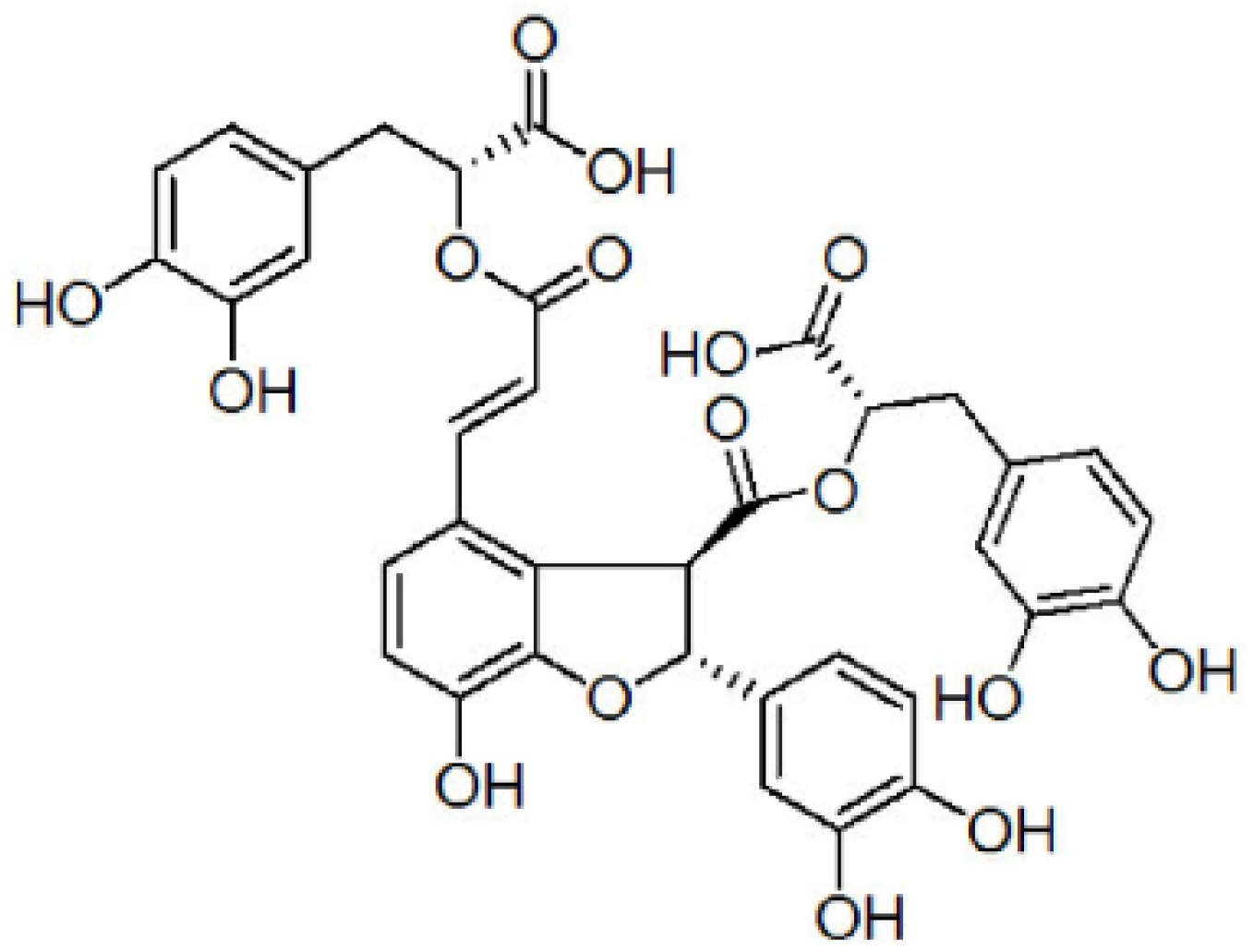
Salvianolic Acid B Inhibits Aβ Generation by Modulating BACE1 Activity in SH-SY5Y-APPsw Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drugs, Reagents and Antibodies
2.2. Cell Culture
2.3. ELISA
2.4. Reactive Oxygen Species (ROS) Production
2.5. MDA, SOD and GSH-Px Assays
2.6. Western Blot Assay
2.7. Statistical Analysis
3. Results
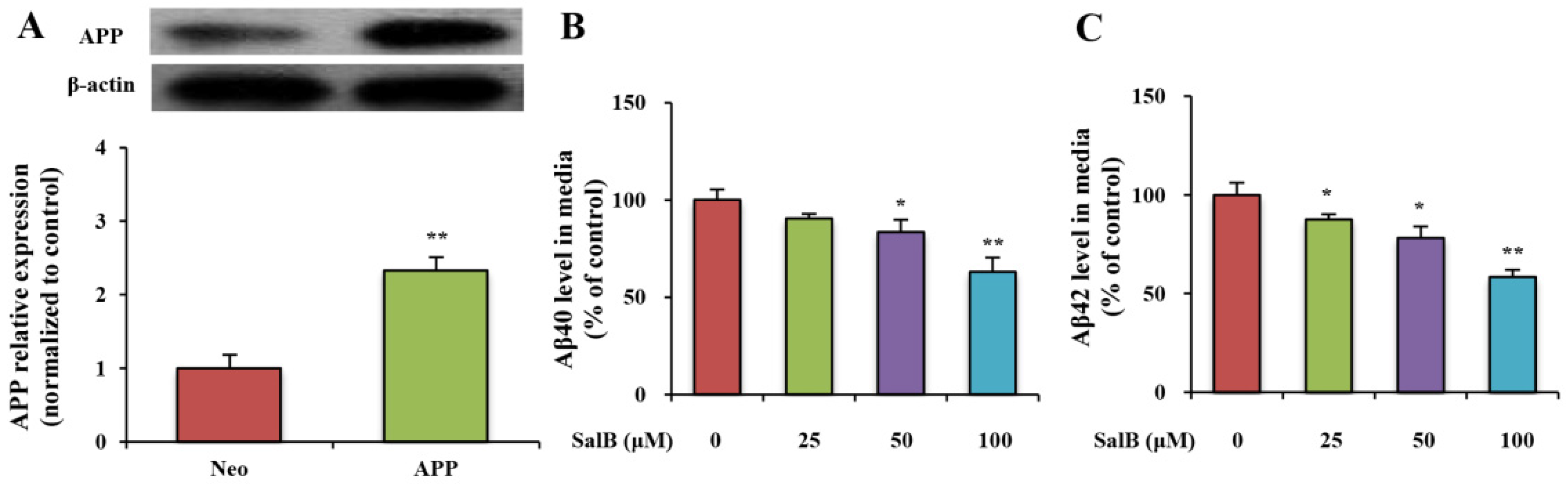
3.1. SalB Reduces the Levels of Aβ40 and Aβ42 in Cell Culture Medium
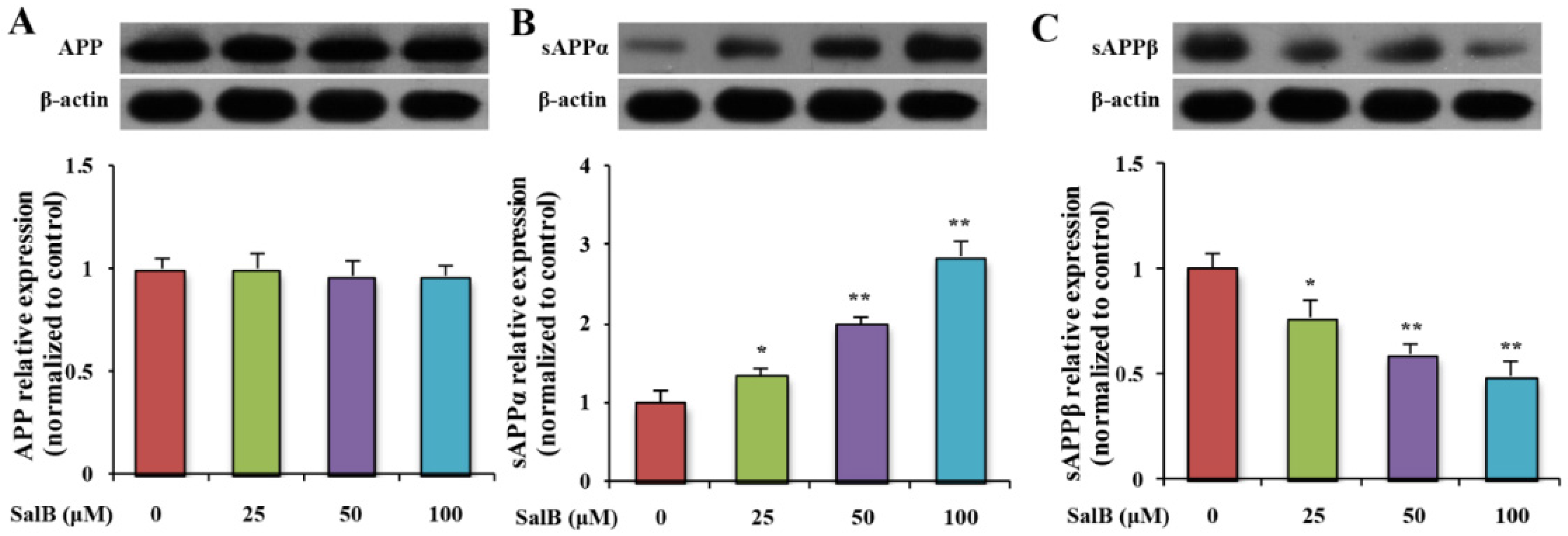
3.2. SalB Reduces the Level of sAPPβ in SH-SY5Y-APPsw Cells
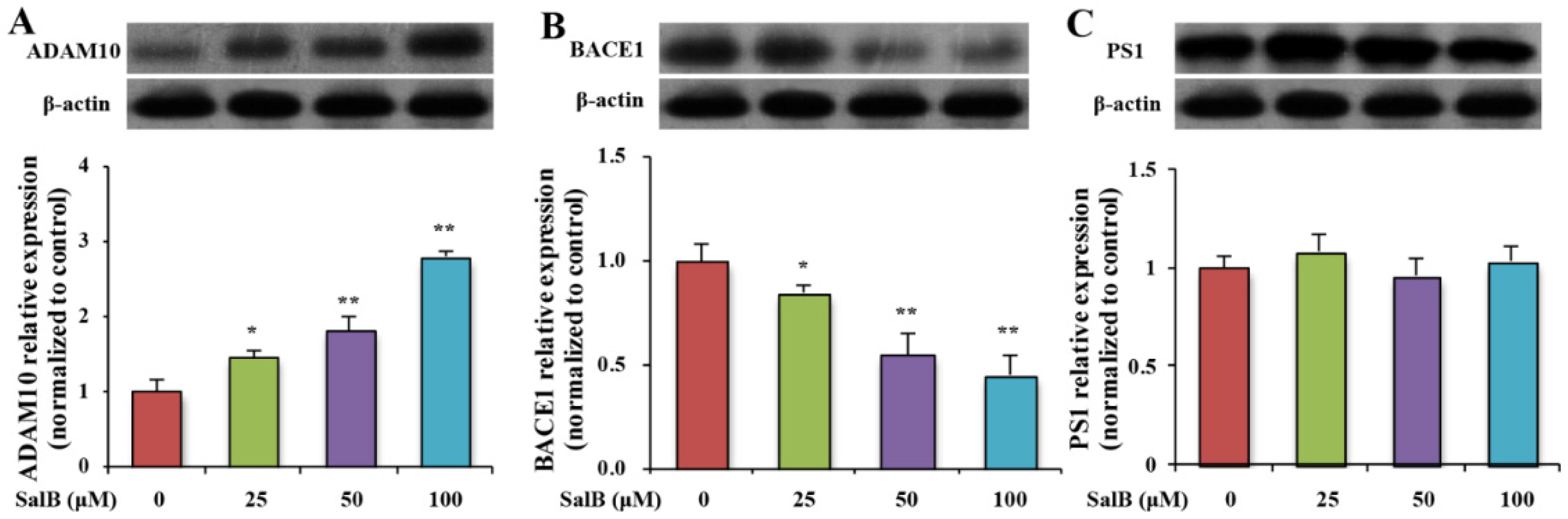
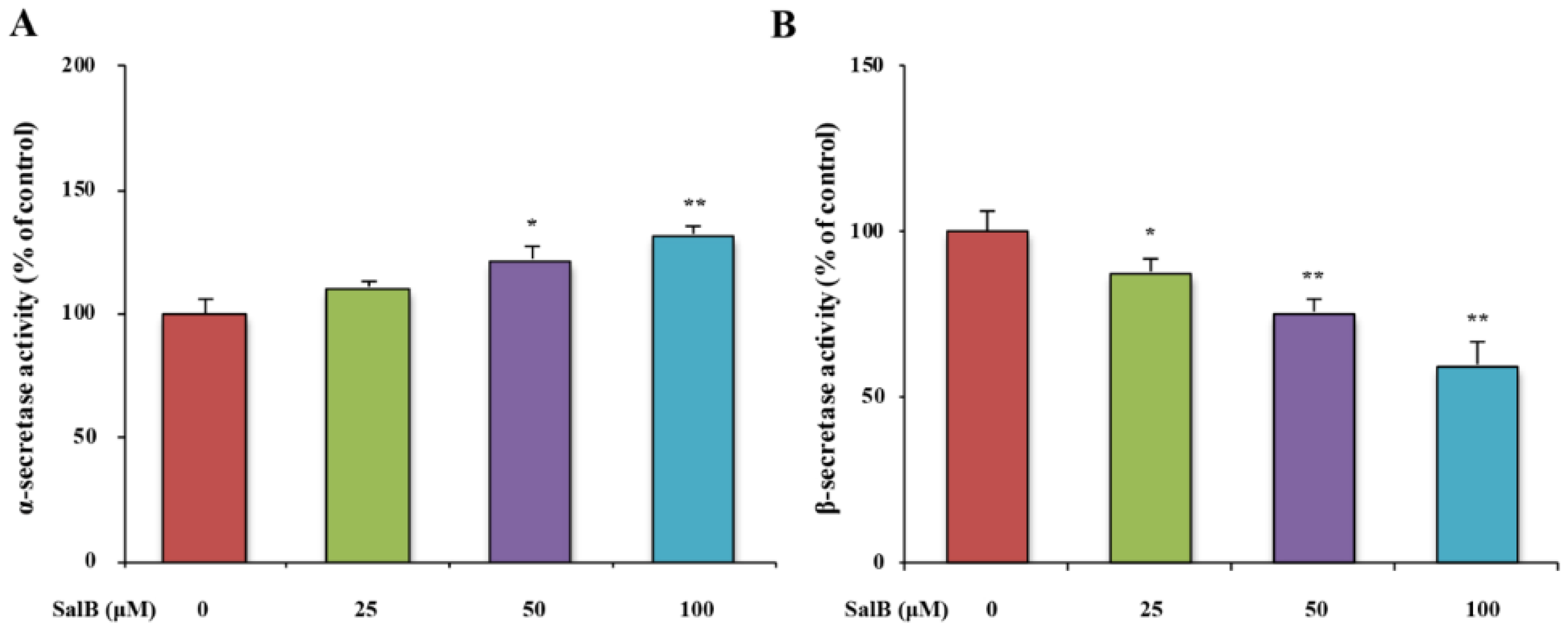
3.3. SalB Decreases the Protein Expression and the Activity of BACE1 in SH-SY5Y-APPsw Cells
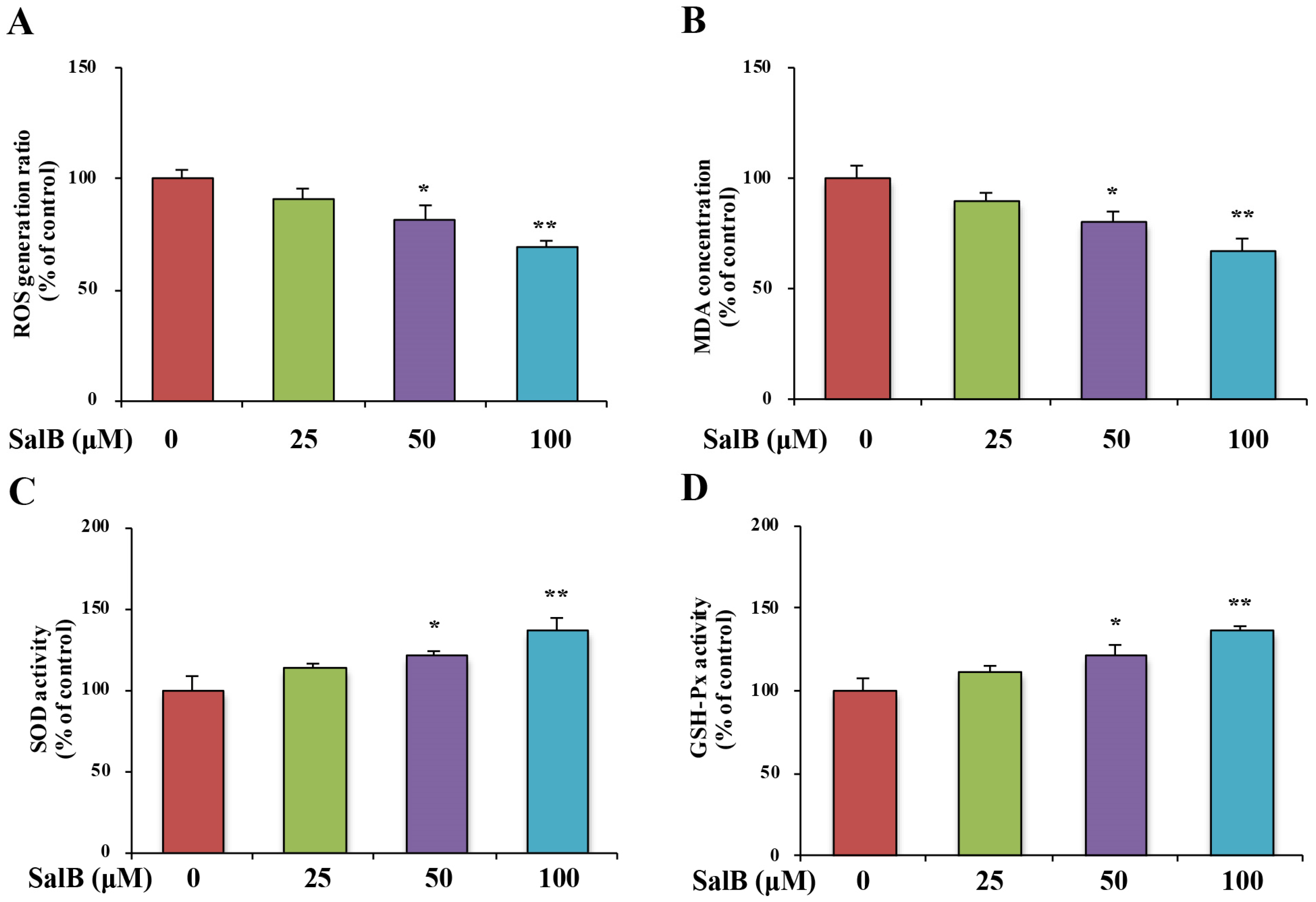
3.4. SalB Ameliorates Oxidative Stress and Inhibits GSK3β Activity in SH-SY5Y-APPsw Cells
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Qian, M.-C.; Liu, J.; Yao, J.-S.; Wang, W.-M.; Yang, J.-H.; Wei, L.-L.; Shen, Y.-D.; Chen, W. Caspase-8 mediates amyloid-β-induced apoptosis in differentiated pc12 cells. J. Mol. Neurosci. 2015, 56, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Billings, L.M.; Oddo, S.; Green, K.N.; McGaugh, J.L.; LaFerla, F.M. Intraneuronal aβ causes the onset of early alzheimer’s disease-related cognitive deficits in transgenic mice. Neuron 2005, 45, 675–688. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-H.; Giunta, B.; Zhou, H.-D.; Tan, J.; Wang, Y.-J. Opinion immunotherapy for alzheimer disease-the challenge of adverse effects. Nat. Rev. Neurol. 2012, 8, 465–469. [Google Scholar] [PubMed]
- Zhang, Y.; Li, X.; Wang, Z. Diversity evaluation of salvia miltiorrhiza using issr markers. Biochem. Genet. 2013, 51, 707–721. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Peng, H.; Shen, Y.; Zhao, R.; Huang, L. The profiling of bioactive ingredients of differently aged salvia miltiorrhiza roots. Microsc. Res. Tech. 2013, 76, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.M.; Zuo, Z.; Chow, M.S.S. Danshen: An overview of its chemistry, pharmacology, pharmacokinetics, and clinical use. J. Clin. Pharmacol. 2005, 45, 1345–1359. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Lin, S.J.; Ku, H.H.; Shiao, M.S.; Lin, F.Y.; Chen, J.W.; Chen, Y.L. Salvianolic acid b attenuates vcam-1 and icam-1 expression in tnf-alpha-treated human aortic endothelial cells. J. Cell. Biochem. 2001, 82, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Wu, C.H.; Luo, M.H.; Huang, Y.J.; Wang, C.N.; Shiao, M.S.; Huang, Y.T. In vitro protective effects of salvianolic acid b on primary hepatocytes and hepatic stellate cells. J. Ethnopharmacol. 2006, 105, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.W.; Kim, D.H.; Jeon, S.J.; Park, S.J.; Kim, J.M.; Jung, J.M.; Lee, H.E.; Bae, S.G.; Oh, H.K.; Son, K.H.H.; et al. Neuroprotective effects of salvianolic acid b on an a beta(25-35) peptide-induced mouse model of alzheimer’s disease. Eur. J. Pharmacol. 2013, 704, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.K.; Zhang, J.T. Salvianolic acid b inhibits fibril formation and neurotoxicity of amyloid beta-protein in vitro. Acta Pharmacol. Sin. 2001, 22, 380–384. [Google Scholar] [PubMed]
- Kim, D.H.; Park, S.J.; Kim, J.M.; Jeon, S.J.; Cho, Y.W.; Son, K.H.; Lee, H.J.; Moon, J.H.; Cheong, J.H.; Ko, K.H.; et al. Cognitive dysfunctions induced by a cholinergic blockade and a beta(25-35) peptide are attenuated by salvianolic acid b. Neuropharmacology 2011, 61, 1432–1440. [Google Scholar] [CrossRef] [PubMed]
- Winkler, E.; Julius, A.; Steiner, H.; Langosch, D. Homodimerization protects the amyloid precursor protein c99 fragment from cleavage by gamma-secretase. Biochemistry 2015, 54, 6149–6152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-W.; Xu, H. Molecular and cellular mechanisms for alzheimer’s disease: Understanding app metabolism. Curr. Mol. Med. 2007, 7, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Vassar, R.; Kandalepas, P.C. The beta-secretase enzyme bace1 as a therapeutic target for alzheimer’s disease. Alzheimers Res. Ther. 2011, 3, 20. [Google Scholar] [CrossRef] [PubMed]
- Thinakaran, G.; Koo, E.H. Amyloid precursor protein trafficking, processing, and function. J. Biol. Chem. 2008, 283, 29615–29619. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, D.K.; Farlow, M.R.; Sambamurti, K.; Greig, N.H.; Giacobini, E.; Schneider, L.S. A critical analysis of new molecular targets and strategies for drug developments in alzheimer’s disease. Curr. Drug Targets 2003, 4, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Ly, P.T.T.; Wu, Y.; Zou, H.; Wang, R.; Zhou, W.; Kinoshita, A.; Zhang, M.; Yang, Y.; Cai, F.; Woodgett, J.; et al. Inhibition of gsk3 beta-mediated bace1 expression reduces alzheimer-associated phenotypes. J. Clin. Investig. 2013, 123, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, D.B.; Soria, J.P.; Tomasiewicz, H.G.; Wood, J.G. Phosphorylation of human tau protein by microtubule-associated kinases: Gsk3 beta and cdk5 are key participants. J. Neurosci. Res. 2000, 62, 463–472. [Google Scholar] [CrossRef]
- Kayed, R.; Head, E.; Thompson, J.L.; McIntire, T.M.; Milton, S.C.; Cotman, C.W.; Glabe, C.G. Common structure of soluble amyloid oligomers implies common mechanism of pathogenesis. Science 2003, 300, 486–489. [Google Scholar] [CrossRef] [PubMed]
- Borchelt, D.R.; Thinakaran, G.; Eckman, C.B.; Lee, M.K.; Davenport, F.; Ratovitsky, T.; Prada, C.M.; Kim, G.; Seekins, S.; Yager, D.; et al. Familial alzheimer’s disease-linked presenilin 1 variants elevate a beta 1-42/1-40 ratio in vitro and in vivo. Neuron 1996, 17, 1005–1013. [Google Scholar] [CrossRef]
- Moir, R.D.; Tanzi, R.E. Lrp-mediated clearance of abeta is inhibited by kpi-containing isoforms of app. Curr. Alzheimer Res. 2005, 2, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Mao, G.; Cui, M.-Z.; Kang, S.-C.; Lamb, B.; Wong, B.-S.; Sy, M.-S.; Xu, X. Effects of gamma-secretase cleavage-region mutations on app processing and a beta formation: Interpretation with sequential cleavage and alpha-helical model. J. Neurochem. 2008, 107, 722–733. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Yan, P.; Ma, X.; Liu, H.; Perez, R.; Zhu, A.; Gonzales, E.; Tripoli, D.L.; Czerniewski, L.; Ballabio, A.; et al. Neuronal-targeted tfeb accelerates lysosomal degradation of app, reducing a beta generation and amyloid plaque pathogenesis. J. Neurosci. 2015, 35, 12137–12151. [Google Scholar] [CrossRef] [PubMed]
- Devasagayam, T.P.A.; Tilak, J.C.; Boloor, K.K.; Sane, K.S.; Ghaskadbi, S.S.; Lele, R.D. Free radicals and antioxidants in human health: Current status and future prospects. J. Assoc. Physicians India 2004, 52, 794–804. [Google Scholar] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Perry, G.; Castellani, R.J.; Smith, M.A.; Harris, P.L.R.; Kubat, Z.; Ghanbari, K.; Jones, P.K.; Cordone, G.; Tabaton, M.; Wolozin, B.; et al. Oxidative damage in the olfactory system in alzheimer’s disease. Acta Neuropathol. 2003, 106, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Varadarajan, S.; Kanski, J.; Aksenova, M.; Lauderback, C.; Butterfield, D.A. Different mechanisms of oxidative stress and neurotoxicity for alzheimer’s a beta(1-42) and a beta(25-35). J. Am. Chem. Soc. 2001, 123, 5625–5631. [Google Scholar] [CrossRef] [PubMed]
- De Felice, F.G.; Velasco, P.T.; Lambert, M.P.; Viola, K.; Fernandez, S.J.; Ferreira, S.T.; Klein, W.L. A beta oligomers induce neuronal oxidative stress through an N-methyl-d-aspartate receptor-dependent mechanism that is blocked by the alzheimer drug memantine. J. Biol. Chem. 2007, 282, 11590–11601. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.D.; Atwood, C.S.; Hartshorn, M.A.; Multhaup, G.; Goldstein, L.E.; Scarpa, R.C.; Cuajungco, M.P.; Gray, D.N.; Lim, J.; Moir, R.D.; et al. The a beta peptide of alzheimer’s disease directly produces hydrogen peroxide through metal ion reduction. Biochemistry 1999, 38, 7609–7616. [Google Scholar] [CrossRef] [PubMed]
- Guglielmotto, M.; Giliberto, L.; Tamagno, E.; Tabaton, M. Oxidative stress mediates the pathogenic effect of different alzheimer’s disease risk factors. Front. Aging Neurosci. 2010, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Borghi, R.; Patriarca, S.; Traverso, N.; Piccini, A.; Storace, D.; Garuti, A.; Cirmena, G.; Odetti, P.; Tabaton, M. The increased activity of bace1 correlates with oxidative stress in alzheimer’s disease. Neurobiol. Aging 2007, 28, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Zhou, W.; Fung, V.; Christensen, M.A.; Qing, H.; Sun, X.; Song, W. Oxidative stress potentiates bace1 gene expression and a beta generation. J. Neural Transm. 2005, 112, 455–469. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, S.; Sun, J.; Shi, Y.; Le, G. Salvianolic acid b relieves oxidative stress in glucose absorption and utilization of mice fed high-sugar diet. Trop. J. Pharm. Res. 2014, 13, 369–375. [Google Scholar] [CrossRef]
- Wu, H.-L.; Li, Y.-H.; Lin, Y.-H.; Wang, R.; Li, Y.-B.; Tie, L.; Song, Q.-L.; Guo, D.-A.; Yu, H.-M.; Li, X.-J. Salvianolic acid b protects human endothelial cells from oxidative stress damage: A possible protective role of glucose-regulated protein 78 induction. Cardiovasc. Res. 2009, 81, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Jope, R.S.; Yuskaitis, C.J.; Beurel, E. Glycogen synthase kinase-3 (gsk3): Inflammation, diseases, and therapeutics. Neurochem. Res. 2007, 32, 577–595. [Google Scholar] [CrossRef] [PubMed]







© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Huang, D.; Zhang, M.-H.; Zhang, W.-S.; Tang, Y.-X.; Shi, Z.-X.; Deng, L.; Zhou, D.-H.; Lu, X.-Y. Salvianolic Acid B Inhibits Aβ Generation by Modulating BACE1 Activity in SH-SY5Y-APPsw Cells. Nutrients 2016, 8, 333. https://0-doi-org.brum.beds.ac.uk/10.3390/nu8060333
Tang Y, Huang D, Zhang M-H, Zhang W-S, Tang Y-X, Shi Z-X, Deng L, Zhou D-H, Lu X-Y. Salvianolic Acid B Inhibits Aβ Generation by Modulating BACE1 Activity in SH-SY5Y-APPsw Cells. Nutrients. 2016; 8(6):333. https://0-doi-org.brum.beds.ac.uk/10.3390/nu8060333
Chicago/Turabian StyleTang, Ying, Dan Huang, Mei-Hua Zhang, Wen-Sheng Zhang, Yu-Xin Tang, Zheng-Xiang Shi, Li Deng, Dai-Han Zhou, and Xin-Yi Lu. 2016. "Salvianolic Acid B Inhibits Aβ Generation by Modulating BACE1 Activity in SH-SY5Y-APPsw Cells" Nutrients 8, no. 6: 333. https://0-doi-org.brum.beds.ac.uk/10.3390/nu8060333