Theobromine Improves Working Memory by Activating the CaMKII/CREB/BDNF Pathway in Rats

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
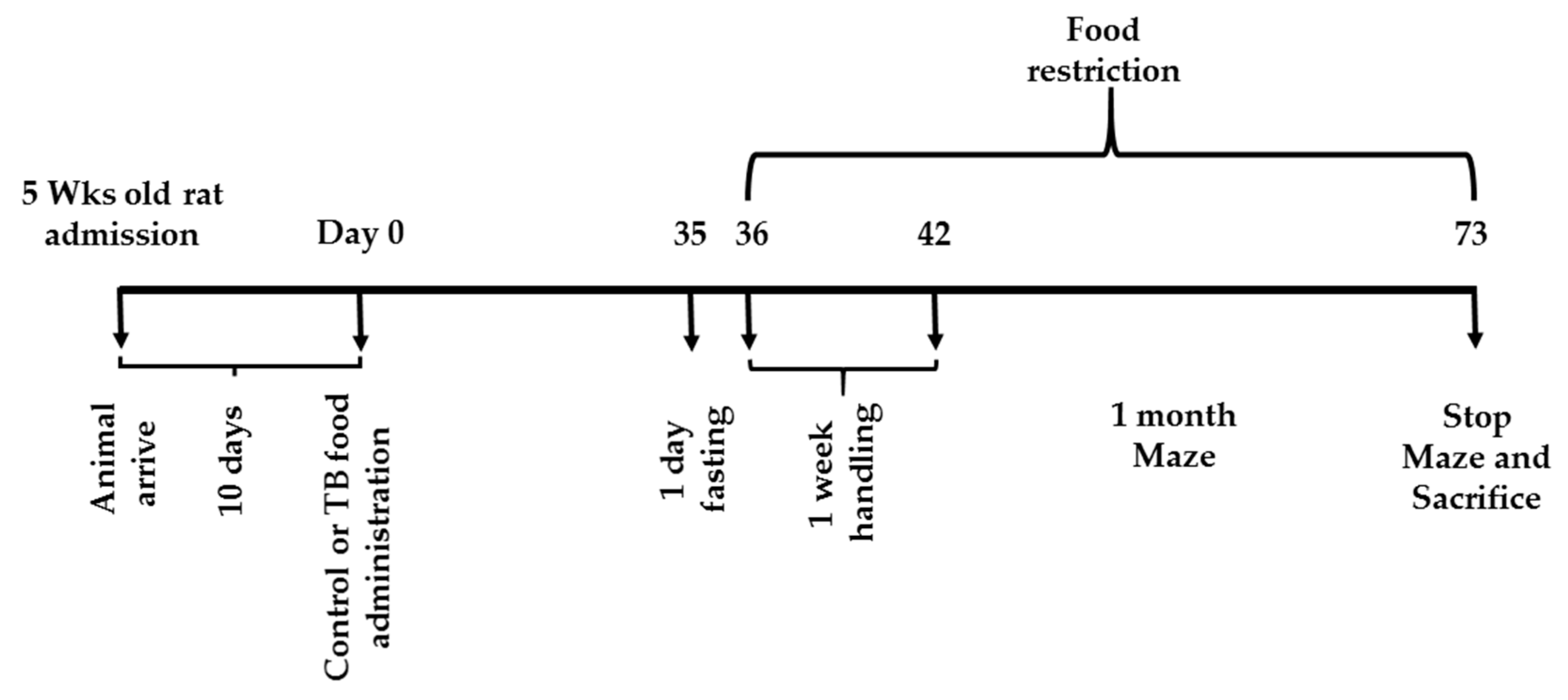
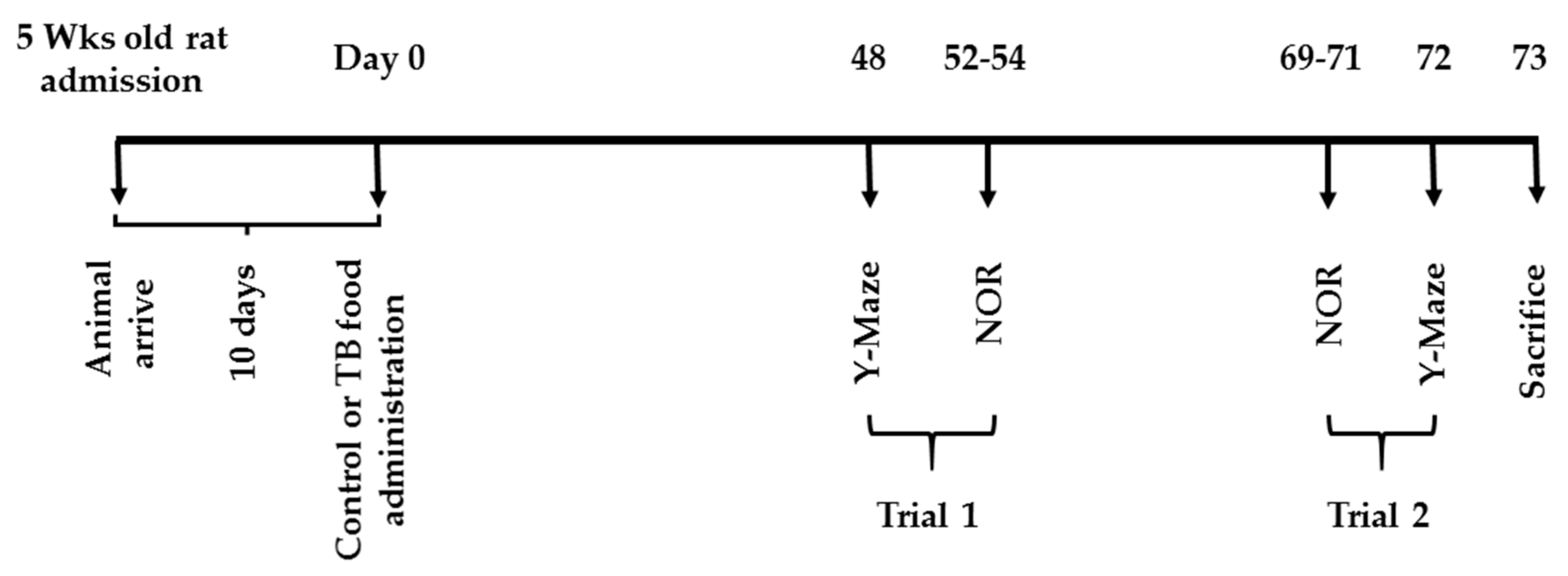
2.2. Feeding and Experiment Schedules
2.2.1. Behavioral Tests
RAM Task (Experiment 1)
Y-Maze Test and NOR Test (Experiment 2)
2.3. Liver and Kidney Function Test
2.4. ELISA
2.5. Western Blot Analysis
2.6. Real Time PCR
2.7. Sample Size of Each Tested Assay
2.8. Statistical Analysis
3. Results
3.1. Experiment 1 (RAM Task)
3.1.1. Body Weight
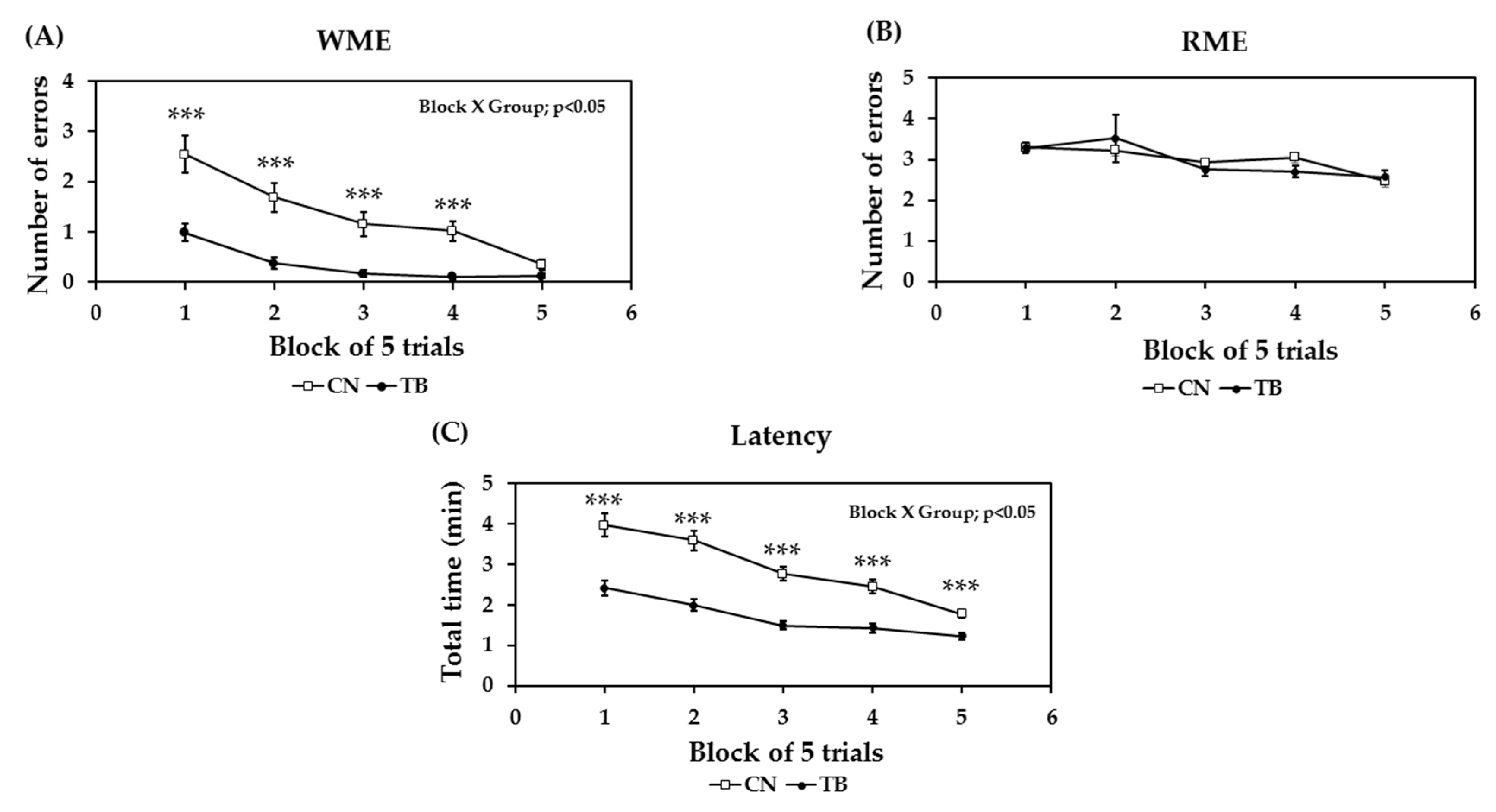
3.1.2. Effects of TB on Memory Related Learning Abilities on the RAM Task
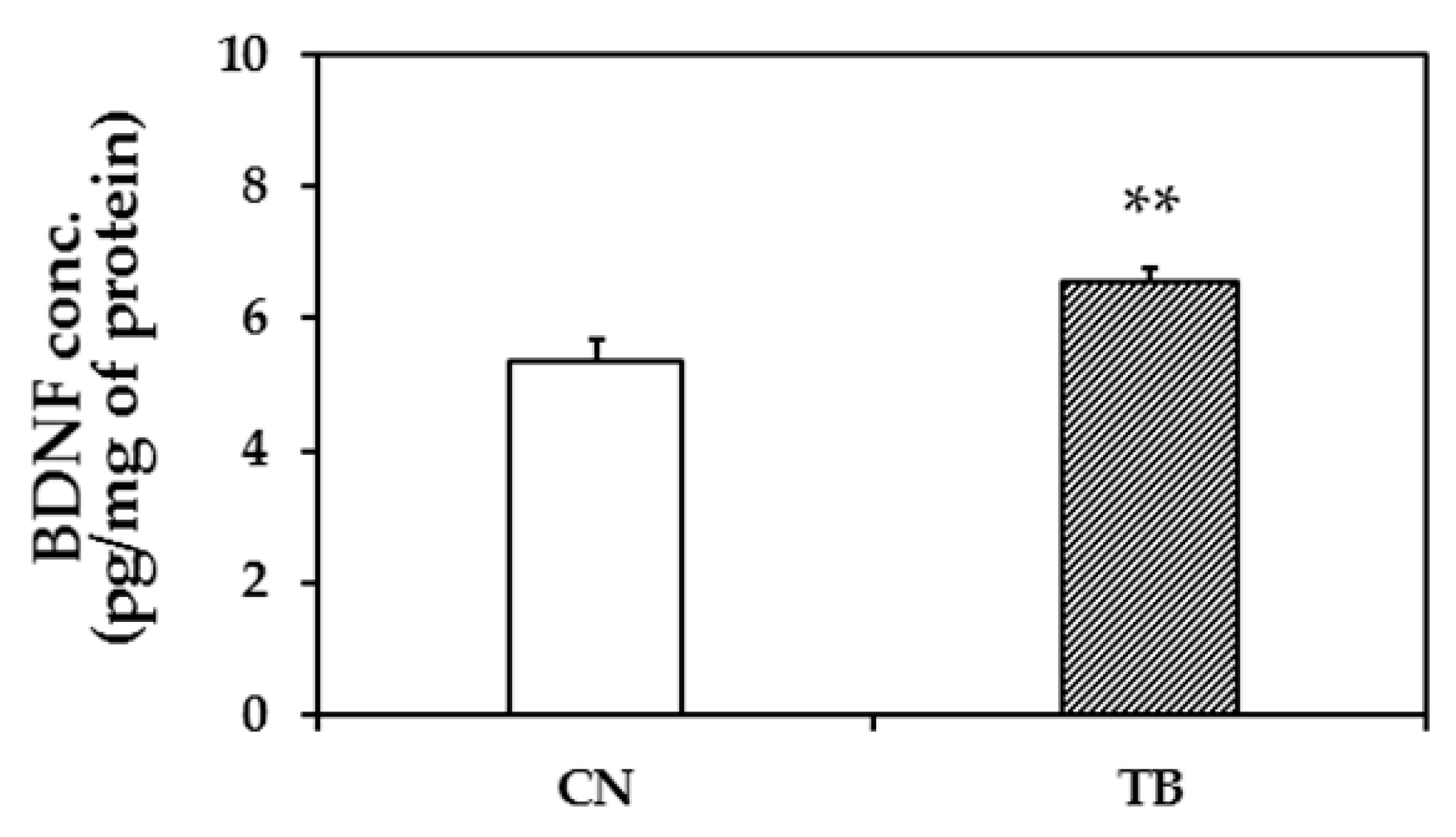
3.1.3. BDNF Protein Expression Level in the mPFC
3.2. Experiment 2 (Y-Maze Test and NOR Test)
3.2.1. Body Weight, Liver Function, and Kidney Function Tests
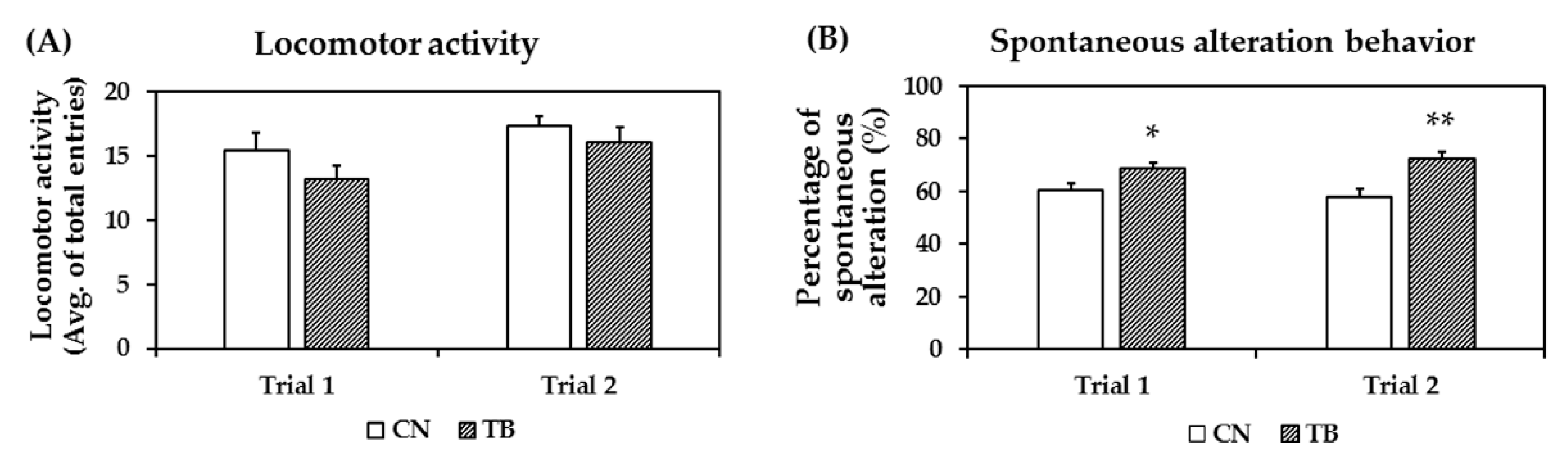
3.2.2. Effects of TB Supplementation on Working Memory Improvement in Y-Maze Test
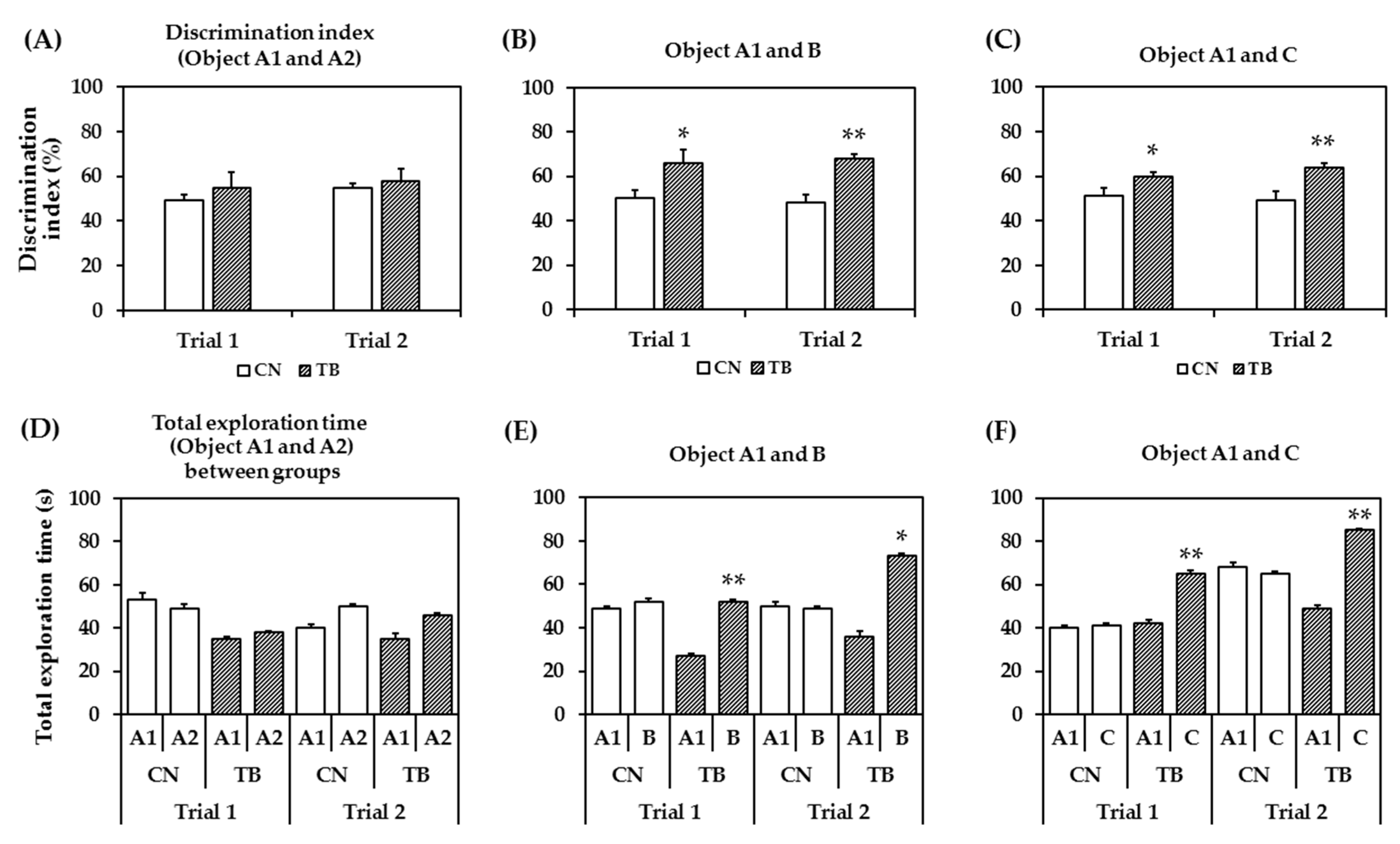
3.2.3. Effects of TB Supplementation on Memory Function in the NOR Test
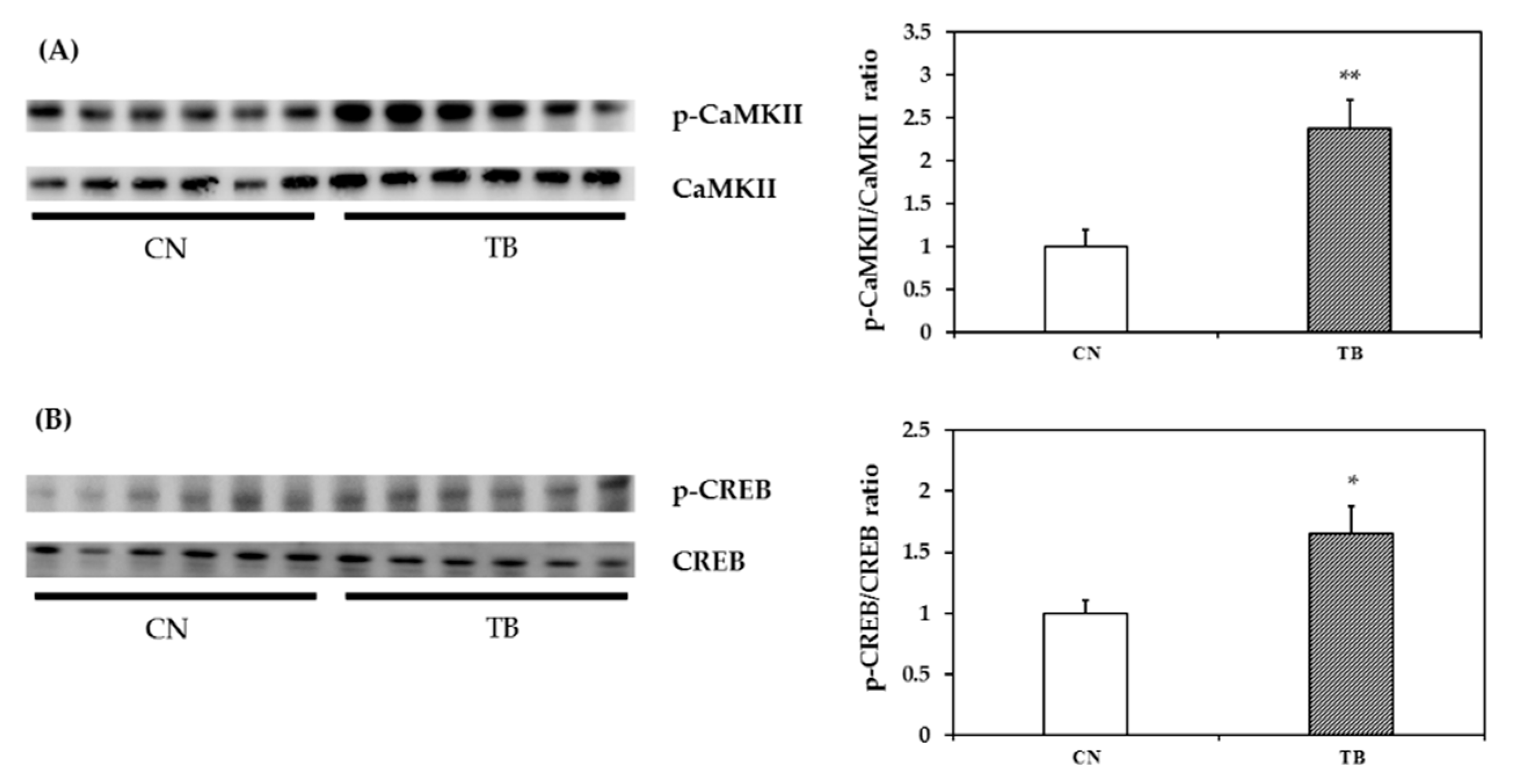
3.2.4. p-CaMKII, p-CREB and BDNF mRNA and Protein Expression in the mPFC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fredholm, B.B.; Bättig, K.; Holmén, J.; Nehlig, A.; Zvartau, E.E. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol. Rev. 1999, 51, 83–133. [Google Scholar]
- Miwa, S.; Sugimoto, N.; Yamamoto, N.; Shirai, T.; Nishida, H.; Hayashi, K.; Kimura, H.; Takeuchi, A.; Igarashi, K.; Yachie, A.; et al. Caffeine induces apoptosis of osteosarcoma cells by inhibiting AKT/mTOR/S6K, NF-κB and MAPK pathways. Anticancer Res. 2012, 32, 3643–3649. [Google Scholar]
- Spencer, J.P.E. Flavonoids and brain health: Multiple effects underpinned by common mechanisms. Genes Nutr. 2009, 4, 243–250. [Google Scholar] [CrossRef]
- Pura Naik, J. Improved high-performance liquid chromatography method to determine theobromine and caffeine in cocoa and cocoa products. J. Agric. Food Chem. 2001, 49, 3579–3583. [Google Scholar] [CrossRef]
- Sokolov, A.N.; Pavlova, M.A.; Klosterhalfen, S.; Enck, P. Chocolate and the brain: Neurobiological impact of cocoa flavanols on cognition and behavior. Neurosci. Biobehav. Rev. 2013, 37, 2445–2453. [Google Scholar] [CrossRef] [PubMed]
- Shively, C.A.; Tarka, S.M. Methylxanthine composition and consumption patterns of cocoa and chocolate products. Prog. Clin. Biol. Res. 1984, 158, 149–178. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, M.; Sugimoto, N.; Katakura, M.; Matsuzaki, K.; Tanigami, H.; Yachie, A.; Ohno-Shosaku, T.; Shido, O. Theobromine up-regulates cerebral brain-derived neurotrophic factor and facilitates motor learning in mice. J. Nutr. Biochem. 2017, 39, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.J.; Spencer, J.P.E.; Rice-Evans, C. Flavonoids: Antioxidants or signalling molecules? Free Radic. Biol. Med. 2004, 36, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, N.; Miwa, S.; Ohno-Shosaku, T.; Tsuchiya, H.; Hitomi, Y.; Nakamura, H.; Tomita, K.; Yachie, A.; Koizumi, S. Activation of tumor suppressor protein PTEN and induction of apoptosis are involved in cAMP-mediated inhibition of cell number in B92 glial cells. Neurosci. Lett. 2011, 497, 55–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, N.; Miwa, S.; Hitomi, Y.; Nakamura, H.; Tsuchiya, H.; Yachie, A. Theobromine, the primary methylxanthine found in Theobroma cacao, prevents malignant glioblastoma proliferation by negatively regulating phosphodiesterase-4, extracellular signal-regulated kinase, Akt/mammalian target of rapamycin kinase, and nuclear fact. Nutr. Cancer 2014, 66, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, N.; Katakura, M.; Matsuzaki, K.; Sumiyoshi, E.; Yachie, A.; Shido, O. Chronic administration of theobromine inhibits mTOR signal in rats. Basic Clin. Pharmacol. Toxicol. 2018, 1–7. [Google Scholar] [CrossRef]
- Khan, Z.U.; Muly, E.C. Molecular mechanisms of working memory. Behav. Brain Res. 2011, 219, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Kolb, B.; Tees, R.C. The Cerebral Cortex of the Rat; The MIT Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Guldin, W.O.; Pritzel, M.; Markowitsch, H.J. Prefrontal cortex of the mouse defined as cortical projection area of the thalamic mediodorsal nucleus. Brain Behav. Evol. 1981, 19, 93–107. [Google Scholar] [CrossRef]
- Jacobsen, C.F. Functions of frontal association area in primates. Arch. Neurol. Psychiatry 1935, 33, 558–569. [Google Scholar] [CrossRef]
- Fuster, J.M.; Alexander, G.E. Neuron activity related to short-term memory. Science 1971, 173, 652–654. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.D.; Perlstein, W.M.; Braver, T.S.; Nystrom, L.E.; Noll, D.C.; Jonides, J.; Smith, E.E. Temporal dynamics of brain activation during a working memory task. Nature 1997, 386, 604–608. [Google Scholar] [CrossRef]
- Goldman-Rakic, P.S. Cellular basis of working memory. Neuron 1995, 14, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Impey, S.; McCorkle, S.R.; Cha-Molstad, H.; Dwyer, J.M.; Yochum, G.S.; Boss, J.M.; McWeeney, S.; Dunn, J.J.; Mandel, G.; Goodman, R.H. Defining the CREB regulon: A genome-wide analysis of transcription factor regulatory regions. Cell 2004, 119, 1041–1054. [Google Scholar] [CrossRef]
- Impey, S.; Smith, D.M.; Obrietan, K.; Donahue, R.; Wade, C.; Storm, D.R. Stimulation of cAMP response element (CRE)-mediated transcription during contextual learning. Nat. Neurosci. 1998, 1, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Barco, A.; Bailey, C.H.; Kandel, E.R. Common molecular mechanisms in explicit and implicit memory. J. Neurochem. 2006, 97, 1520–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Odom, D.T.; Koo, S.-H.; Conkright, M.D.; Canettieri, G.; Best, J.; Chen, H.; Jenner, R.; Herbolsheimer, E.; Jacobsen, E.; et al. Genome-wide analysis of cAMP-response element binding protein occupancy, phosphorylation, and target gene activation in human tissues. Proc. Natl. Acad. Sci. USA 2005, 102, 4459–4464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shieh, P.B.; Hu, S.C.; Bobb, K.; Timmusk, T.; Ghosh, A. Identification of a signaling pathway involved in calcium regulation of BDNF expression. Neuron 1998, 20, 727–740. [Google Scholar] [CrossRef]
- Tao, X.; Finkbeiner, S.; Arnold, D.B.; Shaywitz, A.J.; Greenberg, M.E. Ca2+ influx regulates BDNF transcription by a CREB family transcription factor-dependent mechanism. Neuron 1998, 20, 709–726. [Google Scholar] [CrossRef]
- Gordon, T. The role of neurotrophic factors in nerve regeneration. Neurosurg. Focus 2009, 26, E3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, M.; Thompson, M.A.; Greenberg, M.E. CREB: A Ca(2+)-regulated transcription factor phosphorylated by calmodulin-dependent kinases. Science 1991, 252, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Hossain, S.; Agdul, H.; Shido, O. Docosahexaenoic acid-induced amelioration on impairment of memory learning in amyloid beta-infused rats relates to the decreases of amyloid beta and cholesterol levels in detergent-insoluble membrane fractions. Biochim. Biophys. Acta 2005, 1738, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Mamun, A.; Hashimoto, M.; Katakura, M.; Matsuzaki, K.; Hossain, S.; Arai, H.; Shido, O. Neuroprotective Effect of Madecassoside Evaluated Using Amyloid B1-42-Mediated in Vitro and in Vivo alzheimer’s Disease Models. Int. J. Indig. Med. Plants 2014, 47, 1669–1682. [Google Scholar]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 1998; ISBN 0125476191. [Google Scholar]
- Maurice, T.; Hiramatsu, M.; Itoh, J.; Kameyama, T.; Hasegawa, T.; Nabeshima, T. Behavioral evidence for a modulating role of sigma ligands in memory processes. I. Attenuation of dizocilpine (MK-801)-induced amnesia. Brain Res. 1994, 647, 44–56. [Google Scholar] [CrossRef]
- Kameyama, T.; Ukai, M.; Shinkai, N. Ameliorative effects of tachykinins on scopolamine-induced impairment of spontaneous alternation performance in mice. Methods Find. Exp. Clin. Pharmacol. 1998, 20, 555–560. [Google Scholar] [CrossRef]
- Ukai, M.; Shinkai, N.; Kameyama, T. Involvement of dopamine receptors in beneficial effects of tachykinins on scopolamine-induced impairment of alternation performance in mice. Eur. J. Pharmacol. 1998, 350, 39–45. [Google Scholar] [CrossRef]
- Dossat, A.M.; Jourdi, H.; Wright, K.N.; Strong, C.E.; Sarkar, A.; Kabbaj, M. Viral-mediated Zif268 expression in the prefrontal cortex protects against gonadectomy-induced working memory, long-term memory, and social interaction deficits in male rats. Neuroscience 2017, 340, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, N.; Suemaru, K.; Li, B.; Araki, H. Effects of repeated electroconvulsive seizures on spontaneous alternation behavior and locomotor activity in rats. Biol. Pharm. Bull. 2008, 31, 1928–1932. [Google Scholar] [CrossRef]
- Nakamura, K.; Ito, M.; Liu, Y.; Seki, T.; Suzuki, T.; Arai, H. Effects of single and repeated electroconvulsive stimulation on hippocampal cell proliferation and spontaneous behaviors in the rat. Brain Res. 2013, 1491, 88–97. [Google Scholar] [CrossRef]
- Ennaceur, A.; Delacour, J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav. Brain Res. 1988, 31, 47–59. [Google Scholar] [CrossRef]
- Clark, R.E.; Zola, S.M.; Squire, L.R. Impaired recognition memory in rats after damage to the hippocampus. J. Neurosci. 2000, 20, 8853–8860. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.L.; Lewis, M.J.; Sebastian, V.; Serrano, P.; Luine, V.N. Alcohol administration blocks stress-induced impairments in memory and anxiety, and alters hippocampal neurotransmitter receptor expression in male rats. Horm. Behav. 2013, 63, 659–666. [Google Scholar] [CrossRef]
- Reichel, C.M.; Schwendt, M.; McGinty, J.F.; Olive, M.F.; See, R.E. Loss of object recognition memory produced by extended access to methamphetamine self-administration is reversed by positive allosteric modulation of metabotropic glutamate receptor 5. Neuropsychopharmacology 2011, 36, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.; Renaudineau, S.; Poirier, R.; Poucet, B.; Save, E.; Laroche, S. The formation and stability of recognition memory: What happens upon recall? Front. Behav. Neurosci. 2010, 4, 177. [Google Scholar] [CrossRef]
- Hammond, R.S.; Tull, L.E.; Stackman, R.W. On the delay-dependent involvement of the hippocampus in object recognition memory. Neurobiol. Learn. Mem. 2004, 82, 26–34. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Katakura, M.; Sugimoto, N.; Hara, T.; Hashimoto, M.; Shido, O. Neural progenitor cell proliferation in the hypothalamus is involved in acquired heat tolerance in long-term heat-acclimated rats. PLoS ONE 2017, 12, e0178787. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Katakura, M.; Sugimoto, N.; Hara, T.; Hashimoto, M.; Shido, O. β-amyloid infusion into lateral ventricle alters behavioral thermoregulation and attenuates acquired heat tolerance in rats. Temperature 2015, 2, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, K.; Yamakuni, T.; Hashimoto, M.; Haque, A.M.; Shido, O.; Mimaki, Y.; Sashida, Y.; Ohizumi, Y. Nobiletin restoring beta-amyloid-impaired CREB phosphorylation rescues memory deterioration in Alzheimer’s disease model rats. Neurosci. Lett. 2006, 400, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.E.; Matsuzaki, K.; Katakura, M.; Sugimoto, N.; Mamun, A.; Islam, R.; Hashimoto, M.; Shido, O. Direct exposure to mild heat promotes proliferation and neuronal differentiation of neural stem/progenitor cells in vitro. PLoS ONE 2017, 12, e0190356. [Google Scholar] [CrossRef] [PubMed]
- Li, G.H.; Katakura, M.; Maruyama, M.; Enhkjargal, B.; Matsuzaki, K.; Hashimoto, M.; Shido, O. Changes of noradrenaline-induced contractility and gene expression in aorta of rats acclimated to heat in two different modes. Eur. J. Appl. Physiol. 2008, 104, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pinilla, E.; Oñatibia-Astibia, A.; Franco, R. The relevance of theobromine for the beneficial effects of cocoa consumption. Front. Pharmacol. 2015, 6, 30. [Google Scholar] [CrossRef]
- Sarter, M.; Bodewitz, G.; Stephens, D.N. Attenuation of scopolamine-induced impairment of spontaneous alternation behaviour by antagonist but not inverse agonist and agonist β-carbolines. Psychopharmacology 1988. [Google Scholar] [CrossRef]
- Ennaceur, A.; Neave, N.; Aggleton, J.P. Spontaneous object recognition and object location memory in rats: The effects of lesions in the cingulate cortices, the medial prefrontal cortex, the cingulum bundle and the fornix. Exp. Brain Res. 1997, 113, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Quillfeldt, J.A. Behavioral methods to study learning and memory in rats. In Rodent Model as Tools in Ethical Biomedical Research; Springer: Cham, Switzerland, 2016; pp. 271–311. [Google Scholar]
- Yoon, T.; Okada, J.; Jung, M.W.; Kim, J.J. Prefrontal cortex and hippocampus subserve different components of working memory in rats. Learn. Mem. 2008, 15, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Bhattarai, J. Phasic and tonic type A γ-Aminobutryic acid receptor mediated effect of Withania somnifera on mice hippocampal CA1 pyramidal Neurons. J. Ayurveda Integr. Med. 2014, 5, 216. [Google Scholar] [CrossRef]
- Ravichandran, V.; Kim, M.; Han, S.; Cha, Y. Stachys sieboldii Extract Supplementation Attenuates Memory Deficits by Modulating BDNF-CREB and Its Downstream Molecules, in Animal Models of Memory Impairment. Nutrients 2018, 10, 917. [Google Scholar] [CrossRef]
- Bontempi, B.; Laurent-Demir, C.; Destrade, C.; Jaffard, R. Time-dependent reorganization of brain circuitry underlying long-term memory storage. Nature 1999, 400, 671. [Google Scholar] [CrossRef]
- Mumby, D.G.; Glenn, M.J. Anterograde and retrograde memory for object discriminations and places in rats with perirhinal cortex lesions. Behav. Brain Res. 2000, 114, 119–134. [Google Scholar] [CrossRef]
- Ramos, J.M.J. Long-term spatial memory in rats with hippocampal lesions. Eur. J. Neurosci. 2000, 12, 3375–3384. [Google Scholar] [CrossRef] [PubMed]
- Witter, M.P.; Naber, P.A.; Van Haeften, T.; Machielsen, W.C.M.; Rombouts, S.A.R.B.; Barkhof, F.; Scheltens, P.; da Silva, F.H. Cortico-hippocampal communication by way of parallel parahippocampal-subicular pathways. Hippocampus 2000, 10, 398–410. [Google Scholar] [CrossRef]
- Graff, J.; Woldemichael, B.T.; Berchtold, D.; Dewarrat, G.; Mansuy, I.M. Dynamic histone marks in the hippocampus and cortex facilitate memory consolidation. Nat. Commun. 2012, 3, 991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankland, P.W.; Bontempi, B.; Talton, L.E.; Kaczmarek, L.; Silva, A.J. The involvement of the anterior cingulate cortex in remote contextual fear memory. Science 2004, 304, 881–883. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.W.; Baeg, E.H.; Kim, M.J.; Kim, Y.B.; Kim, J.J. Plasticity and memory in the prefrontal cortex. Rev. Neurosci. 2008, 19, 29–46. [Google Scholar] [CrossRef]
- Nieuwenhuis, I.L.C.; Takashima, A. The role of the ventromedial prefrontal cortex in memory consolidation. Behav. Brain Res. 2011, 218, 325–334. [Google Scholar] [CrossRef]
- Squire, L.R.; Alvarez, P. Retrograde amnesia and memory consolidation: A neurobiological perspective. Curr. Opin. Neurobiol. 1995, 5, 169–177. [Google Scholar] [CrossRef]
- Carew, T.J. Molecular enhancement of memory formation. Neuron 1996, 16, 5–8. [Google Scholar] [CrossRef]
- Martin, K.C.; Barad, M.; Kandel, E.R. Local protein synthesis and its role in synapse-specific plasticity. Curr. Opin. Neurobiol. 2000, 10, 587–592. [Google Scholar] [CrossRef]
- Kelleher, R.J., III; Govindarajan, A.; Tonegawa, S. Translational regulatory mechanisms in persistent forms of synaptic plasticity. Neuron 2004, 44, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Briand, L.A.; Gritton, H.; Howe, W.M.; Young, D.A.; Sarter, M. Modulators in concert for cognition: Modulator interactions in the prefrontal cortex. Prog. Neurobiol. 2007, 83, 69–91. [Google Scholar] [CrossRef]
- Dudai, Y. Molecular bases of long-term memories: A question of persistence. Curr. Opin. Neurobiol. 2002, 12, 211–216. [Google Scholar] [CrossRef]
- Lamprecht, R.; LeDoux, J. Structural plasticity and memory. Nat. Rev. Neurosci. 2004, 5, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Poo, M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2013, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Henderson, C.E. Role of neurotrophic factors in neuronal development. Curr. Opin. Neurobiol. 1996, 6, 64–70. [Google Scholar] [CrossRef]
- Levi-Montalcini, R. The nerve growth factor 35 years later. Science 1987, 237, 1154–1162. [Google Scholar] [CrossRef]
- Davies, A.M. The neurotrophic hypothesis: Where does it stand? Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 389–394. [Google Scholar]
- Miyamoto, E. Molecular mechanism of neuronal plasticity: Induction and maintenance of long-term potentiation in the hippocampus. J. Pharmacol. Sci. 2006, 100, 433–442. [Google Scholar] [CrossRef]
- Leinninger, G.M.; Backus, C.; Uhler, M.D.; Lentz, S.I.; Feldman, E.L. Phosphatidylinositol 3-kinase and Akt effectors mediate insulin-like growth factor-I neuroprotection in dorsal root ganglia neurons. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2004, 18, 1544–1546. [Google Scholar] [CrossRef] [PubMed]
- Vitolo, O.V.; Sant’Angelo, A.; Costanzo, V.; Battaglia, F.; Arancio, O.; Shelanski, M. Amyloid beta -peptide inhibition of the PKA/CREB pathway and long-term potentiation: Reversibility by drugs that enhance cAMP signaling. Proc. Natl. Acad. Sci. USA 2002, 99, 13217–13221. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Brinton, R.D. Vasopressin-induced cytoplasmic and nuclear calcium signaling in embryonic cortical astrocytes: Dynamics of calcium and calcium-dependent kinase translocation. J. Neurosci. 2003, 23, 4228–4239. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Liu, J.; Ye, Z.; Huang, J.; He, F.; Xiao, W.; Hu, X.; Luo, Z. CaMKII-Mediated CREB phosphorylation is involved in Ca2+-Induced BDNF mRNA transcription and neurite outgrowth promoted by electrical stimulation. PLoS ONE 2016, 11, e0162784. [Google Scholar] [CrossRef]
- Kandel, E.R. The molecular biology of memory storage: A dialogue between genes and synapses. Science 2001, 294, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Schulman, H.; Hanson, P.I. Multifunctional Ca2+/calmodulin-dependent protein kinase. Neurochem. Res. 1993, 18, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, M.B. Signal transduction molecules at the glutamatergic postsynaptic membrane. Brain Res. Brain Res. Rev. 1998, 26, 243–257. [Google Scholar] [CrossRef]
- Lisman, J.; Yasuda, R.; Raghavachari, S. Mechanisms of CaMKII action in long-term potentiation. Nat. Rev. Neurosci. 2012, 13, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Korte, M.; Kang, H.; Bonhoeffer, T.; Schuman, E. A role for BDNF in the late-phase of hippocampal long-term potentiation. Neuropharmacology 1998, 37, 553–559. [Google Scholar] [CrossRef]
- Lessmann, V. Neurotrophin-dependent modulation of glutamatergic synaptic transmission in the mammalian CNS. Gen. Pharmacol. 1998, 31, 667–674. [Google Scholar] [CrossRef]
- Bimonte, H.A.; Nelson, M.E.; Granholm, A.-C.E. Age-related deficits as working memory load increases: Relationships with growth factors. Neurobiol. Aging 2003, 24, 37–48. [Google Scholar] [CrossRef]
- Bimonte-Nelson, H.A.; Hunter, C.L.; Nelson, M.E.; Granholm, A.-C.E. Frontal cortex BDNF levels correlate with working memory in an animal model of Down syndrome. Behav. Brain Res. 2003, 139, 47–57. [Google Scholar] [CrossRef]
- Arnsten, A.F.T.; Ramos, B.P.; Birnbaum, S.G.; Taylor, J.R. Protein kinase A as a therapeutic target for memory disorders: Rationale and challenges. Trends Mol. Med. 2005, 11, 121–128. [Google Scholar] [CrossRef]
- Germano, C.; Kinsella, G.J. Working memory and learning in early Alzheimer’s disease. Neuropsychol. Rev. 2005, 15, 1–10. [Google Scholar] [CrossRef]
- Goldman-Rakic, P.S.; Castner, S.A.; Svensson, T.H.; Siever, L.J.; Williams, G.V. Targeting the dopamine D1 receptor in schizophrenia: Insights for cognitive dysfunction. Psychopharmacology 2004, 174, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.G.; Baddeley, A.D. Primary and working memory functioning in Alzheimer-type dementia. J. Clin. Exp. Neuropsychol. 1988, 10, 279–296. [Google Scholar] [CrossRef]
- Castellanos, F.X.; Tannock, R. Neuroscience of attention-deficit/hyperactivity disorder: The search for endophenotypes. Nat. Rev. Neurosci. 2002, 3, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Yabuki, Y.; Shioda, N.; Maeda, T.; Hiraide, S.; Togashi, H.; Fukunaga, K. Aberrant CaMKII activity in the medial prefrontal cortex is associated with cognitive dysfunction in ADHD model rats. Brain Res. 2014, 1557, 90–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabir, A.; Hooton, A.; Tallis, J.; F Higgins, M. The Influence of Caffeine Expectancies on Sport, Exercise, and Cognitive Performance. Nutrients 2018, 10, 1528. [Google Scholar] [CrossRef]
- Ritchie, K.; Carriere, I.; de Mendonca, A.; Portet, F.; Dartigues, J.F.; Rouaud, O.; Barberger-Gateau, P.; Ancelin, M.L. The neuroprotective effects of caffeine: A prospective population study (the Three City Study). Neurology 2007, 69, 536–545. [Google Scholar] [CrossRef] [PubMed]
- van Gelder, B.M.; Buijsse, B.; Tijhuis, M.; Kalmijn, S.; Giampaoli, S.; Nissinen, A.; Kromhout, D. Coffee consumption is inversely associated with cognitive decline in elderly European men: The FINE Study. Eur. J. Clin. Nutr. 2007, 61, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.; Lunet, N.; Azevedo, A.; de Mendonca, A.; Ritchie, K.; Barros, H. Caffeine intake is associated with a lower risk of cognitive decline: A cohort study from Portugal. J. Alzheimers. Dis. 2010, 20 (Suppl. 1), S175–S185. [Google Scholar] [CrossRef] [PubMed]
- Alzoubi, K.H.; Mhaidat, N.M.; Obaid, E.A.; Khabour, O.F. Caffeine Prevents Memory Impairment Induced by Hyperhomocysteinemia. J. Mol. Neurosci. 2018, 66, 222–228. [Google Scholar] [CrossRef]
- Chen, X.; Ghribi, O.; Geiger, J.D. Caffeine protects against disruptions of the blood-brain barrier in animal models of Alzheimer’s and Parkinson’s diseases. J. Alzheimers. Dis. 2010, 20 (Suppl. 1), S127–S141. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight (g) | p Value | ||
|---|---|---|---|
| CN Rats | TB Rats | ||
| Initial day (day 0) | 239.2 ± 7.13 | 229.9 ± 5.20 | 0.32 |
| Half way (day 45) | 360.6 ± 3.95 | 353.6 ± 4.98 | 0.29 |
| Final (day 73) | 384.9 ± 3.10 | 375.5 ± 4.65 | 0.12 |
| Body Weight (g) | p Value | ||
|---|---|---|---|
| CN Rats | TB Rats | ||
| Initial day (day 0) | 183.1 ± 2.03 | 185.5 ± 2.52 | 0.47 |
| Half way (day 45) | 416.0 ± 5.67 | 428.3 ± 9.92 | 0.30 |
| Final (day 73) | 463.0 ± 7.89 | 481.0 ± 11.3 | 0.21 |
| Liver Function Test | Kidney Function Test | ||||||
|---|---|---|---|---|---|---|---|
| Biochemical Parameters | CN Rats | TB Rats | p Value | Biochemical Parameters | CN Rats | TB Rats | p Value |
| GOT/AST (IU/L) | 67.4 ± 8.43 | 63.7 ± 8.14 | 0.76 | T-Pro (g/dl) | 5.4 ± 0.10 | 5.2 ± 0.12 | 0.09 |
| GPT/ALT (IU/L) | 25.1 ± 1.74 | 23.7 ± 2.23 | 0.63 | Alb (g/dl) | 3.1 ± 0.05 | 3.0 ± 0.07 | 0.50 |
| GGT (IU/L) | 2.7 ± 0.21 | 3.3 ± 0.21 | 0.06 | BUN (mg/dl) | 16.6 ± 0.40 | 16.3 ± 0.50 | 0.64 |
| T-Cho (mg/dl) | 79.0 ± 4.94 | 90.6 ± 5.74 | 0.14 | UA (mg/dl) | 1.2 ± 0.06 | 1.0 ± 0.09 | 0.11 |
| TG (mg/dl) | 95.0 ± 10.97 | 102.6 ± 13.44 | 0.67 | Cre-2 (mg/dl) | 0.3 ± 0.02 | 0.3 ± 0.02 | 0.75 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, R.; Matsuzaki, K.; Sumiyoshi, E.; Hossain, M.E.; Hashimoto, M.; Katakura, M.; Sugimoto, N.; Shido, O. Theobromine Improves Working Memory by Activating the CaMKII/CREB/BDNF Pathway in Rats. Nutrients 2019, 11, 888. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11040888
Islam R, Matsuzaki K, Sumiyoshi E, Hossain ME, Hashimoto M, Katakura M, Sugimoto N, Shido O. Theobromine Improves Working Memory by Activating the CaMKII/CREB/BDNF Pathway in Rats. Nutrients. 2019; 11(4):888. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11040888
Chicago/Turabian StyleIslam, Rafiad, Kentaro Matsuzaki, Eri Sumiyoshi, Md Emon Hossain, Michio Hashimoto, Masanori Katakura, Naotoshi Sugimoto, and Osamu Shido. 2019. "Theobromine Improves Working Memory by Activating the CaMKII/CREB/BDNF Pathway in Rats" Nutrients 11, no. 4: 888. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11040888