Richard, D.E. The Prolyl Isomerase Pin1 Regulates Hypoxia-Inducible Transcription Factor (HIF) Activity. Cell. Signal. 2014, 26, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Han, H.J.; Kwon, N.; Choi, M.A.; Jung, K.O.; Piao, J.Y.; Ngo, H.K.; Kim, S.J.; Kim, D.H.; Chung, J.K.; Cha, Y.N.; et al. Peptidyl Prolyl Isomerase PIN1 Directly Binds to and Stabilizes Hypoxia-Inducible Factor-1alpha. PLoS ONE 2016, 11, e0147038. [Google Scholar]
- Li, W.; He, X.; Xue, R.; Zhang, Y.; Zhang, X.; Lu, J.; Zhang, Z.; Xue, L. Combined Over-Expression of the Hypoxia-Inducible Factor 2alpha Gene and its Long Non-Coding RNA Predicts Unfavorable Prognosis of Patients with Osteosarcoma. Pathol. Res. Pract. 2016, 212, 861–866. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, Y.; Rao, Q.; Xu, H.; Li, L.; Chang, C. Androgen Receptor (AR) Suppresses miRNA-145 to Promote Renal Cell Carcinoma (RCC) Progression Independent of VHL Status. Oncotarget 2015, 6, 31203–31215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, D.; Li, L.; Zhu, G.; Liang, L.; Guan, Z.; Chang, L.; Chen, Y.; Yeh, S.; Chang, C. ASC-J9 Suppresses Renal Cell Carcinoma Progression by Targeting an Androgen Receptor-Dependent HIF2alpha/VEGF Signaling Pathway. Cancer Res. 2014, 74, 4420–4430. [Google Scholar] [CrossRef] [Green Version]
- Yuan, P.; Cao, W.; Zang, Q.; Li, G.; Guo, X.; Fan, J. The HIF-2alpha-MALAT1-miR-216b Axis Regulates Multi-Drug Resistance of Hepatocellular Carcinoma Cells Via Modulating Autophagy. Biochem. Biophys. Res. Commun. 2016, 478, 1067–1073. [Google Scholar] [CrossRef]
- Klec, C.; Prinz, F.; Pichler, M. Involvement of the Long Noncoding RNA NEAT1 in Carcinogenesis. Mol. Oncol. 2019, 13, 46–60. [Google Scholar] [CrossRef] [Green Version]
- Dong, P.; Xiong, Y.; Yue, J.; Hanley, S.J.B.; Kobayashi, N.; Todo, Y.; Watari, H. Long Non-Coding RNA NEAT1: A Novel Target for Diagnosis and Therapy in Human Tumors. Front. Genet. 2018, 9, 471. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Zhang, Y.; Liu, Y.; Fang, L.; Li, L.; Sun, J.; Pan, Z.; Xin, W.; Huang, P. HIF-2α Activated lncRNA NEAT1 Promotes Hepatocellular Carcinoma Cell Invasion and Metastasis by Affecting the Epithelial-Mesenchymal Transition. J. Cell. Biochem. 2018, 119, 3247–3256. [Google Scholar] [CrossRef]
- Choudhry, H.; Albukhari, A.; Morotti, M.; Haider, S.; Moralli, D.; Smythies, J.; Schodel, J.; Green, C.M.; Camps, C.; Buffa, F.; et al. Tumor Hypoxia Induces Nuclear Paraspeckle Formation through HIF-2alpha Dependent Transcriptional Activation of NEAT1 Leading to Cancer Cell Survival. Oncogene 2015, 34, 4546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Zhao, Y.; Li, X.; Tao, Z.; Hou, M.; Ma, H. Overexpression of HIF-2alpha-Dependent NEAT1 Promotes the Progression of Non-Small Cell Lung Cancer through miR-101-3p/SOX9/Wnt/Beta-Catenin Signal Pathway. Cell. Physiol. Biochem. 2019, 52, 368–381. [Google Scholar] [PubMed] [Green Version]
- Zhang, P.; Cao, L.; Zhou, R.; Yang, X.; Wu, M. The lncRNA Neat1 Promotes Activation of Inflammasomes in Macrophages. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]


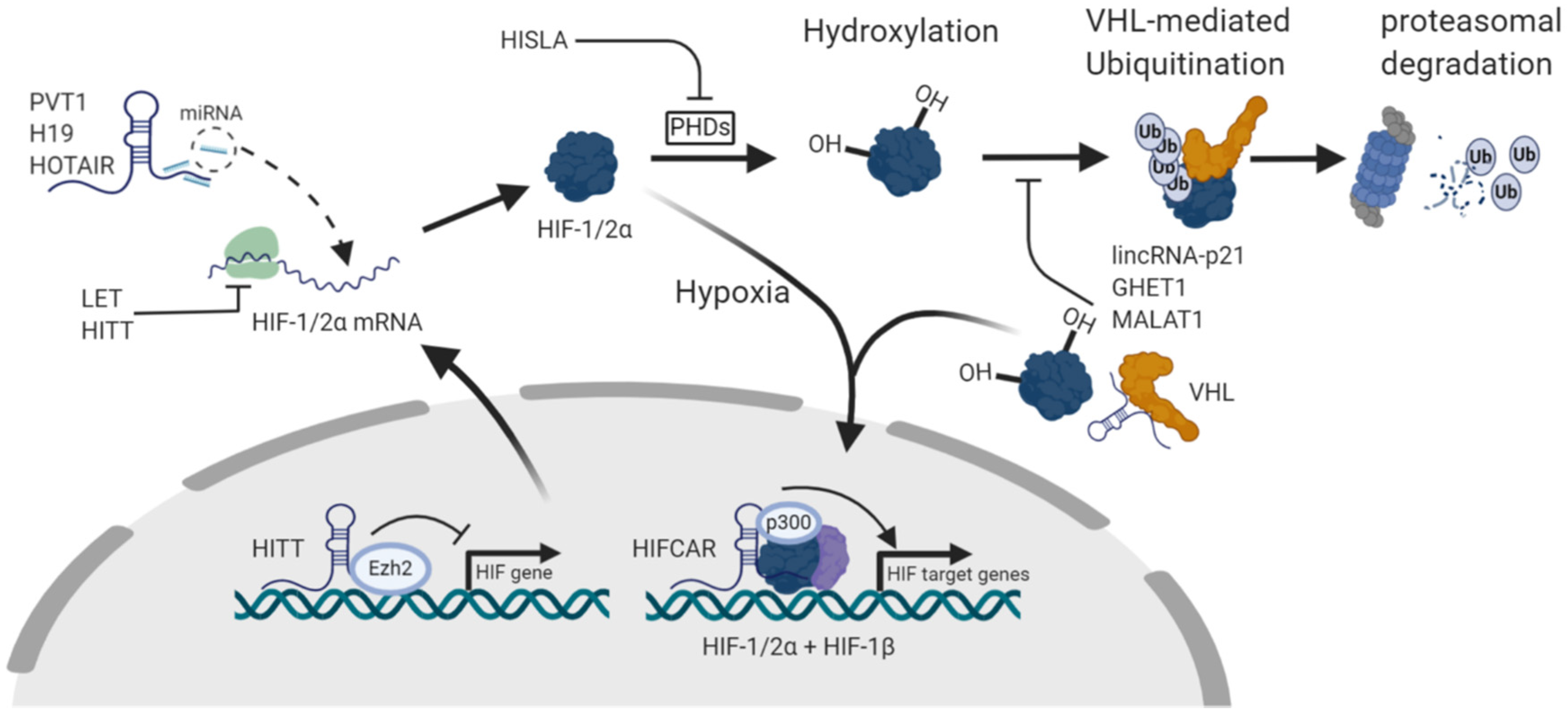{kind=link}
| lncRNA | Impact on HIF | Role in HIF Regulation | References | |
|---|---|---|---|---|
| HIF-1α | ||||
| PVT1 | ↑ | increases HIF-1α expression and stability | [41,42,43] | |
| LINK-A | ↑ | increases HIF-1α stability and activation | [44,45,46] | |
| lincRNA-p21 | ↑ | increases HIF-1α protein stability | [40] | |
| HISLA | ↑ | increases HIF-1α protein stability | [47] | |
| GHET1 | ↑ | increases HIF-1α expression and stability | [39,48] | |
| MIR31HG/HIFCAR | ↑ | enhances HIF-1α activation | [38,49] | |
| DANCR | ↑ | stabilizes HIF-1α mRNA | [50] | |
| CASC9 | ↑ | increases HIF-1α protein stability | [51,52] | |
| MALAT1 | ↑ | increases HIF-1α protein stability | [53] | |
| MTA2TR | ↑ | increases HIF-1α protein stability | [54] | |
| UBE2CP3 | ↑ | no specific mechanism defined | [55] | |
| AWPPH | ↑ | no specific mechanism defined | [56] | |
| LET | ↓ | decreases HIF-1α mRNA stability | [57] | |
| ENST00000480739 | ↓ | decreases HIF-1α protein stability | [58] | |
| CPS1-IT1 | ↓ | decreases HIF-1α activation | [59,60,61] | |
| HITT | ↓ | inhibits HIF-1α transcription and translation | [35,36] | |
| MEG3 | ↓ | Increases HIF-1α expression | [62] | |
| IDH1-AS1 | ↓ | decreases HIF-1α protein stability | [37] | |
| PIN1-v2 | ↓ | inhibits HIF-1α transcription | [63] | |
| HOTAIRM1 | ↓ | post-transcriptionally inhibits HIF-1α expression | [64] | |
| HIF-2α | ||||
| HIF2PUT | ↑ | no specific mechanism defined, increases expression | [65,66,67] | |
| SARCC | ↓ | decreases HIF-2α transcription and translation | [68] | |
| MALAT1 | ↑ | increases HIF-2α protein stability | [69] | |
| lincRNA-p21 | ↑ | increases HIF-2α protein stability | [40] |
| lncRNA | Cancer | Expression Pattern | Impact on HIF Expression | Pathway | Reference | |
|---|---|---|---|---|---|---|
| HIF-1α | ||||||
| PVT1 | Gastric cancer | ↑ | increase | PVT1/miR-186/HIF-1α | [43] | |
| Non-small cell lung cancer | ↑ | increase | PVT1/miR-199a-5p/HIF-1α | [42] | ||
| H19 | Endometrial cancer | ↑ | increase | H19/miR-20b-5p/HIF-1α/AXL | [89] | |
| Breast cancer | ↑ | increase | H19/miR-let-7/HIF-1α/PDK1 | [88] | ||
| HOTAIR | Hepatic cancer | ↑ | increase | HOTAIR/miR-130a-3p/HIF-1α | [87] | |
| Renal cell carcinoma | ↑ | increase | HOTAIR/miR-217/HIF-1α/AXL | [84] | ||
| Cervical cancer | ↑ | increase | HOTAIR/miR-127/HIF-1α | [86] | ||
| HIF1A-AS2 | Breast cancer | ↑ | increase | HIF1A-AS2/miR-548c-3p/HIF-1α/VEGF | [83] | |
| UCA1 | Breast cancer | ↑ | increase | UCA1/miR-18a/HIF-1α | [82] | |
| CDKN2B-AS1 | Ovarian cancer | ↑ | increase | CDKN2B-AS1/miR-411-3p/HIF-1α/VEGF | [81] | |
| DLX6-AS1 | Nasopharyngeal carcinoma | ↑ | increase | DLX6-AS1/miR-199a-5p/HIF-1α | [80] | |
| FEZF1-AS1 | Pancreatic cancer | ↑ | increase | FEZF1-AS1/miR-142/HIF-1α | [79] | |
| LINC00512 | Gallbladder carcinoma | ↑ | increase | LINC00512/miR-138/HIF-1α | [78] | |
| RoR | Hepatic cancer | ↑ | increase | RoR/miR-145/HIF-1α | [77] | |
| SNHG6 | Esophageal squamous cell carcinoma | ↑ | increase | SNHG6/miR-186-5p/HIF-1α | [76] | |
| TMPO-AS1 | Retinoblastoma | ↑ | increase | TMPO-AS1/miR-199a-5p/HIF-1α | [75] | |
| TUG1 | Osteosarcoma | ↑ | increase | TUG1/miR-143-5p/HIF-1α | [74] | |
| XIST | Colorectal cancer | ↑ | increase | XIST/miR-93-5p/HIF-1α/AXL | [73] | |
| ZEB2-AS1 | Gastric cancer | ↑ | increase | ZEB2-AS1/miR-143-5p | [72] | |
| HIF-2α | ||||||
| NEAT1 | Osteosarcoma | ↑ | increase | NEAT1/miR-186-5p/HIF-2α | [91] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barth, D.A.; Prinz, F.; Teppan, J.; Jonas, K.; Klec, C.; Pichler, M. Long-Noncoding RNA (lncRNA) in the Regulation of Hypoxia-Inducible Factor (HIF) in Cancer. Non-Coding RNA 2020, 6, 27. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna6030027
Barth DA, Prinz F, Teppan J, Jonas K, Klec C, Pichler M. Long-Noncoding RNA (lncRNA) in the Regulation of Hypoxia-Inducible Factor (HIF) in Cancer. Non-Coding RNA. 2020; 6(3):27. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna6030027
Chicago/Turabian StyleBarth, Dominik A., Felix Prinz, Julia Teppan, Katharina Jonas, Christiane Klec, and Martin Pichler. 2020. "Long-Noncoding RNA (lncRNA) in the Regulation of Hypoxia-Inducible Factor (HIF) in Cancer" Non-Coding RNA 6, no. 3: 27. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna6030027