n-3 Fatty Acid Supplementation and Leukocyte Telomere Length in Patients with Chronic Kidney Disease

Abstract
:1. Introduction
2. Volunteers and Methods
2.1. Participants
2.2. Study Design
2.3. Measurement of Leukocyte Telomere Length and Plasma F2-Isoprostanes
2.4. Statistical Analysis
3. Results
3.1. Telomere Length at Baselinein Patients with CKD
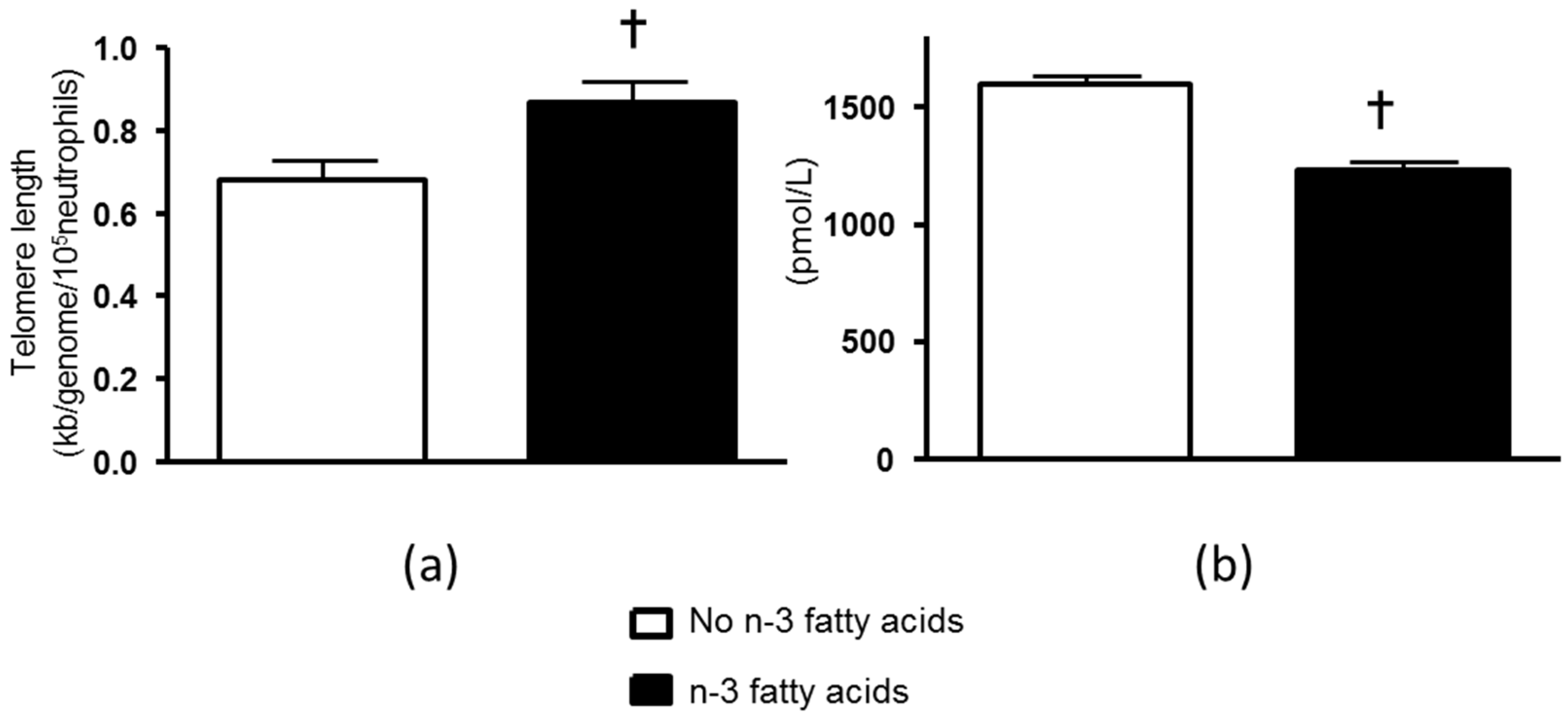
3.2. Effect of n-3 Fatty Acids and CoQ on Telomere Length
3.3. Effect of n-3 Fatty Acids and CoQ on Telomere Length Corrected for Cell Count
3.4. Relationship between Plasma F2-Isoprostanes and Telomere Length
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Aviv, A. Telomeres and human somatic fitness. J. Gerontol. Ser. A-Biol. Sci. Med. Sci. 2006, 61, 871–873. [Google Scholar] [CrossRef]
- Lipcsey, M.; Soederberg, E.; Basu, S.; Larsson, A.; Sjoelin, J.; Astroem, M.; Eriksson, M.B. F-2-isoprostane, inflammation, cardiac function and oxygenation in the endotoxaemic pig. Prostag. Leukotr. Ess. 2008, 78, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Bansal, N.; Whooley, M.A.; Regan, M.; McCulloch, C.E.; Ix, J.H.; Epel, E.; Blackburn, E.; Lin, J.; Hsu, C.-Y. Association between kidney function and telomere length: The heart and soul study. Am. J. Nephrol. 2012, 36, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Boxall, M.C.; Goodship, T.H.J.; Brown, A.L.; Ward, M.C.; von Zglinicki, T. Telomere shortening and haemodialysis. Blood Purif. 2006, 24, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Tsirpanlis, G.; Chatzipanagiotou, S.; Boufidou, F.; Kordinas, V.; Alevyzaki, F.; Zoga, M.; Kyritsis, I.; Stamatelou, K.; Triantafyllis, G.; Nicolaou, C. Telomerase activity is decreased in peripheral blood mononuclear cells of hemodialysis patients. Am. J. Nephrol. 2006, 26, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Carrero, J.J.; Stenvinkel, P.; Fellstrom, B.; Qureshi, A.R.; Lamb, K.; Heimburger, O.; Barany, P.; Radhakrishnan, K.; Lindholm, B.; Soveri, I.; et al. Telomere attrition is associated with inflammation, low fetuin-a levels and high mortality in prevalent haemodialysis patients. J. Intern. Med. 2008, 263, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Aviv, A.; Chen, W.; Gardner, J.P.; Kimura, M.; Brimacombe, M.; Cao, X.; Srinivasan, S.R.; Berenson, G.S. Leukocyte telomere dynamics: Longitudinal findings among young adults in the Bogalusa heart study. Am. J. Epidemiol. 2009, 169, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.L.; Fitzpatrick, A.L.; Boudreau, R.M.; Arnold, A.M.; Aviv, A.; Kimura, M.; Fried, L.F.; Harris, T.B.; Newman, A.B. Leukocyte telomere length is associated with noninvasively measured age-related disease: The cardiovascular health study. J. Gerontol. Ser. A-Biol. Sci. Med. Sci. 2012, 67, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Shammas, M.A. Telomeres, lifestyle, cancer, and aging. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Pavlaki, K.I.; Kastrinaki, M.-C.; Klontzas, M.; Velegraki, M.; Mavroudi, I.; Papadaki, H.A. Abnormal telomere shortening of peripheral blood mononuclear cells and granulocytes in patients with chronic idiopathic neutropenia. Haematol. Hematol. J. 2012, 97, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Brummendorf, T.H.; Maciejewski, J.P.; Mak, J.; Young, N.S.; Lansdorp, P.M. Telomere length in leukocyte subpopulations of patients with aplastic anemia. Blood 2001, 97, 895–900. [Google Scholar] [CrossRef] [PubMed]
- Compte, N.; Bailly, B.; De Breucker, S.; Goriely, S.; Pepersack, T. Study of the association of total and differential white blood cell counts with geriatric conditions, cardio-vascular diseases, seric il-6 levels and telomere length. Exp. Gerontol. 2015, 61, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Sela, S.; Shurtz-Swirski, R.; Cohen-Mazor, M.; Mazor, R.; Chezar, J.; Shapiro, G.; Hassan, K.; Shkolnik, G.; Geron, R.; Kristal, B. Primed peripheral polymorphonuclear leukocyte: A culprit underlying chronic low-grade inflammation and systemic oxidative stress in chronic kidney disease. J. Am. Soc. Nephrol. 2005, 16, 2431–2438. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Puddey, I.B.; Burke, V.; Croft, K.D.; Dunstan, D.W.; Rivera, J.H.; Beilin, L.J. Effect of ω 3 fatty acids on oxidative stress in humans: GC-MS measurement of urinary F2-isoprostane excretion. Redox Rep. 2000, 5, 45–46. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Dunstan, D.W.; Burke, V.; Croft, K.D.; Rivera, J.H.; Beilin, L.J.; Puddey, I.B. Effect of dietary fish and exercise training on urinary F2-isoprostane excretion in non-insulin-dependent diabetic patients. Metabolism 1999, 48, 1402–1408. [Google Scholar] [CrossRef]
- Mas, E.; Woodman, R.J.; Burke, V.; Puddey, I.B.; Beilin, L.J.; Durand, T.; Mori, T.A. The omega-3 fatty acids EPA and DHA decrease plasma F2-isoprostanes: Results from two placebo-controlled interventions. Free Radic. Res. 2010, 44, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.; Devaraj, S.; Jialal, I. Coenzyme Q10 supplementation and heart failure. Nutr. Rev. 2007, 65, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Alleva, R.; Tomasetti, M.; Battino, M.; Curatola, G.; Littarru, G.P.; Folkers, K. The roles of coenzyme Q10 and vitamin-E on the peroxidation of human low-density-lipoprotein subfractions. Proc. Natl. Acad. Sci. USA 1995, 92, 9388–9391. [Google Scholar] [CrossRef] [PubMed]
- Tomasetti, M.; Littarru, G.P.; Stocker, R.; Alleva, R. Coenzyme Q10 enrichment decreases oxidative DNA damage in human lymphocytes. Free Radic. Biol. Med. 1999, 27, 1027–1032. [Google Scholar] [CrossRef]
- Cachofeiro, V.; Goicochea, M.; Garcia de Vinuesa, S.; Oubina, P.; Lahera, V.; Luno, J. Oxidative stress and inflammation, a link between chronic kidney disease and cardiovascular disease. Kidney Int. 2008, 74, S4–S9. [Google Scholar] [CrossRef] [PubMed]
- Richard, M.J.; Sirajeddine, M.K.; Cordonnier, D.; Tricot, F.; Foulon, T.; Mouneimne, A.; Maurizi, J. Relationship of omega-3 fatty acid supplementation to plasma lipid peroxidation in predialysis patients with hypertriglyceridaemia. Eur. J. Med. 1993, 2, 15–18. [Google Scholar] [PubMed]
- Mori, T.A.; Burke, V.; Puddey, I.B.; Irish, A.B.; Cowpland, C.A.; Beilin, L.J.; Dogra, G.K.; Watts, G.F. The effects of omega 3 fatty acids and coenzyme Q10 on blood pressure and heart rate in chronic kidney disease: A randomized controlled trial. J. Hypertens. 2009, 27, 1863–1872. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Pan, Y.; Kao, S.Y.; Li, C.; Kohane, I.; Chan, J.; Yankner, B.A. Gene regulation and DNA damage in the ageing human brain. Nature 2004, 429, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Cawthon, R.M. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002, 30, e47. [Google Scholar] [CrossRef] [PubMed]
- Tsai, I.J.; Croft, K.D.; Mori, T.A.; Falck, J.R.; Beilin, L.J.; Puddey, I.B.; Barden, A.E. 20-HETE and F2-isoprostanes in the metabolic syndrome: The effect of weight reduction. Free Radic. Biol. Med. 2009, 46, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Woodman, R.J.; Watts, G.F.; Puddey, I.B.; Burke, V.; Mori, T.A.; Hodgson, J.M.; Beilin, L.J. Leukocyte count and vascular function in type 2 diabetic subjects with treated hypertension. Atherosclerosis 2002, 163, 175–181. [Google Scholar] [CrossRef]
- Weng, N.P. Interplay between telomere length and telomerase in human leukocyte differentiation and aging. J. Leukoc. Biol. 2001, 70, 861–867. [Google Scholar] [PubMed]
- Denburg, J.A.; Hatfield, H.M.; Cyr, M.M.; Hayes, L.; Holt, P.G.; Sehmi, R.; Dunstan, J.A.; Prescott, S.L. Fish oil supplementation in pregnancy modifies neonatal progenitors at birth in infants at risk of atopy. Pediatr. Res. 2005, 57, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Summers, C.; Rankin, S.M.; Condliffe, A.M.; Singh, N.; Peters, A.M.; Chilvers, E.R. Neutrophil kinetics in health and disease. Trends Immunol. 2010, 31, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Yao, S.L.; Shu, H.Q.; Liu, H.; Yuan, S.Y.; Chen, X.D.; Li, R.D.; Wu, X.Y.; Guo, L.; Wang, Y. Resolvin D1 attenuates lipopolysaccharide induced acute lung injury through CXCL-12/CXCR4 pathway. J. Surg. Res. 2014, 188, 213–221. [Google Scholar]
- Mas, E.; Barden, A.; Burke, V.; Beilin, L.J.; Watts, G.F.; Huang, R.C.; Puddey, I.B.; Irish, A.B.; Mori, T.A. A randomized controlled trial of the effects of n-3 fatty acids on resolvins in chronic kidney disease. Clin. Nutr. 2015, in press. [Google Scholar] [CrossRef] [PubMed]
- Barden, A.E.; Burke, V.; Mas, E.; Beilin, L.J.; Puddey, I.B.; Watts, G.F.; Irish, A.B.; Mori, T.A. n-3 fatty acids reduce plasma 20-hydroxyeicosatetraenoic acid and blood pressure in patients with chronic kidney disease. J. Hypertens. 2015, 33, 1947–1953. [Google Scholar] [CrossRef] [PubMed]
- Farzaneh-Far, R.; Lin, J.; Epel, E.S.; Harris, W.S.; Blackburn, E.H.; Whooley, M.A. Association of marine omega-3 fatty acid levels with telomeric aging in patients with coronary heart disease. JAMA J. Am. Med. Assoc. 2010, 303, 250–257. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, N.; Parletta, N.; Milte, C.M.; Benassi-Evans, B.; Fenech, M.; Howe, P.R.C. Telomere shortening in elderly individuals with mild cognitive impairment may be attenuated with omega-3 fatty acid supplementation: A randomized controlled pilot study. Nutrition 2014, 30, 489–491. [Google Scholar] [CrossRef] [PubMed]
- Kiecolt-Glaser, J.K.; Epel, E.S.; Belury, M.A.; Andridge, R.; Lin, J.; Glaser, R.; Malarkey, W.B.; Hwang, B.S.; Blackburn, E. Omega-3 fatty acids, oxidative stress, and leukocyte telomere length: A randomized controlled trial. Brain Behav. Immun. 2013, 28, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Cattan, V.; Mercier, N.; Gardner, J.P.; Regnault, V.; Labat, C.; Maki-Jouppila, J.; Nzietchueng, R.; Benetos, A.; Kimura, M.; Aviv, A.; et al. Chronic oxidative stress induces a tissue-specific reduction in telomere length in cast/ei mice. Free Radic. Biol. Med. 2008, 44, 1592–1598. [Google Scholar] [CrossRef] [PubMed]
- Tarry-Adkins, J.L.; Ozanne, S.E.; Norden, A.; Cherif, H.; Hales, C.N. Lower antioxidant capacity and elevated p53 and p21 may be a link between gender disparity in renal telomere shortening, albuminuria, and longevity. Am. J. Physiol. Ren. Physiol. 2006, 290, F509–F516. [Google Scholar] [CrossRef] [PubMed]
- Epel, E.S.; Blackburn, E.H.; Lin, J.; Dhabhar, F.S.; Adler, N.E.; Morrow, J.D.; Cawthon, R.M. Accelerated telomere shortening in response to life stress. Proc. Natl. Acad. Sci. USA 2004, 101, 17312–17315. [Google Scholar] [CrossRef] [PubMed]
- Demissie, S.; Levy, D.; Benjamin, E.J.; Cupples, L.A.; Gardner, J.P.; Herbert, A.; Kimura, M.; Larson, M.G.; Meigs, J.B.; Keaney, J.F.; et al. Insulin resistance, oxidative stress, hypertension, and leukocyte telomere length in men from the framingham heart study. Aging Cell 2006, 5, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Ziegler-Heitbrock, L. Monocyte subsets in man and other species. Cell. Immunol. 2014, 289, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Hellerstein, M.K. Measurement of T-cell kinetics: Recent methodologic advances. Immunol. Today 1999, 20, 438–441. [Google Scholar] [CrossRef]
- Dlouha, D.; Maluskova, J.; Lesna, I.K.; Lanska, V.; Hubacek, J.A. Comparison of the relative telomere length measured in leukocytes and eleven different human tissues. Physiol. Res. 2014, 63, S343–S350. [Google Scholar] [PubMed]
- Lin, J.; Epel, E.; Cheon, J.; Kroenke, C.; Sinclair, E.; Bigos, M.; Wolkowitz, O.; Mellon, S.; Blackburn, E. Analyses and comparisons of telomerase activity and telomere length in human T and B cells: Insights for epidemiology of telomere maintenance. J. Immunol. Methods 2010, 352, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Krysiak, R.; Okopien, B. Lymphocyte-suppressing action of angiotensin-converting enzyme inhibitors in coronary artery disease patients with normal blood pressure. Pharmacol. Rep. 2011, 63, 1151–1161. [Google Scholar] [CrossRef]
- Dincer, Y.; Sekercioglu, N.; Pekpak, M.; Gunes, K.N.; Akcay, T. Assessment of DNA oxidation and antioxidant activity in hypertensive patients with chronic kidney disease. Ren. Fail. 2008, 30, 1006–1011. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Tortosa, M.C.; Granados, S.; Ramirez-Tortosa, C.L.; Ochoa, J.J.; Camacho, P.; Garcia-Valdes, L.; Battino, M.; Quiles, J.L. Oxidative stress status in liver mitochondria and lymphocyte DNA damage of atherosclerotic rabbits supplemented with water soluble coenzyme Q10. Biofactors 2008, 32, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Niklowitz, P.; Sonnenschein, A.; Janetzky, B.; Andler, W.; Menke, T. Enrichment of coenzyme Q10 in plasma and blood cells: Defense against oxidative damage. Int. J. Biol. Sci. 2007, 3, 257–262. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Control (n = 15) | n-3 FA (n = 19) | CoQ (n = 21) | n-3 FA + CoQ (n = 18) | ANOVA at Baseline (p Value) | Interaction (p Value) | ||
|---|---|---|---|---|---|---|---|
| Main Effects (p Value) | |||||||
| n-3 FA | CoQ | ||||||
| Neutrophil telomere length (kb/genome) | |||||||
| Baseline | 163.4 ± 21.5 | 159.1 ± 16.1 | 181.2 ± 14.3 | 149.7 ± 12.3 | NS | ||
| Post | 166.9 ± 11.5 | 160.9 ± 10.3 | 155.6 ± 9.8 | 184.8 ± 10.6 | 29.3 ± 14.6 | 24.6 ± 14.7 | |
| (p = 0.28) | (p = 0.54) | p = 0.1 | |||||
| Neutrophil telomere length corrected for neutrophil count (kb/genome/105 cells) | |||||||
| Baseline | 0.61 ± 0.10 | 0.82 ± 0.12 | 0.97 ± 0.18 | 0.55 ± 0.06 | p = 0.25 | ||
| Post | 0.72 ± 0.08 | 0.91 ± 0.70 | 0.63 ± 0.07 | 0.82 ± 0.08 | 0.19 ± 0.07 | −0.10 ± 0.07 | |
| (p = 0.015) | (p = 0.17) | p = 0.97 | |||||
| PBMC telomere length (kb/genome) | |||||||
| Baseline | 103.9 ± 11.6 | 86.5 ± 11.4 | 114.1 ± 9.8 | 75.0 ± 8.2 * | p = 0.03 | ||
| Post | 107.1 ± 11.6 | 101.9 ± 10.2 | 96.5 ± 9.7 | 107.3 ± 10.5 | 10.5 ± 14.7 | 5.1 ± 14.5 | |
| (p = 0.48) | (p = 0.73) | p = 0.46 | |||||
| PBMC telomere length corrected for PBMC count (kb/genome/105 cells) | |||||||
| Baseline | 1.06 ± 0.14 | 0.89 ± 0.13 | 1.18 ± 0.14 | 0.66 ± 0.11 * | p = 0.03 | ||
| Post | 1.16 ± 0.15 | 0.95 ± 0.14 | 0.99 ± 0.13 | 1.08 ± 0.14 | −0.64 ± 0.14 | −0.02 ± 0.14 | |
| (p = 0.9) | (p = 0.73) | p = 0.29 | |||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barden, A.; O’Callaghan, N.; Burke, V.; Mas, E.; Beilin, L.J.; Fenech, M.; Irish, A.B.; Watts, G.F.; Puddey, I.B.; Huang, R.-C.; et al. n-3 Fatty Acid Supplementation and Leukocyte Telomere Length in Patients with Chronic Kidney Disease. Nutrients 2016, 8, 175. https://0-doi-org.brum.beds.ac.uk/10.3390/nu8030175
Barden A, O’Callaghan N, Burke V, Mas E, Beilin LJ, Fenech M, Irish AB, Watts GF, Puddey IB, Huang R-C, et al. n-3 Fatty Acid Supplementation and Leukocyte Telomere Length in Patients with Chronic Kidney Disease. Nutrients. 2016; 8(3):175. https://0-doi-org.brum.beds.ac.uk/10.3390/nu8030175
Chicago/Turabian StyleBarden, Anne, Nathan O’Callaghan, Valerie Burke, Emile Mas, Lawrence J. Beilin, Michael Fenech, Ashley B. Irish, Gerald F. Watts, Ian B. Puddey, Rae-Chi Huang, and et al. 2016. "n-3 Fatty Acid Supplementation and Leukocyte Telomere Length in Patients with Chronic Kidney Disease" Nutrients 8, no. 3: 175. https://0-doi-org.brum.beds.ac.uk/10.3390/nu8030175