Habitat Affects the Chemical Profile, Allelopathy, and Antioxidant Properties of Essential Oils and Phenolic Enriched Extracts of the Invasive Plant Heliotropium Curassavicum

 , ,
, , 

Abstract
:1. Introduction
2. Results and Discussion
2.1. Soil Physico-Chemical Parameters
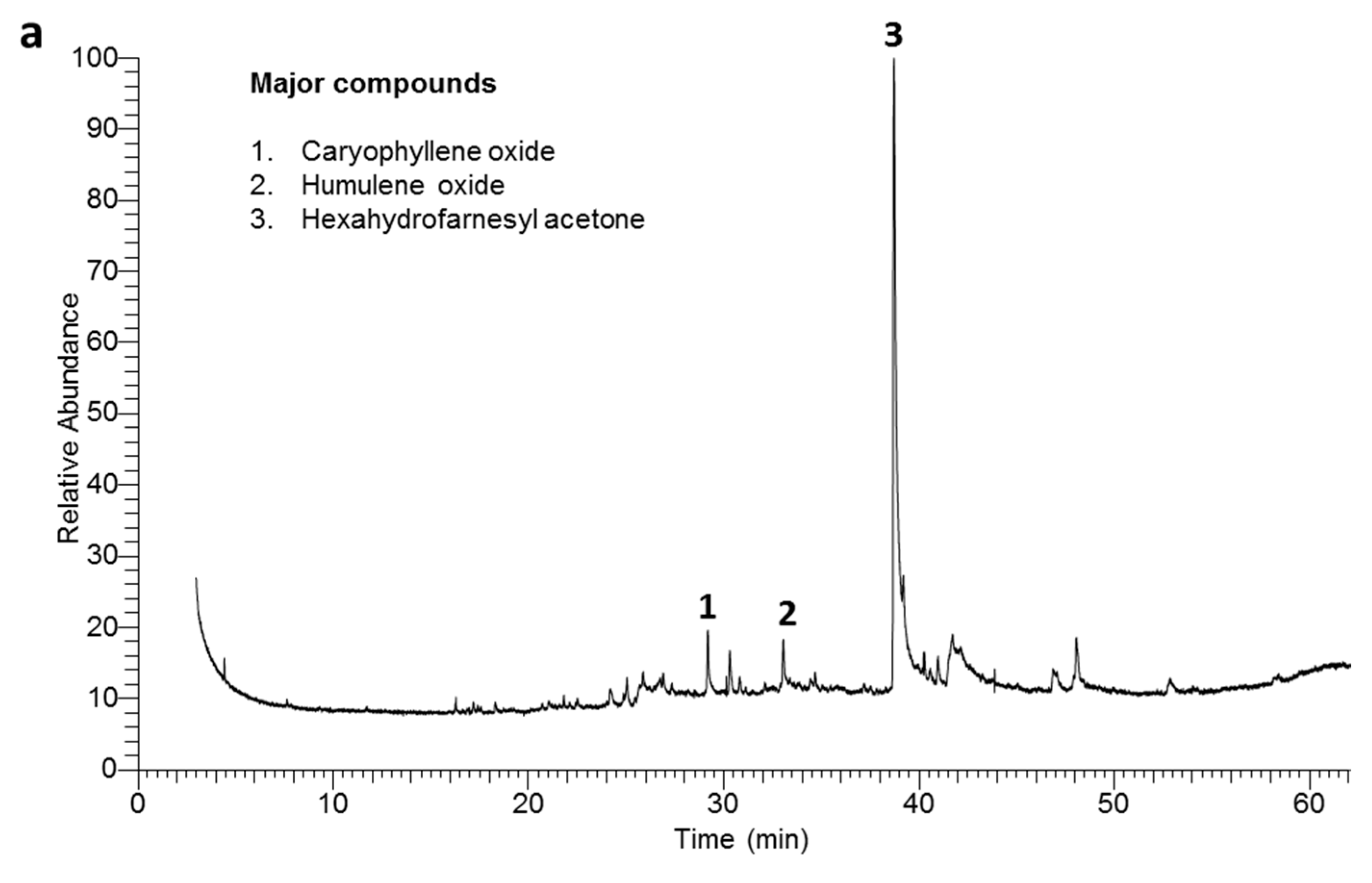
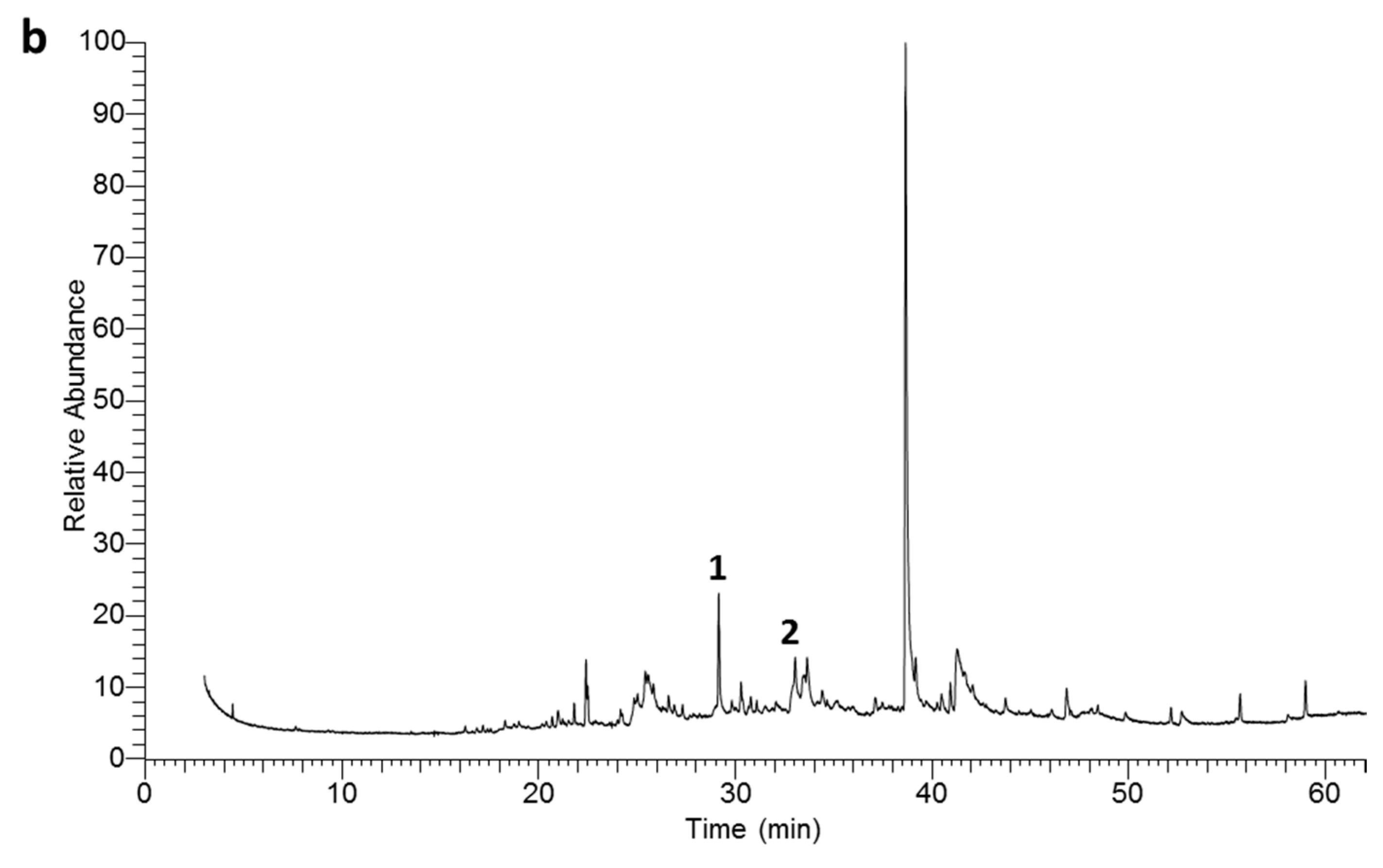
2.2. Chemical Composition of the EOs
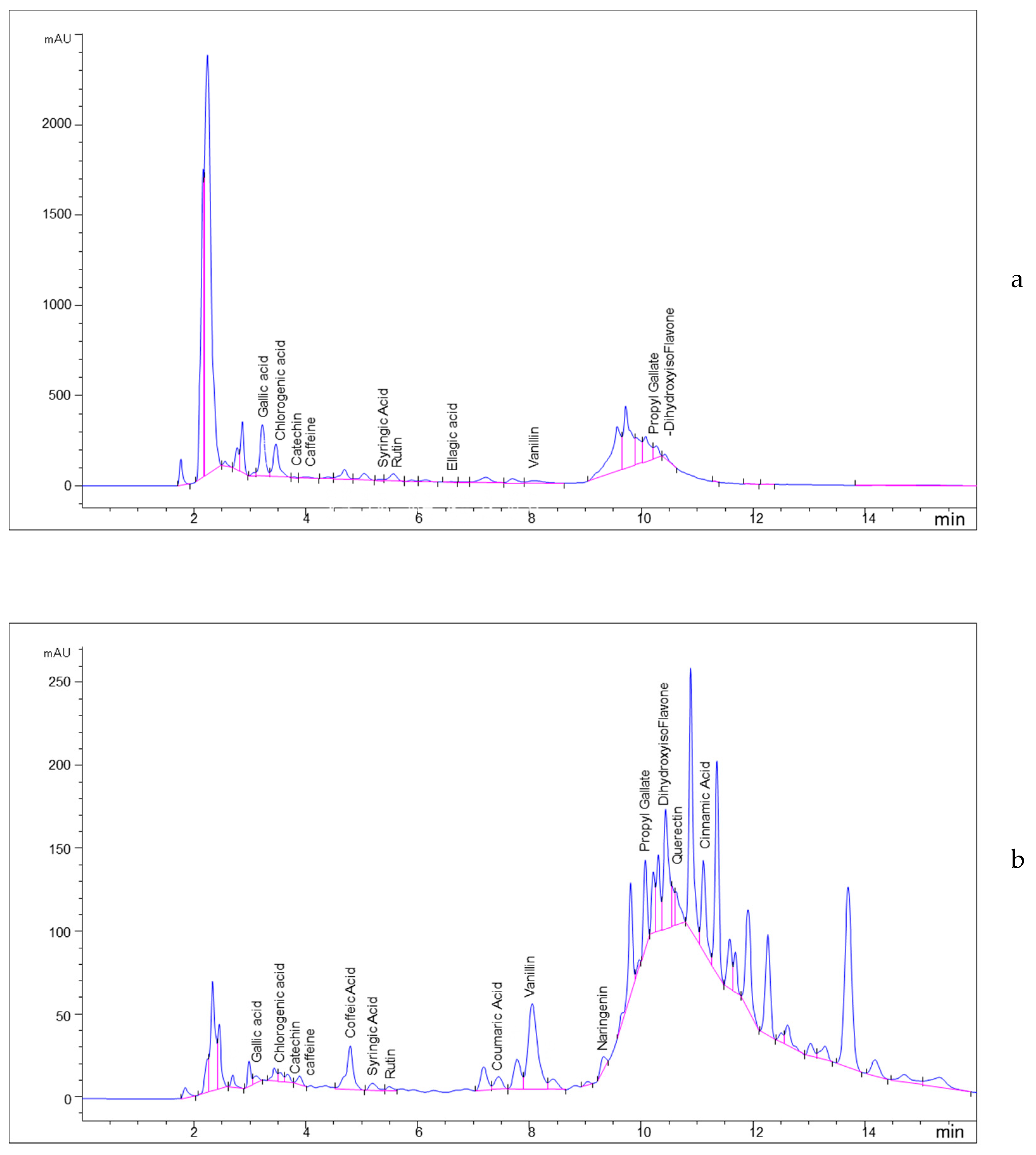
2.3. Phenolic Profile of 70% MeOH Extract
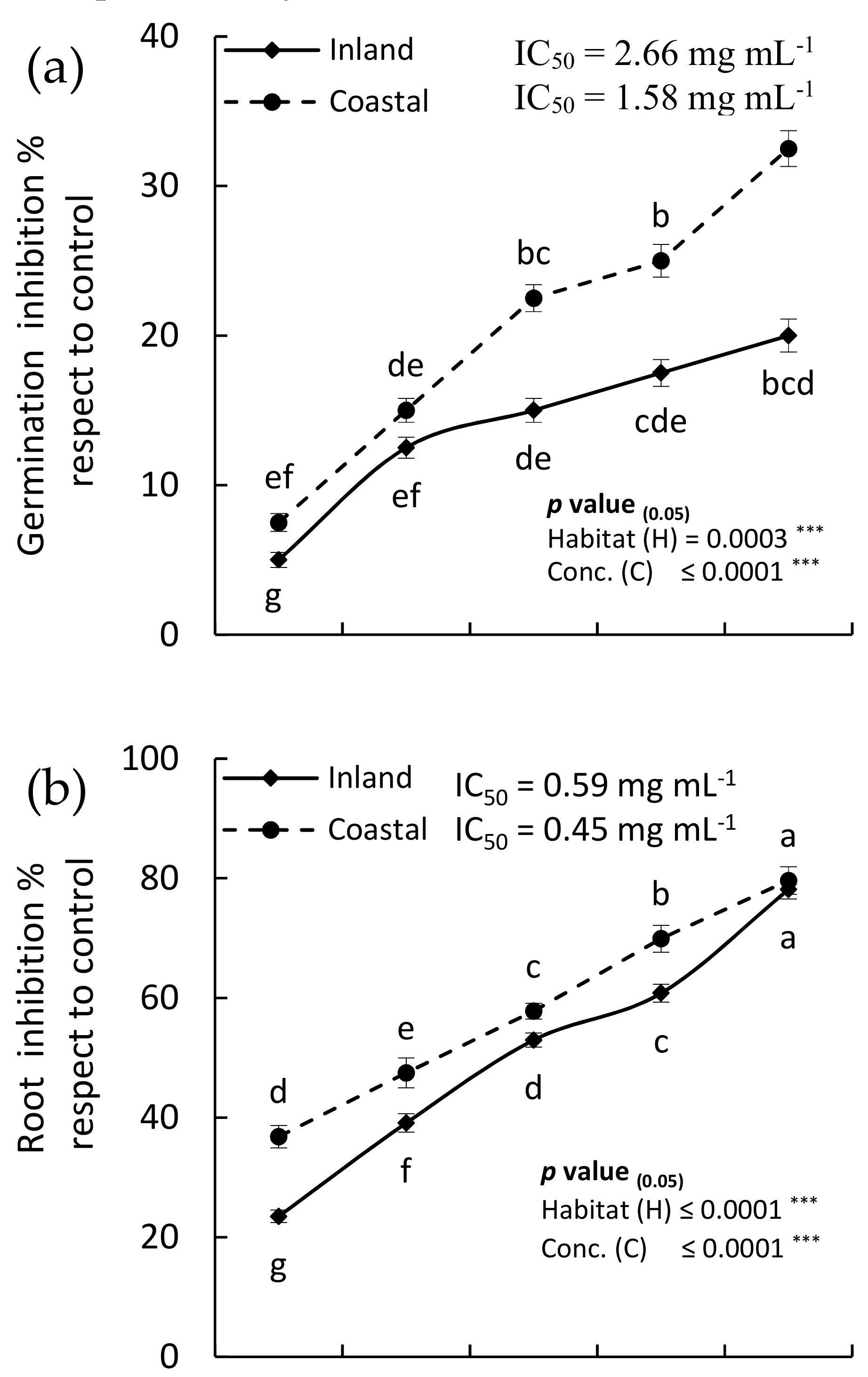
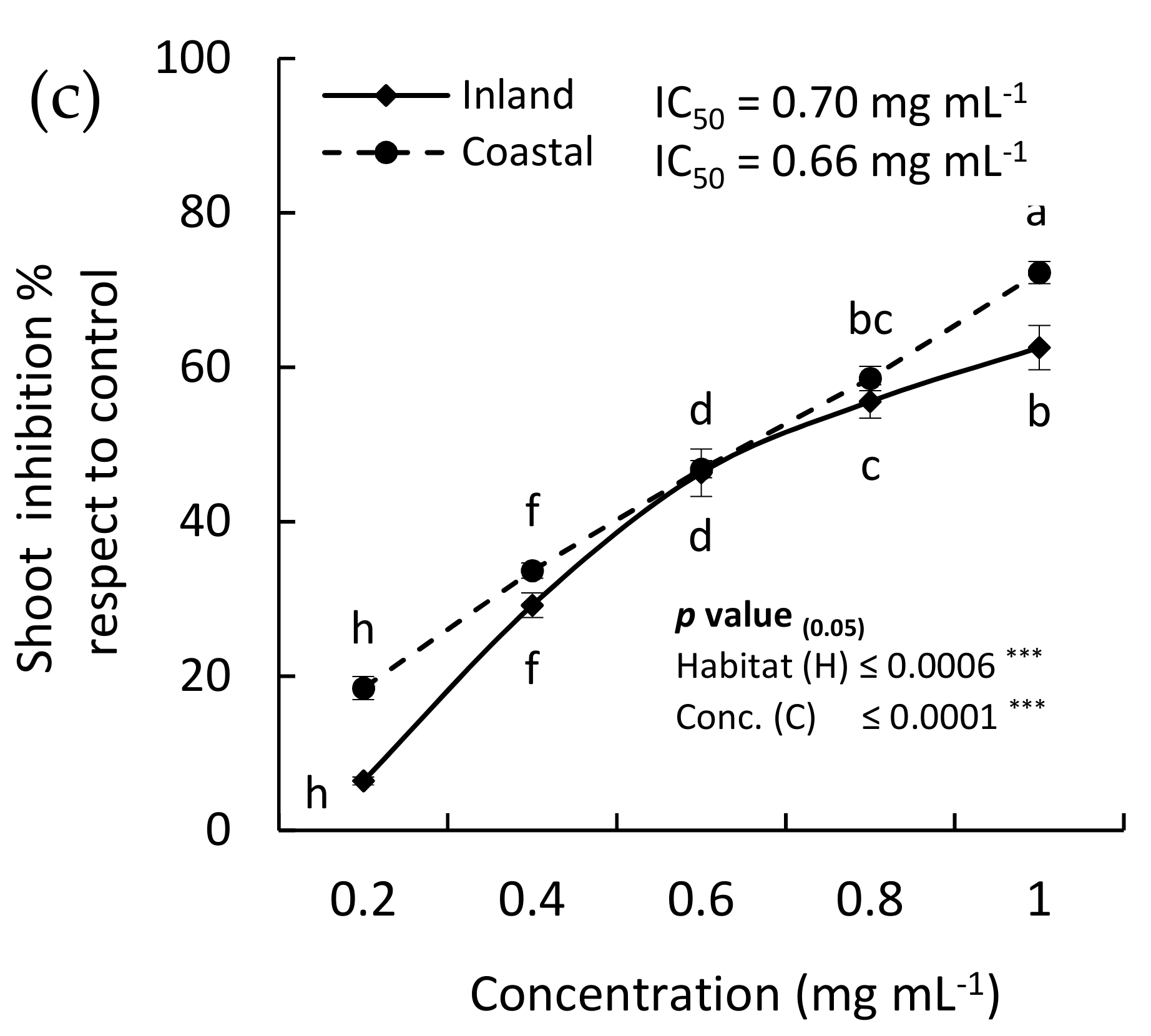
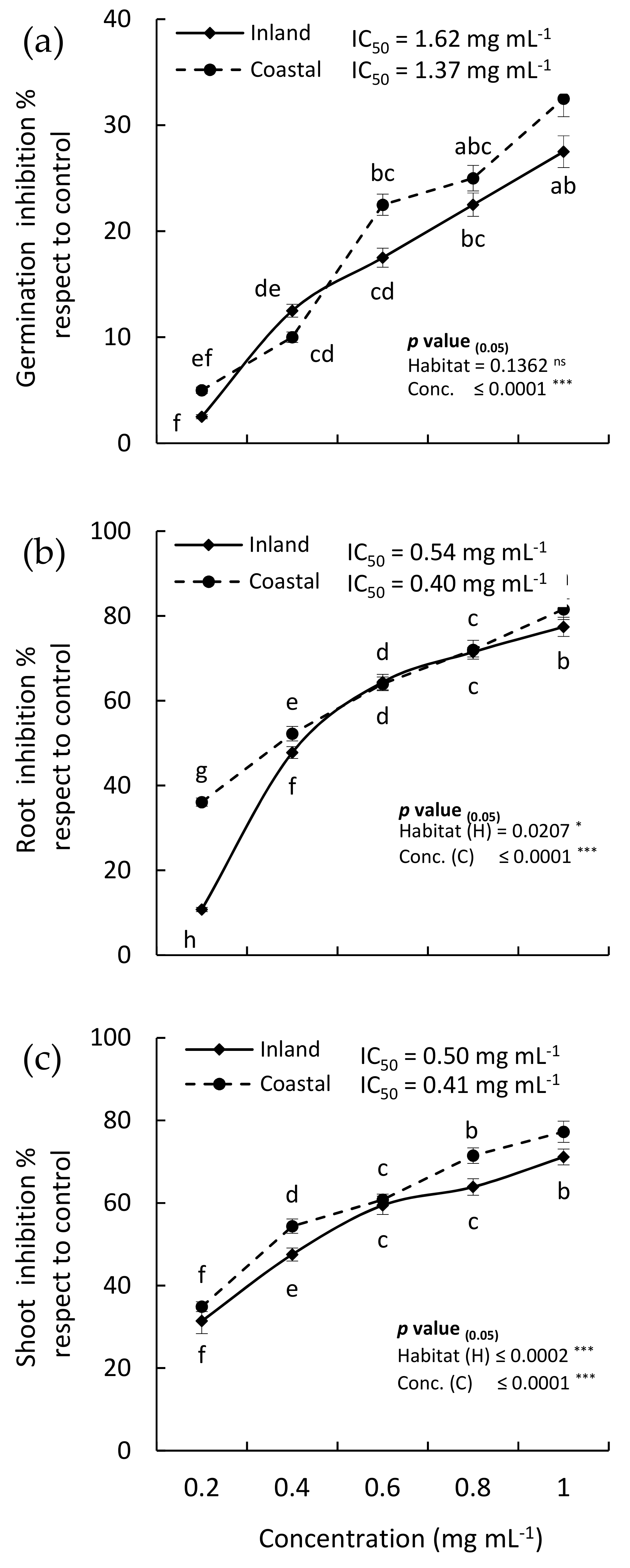
2.4. Allelopathic Activity
2.4.1. Allelopathic Effect of the EOs
2.4.2. Allelopathic Effect of the MeOH Extract
2.5. Antioxidant activity
3. Material and Methods
3.1. Plant Material
3.2. Soil Analyses
3.3. Extraction and Identification of EOs
3.4. Preparation of MeOH Extract and HPLC Analysis
3.5. Allelopathic Activity
3.6. Antioxidant Activity
3.6.1. DPPH Radical Scavenging Activity
3.6.2. ABTS-Free Radical Scavenging Activity
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hussain, K.; Shahazad, A.; Zia-ul-Hussnain, S. An ethnobotanical survey of important wild medicinal plants of Hattar district Haripur, Pakistan. Ethnobot. Leafl. 2008, 2008, 5. [Google Scholar]
- David, B.; Wolfender, J.-L.; Dias, D.A. The pharmaceutical industry and natural products: Historical status and new trends. Phytochem. Rev. 2015, 14, 299–315. [Google Scholar] [CrossRef]
- Elshamy, A.; Abd-ElGawad, A.M.; El-Amier, Y.A.; El Gendy, A.; Al-Rowaily, S. Interspecific variation, antioxidant and allelopathic activity of the essential oil from three Launaea species growing naturally in heterogeneous habitats in Egypt. Flavour Fragr. J. 2019, 34, 316–328. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Ademiluyi, A.O. Biological activities of essential oils: From plant chemoecology to traditional healing systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef]
- Abd El-Gawad, A.M. Chemical constituents, antioxidant and potential allelopathic effect of the essential oil from the aerial parts of Cullen plicata. Ind. Crops Prod. 2016, 80, 36–41. [Google Scholar] [CrossRef]
- Kahraman, C.; Topcu, G.; Bedir, E.; Tatli, I.I.; Ekizoglu, M.; Akdemir, Z.S. Phytochemical screening and evaluation of the antimicrobial and antioxidant activities of Ferula caspica M. Bieb. extracts. Saudi Pharm. J. 2019, 27, 525–531. [Google Scholar] [CrossRef]
- Shoaib, M.; Shah, I.; Ali, N.; Adhikari, A.; Tahir, M.N.; Shah, S.W.A.; Ishtiaq, S.; Khan, J.; Khan, S.; Umer, M.N. Sesquiterpene lactone! A promising antioxidant, anticancer and moderate antinociceptive agent from Artemisia macrocephala jacquem. BMC Complement. Altern. Med. 2017, 17, 27. [Google Scholar] [CrossRef]
- Tofighi, Z.; Alipour, F.; Yassa, N.; Hadjiakhoondi, A.; Hadavinia, H.; Goodarzy, S.; Golestani, R. Chemical composition and antioxidant activity of Otostegia persica essential oil from Iran. Int. J. Essent. Oil Ther. 2009, 3, 45–48. [Google Scholar]
- Sakr, A.A.; Ghaly, M.F.; Abdel-Haliem, M.E.-S.F. The efficacy of specific essential oils on yeasts isolated from the royal tomb paintings at Tanis, Egypt. Int. J. Conserv. Sci. 2012, 3, 87–92. [Google Scholar]
- Rotolo, V.; De Caro, M.; Giordano, A.; Palla, F. Solunto archaeological park in Sicily: Life under mosaic tesserae. Flora Medit 2018, 28, 233–245. [Google Scholar]
- Walentowska, J.; Foksowicz-Flaczyk, J. Thyme essential oil for antimicrobial protection of natural textiles. Int. Biodeter. Biodegr. 2013, 84, 407–411. [Google Scholar] [CrossRef]
- Abd El-Gawad, A.M.; El Gendy, A.G.; Elshamy, A.I.; Omer, E.A. Chemical composition of the essential oil of Trianthema portulacastrum L. Aerial parts and potential antimicrobial and phytotoxic activities of its extract. J. Essent. Oil Bear. Plants 2016, 19, 1684–1692. [Google Scholar] [CrossRef]
- Bruin, J.; Sabelis, M.W.; Dicke, M. Do plants tap SOS signals from their infested neighbours? Trends Ecol. Evol. 1995, 10, 167–170. [Google Scholar] [CrossRef] [Green Version]
- Miresmailli, S.; Isman, M.B. Botanical insecticides inspired by plant—Herbivore chemical interactions. Trends Plant Sci. 2014, 19, 29–35. [Google Scholar] [CrossRef]
- Cseke, L.J.; Kaufman, P.B.; Kirakosyan, A. The biology of essential oils in the pollination of flowers. Nat. Prod. Commun. 2007, 2, 1317–1336. [Google Scholar] [CrossRef]
- Barney, J.N.; Tekiela, D.R.; Dollete, E.S.; Tomasek, B.J. What is the “real” impact of invasive plant species? Front. Ecol. Environ. 2013, 11, 322–329. [Google Scholar] [CrossRef]
- Zheng, Y.L.; Feng, Y.L.; Zhang, L.K.; Callaway, R.M.; Valiente-Banuet, A.; Luo, D.Q.; Liao, Z.Y.; Lei, Y.B.; Barclay, G.F.; Silva-Pereyra, C. Integrating novel chemical weapons and evolutionarily increased competitive ability in success of a tropical invader. New Phytol. 2015, 205, 1350–1359. [Google Scholar] [CrossRef]
- Einhellig, F.A. Allelopathy: Current Status and Future Goals; ACS Publications: Washington, DC, USA, 1995. [Google Scholar]
- Al-Shehbaz, I.A. The genera of Boraginaceae in the southeastern United States. J. Arnold. Arbor. 1991, 1, 1–169. [Google Scholar]
- Hegazy, A.; Mussa, S.; Farrag, H. Invasive plant communities in the Nile Delta coast. Glob. J. Environ. Res. 2008, 2, 53–61. [Google Scholar]
- Hegazy, A.K. Trade-off between sexual and vegetative reproduction of the weedy Heliotropium curassavicum. J. Arid Environ. 1994, 27, 209–220. [Google Scholar] [CrossRef]
- Abd-ElGawad, A.M.; Elshamy, A.I.; El-Amier, Y.A.; El Gendy, A.E.-N.G.; Al-Barati, S.A.; Dar, B.A.; Al-Rowaily, S.L.; Assaeed, A.M. Chemical composition variations, allelopathic, and antioxidant activities of Symphyotrichum squamatum (Spreng.) Nesom essential oils growing in heterogeneous habitats. Arab. J. Chem. 2019, in press. [Google Scholar] [CrossRef]
- Souza, J.S.N.; Machado, L.L.; Pessoa, O.D.; Braz-Filho, R.; Overk, C.R.; Yao, P.; Cordell, G.A.; Lemos, T.L. Pyrrolizidine alkaloids from Heliotropium indicum. J. Braz. Chem. Soc. 2005, 16, 1410–1414. [Google Scholar] [CrossRef]
- Yeo, D.; Attioua, B.; Lehalle, C.; Kossi, M.; N’guessan, J.D.; Djaman, A.J.; Lobstein, A.; Frossard, N. Isolation of wound healing compounds from Heliotropium indicum. J. Appl. Pharm. Sci. 2011, 1, 102–106. [Google Scholar]
- Singh, B.; Sahu, P.M.; Sharma, R.A. Flavonoids from Heliotropium subulatum exudate and their evaluation for antioxidant, antineoplastic and cytotoxic activities II. Cytotechnology 2017, 69, 103–115. [Google Scholar] [CrossRef]
- Singh, B.; Sahu, P.; Sharma, R. In vitro and in vivo evaluation of flavonoids from Heliotropium ellipticum exudate for antioxidant, antineoplastic and cytotoxic activities ii. Indian J. Pharm. Sci. 2018, 79, 939–947. [Google Scholar] [CrossRef]
- Jain, S.; Singh, B.; Jain, R. Antimicrobial activity of triterpenoids from Heliotropium ellipticum. Fitoterapia 2001, 72, 666–668. [Google Scholar] [CrossRef]
- Modak, B.; Rojas, M.; Torres, R. Chemical analysis of the resinous exudate isolated from Heliotropium taltalense and evaluation of the antioxidant activity of the phenolics components and the resin in homogeneous and heterogeneous systems. Molecules 2009, 14, 1980–1989. [Google Scholar] [CrossRef]
- Khan, H.; Khan, M.A.; Gul, F.; Hussain, S.; Ashraf, N. Anti-inflammatory activity of Heliotropium strigosum in animal models. Toxicol. Ind. Health 2015, 31, 1281–1287. [Google Scholar] [CrossRef]
- Cordovilla, M.P.; Bueno, M.; Aparicio, C.; Urrestarazu, M. Effects of salinity and the interaction between Thymus vulgaris and Lavandula angustifolia on growth, ethylene production and essential oil contents. J. Plant Nutr. 2014, 37, 875–888. [Google Scholar] [CrossRef]
- Abd El-Gawad, A.M.; Elshamy, A.I.; El Gendy, A.E.-N.; Gaara, A.; Assaeed, A.M. Volatiles profiling, allelopathic activity, and antioxidant potentiality of Xanthium strumarium leaves essential oil from Egypt: Evidence from chemometrics analysis. Molecules 2019, 24, 584. [Google Scholar] [CrossRef]
- Ogunbinu, A.O.; Flamini, G.; Cioni, P.L.; Adebayo, M.A.; Ogunwande, I.A. Constituents of Cajanus cajan (L.) Millsp., Moringa oleifera Lam., Heliotropium indicum L. and Bidens pilosa L. from Nigeria. Nat. Prod. Commun. 2009, 4, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Abd-ElGawad, A.M.; Elshamy, A.; El Gendy, A.E.-N.; Al-Rowaily, S.L.; Assaeed, A.M. Preponderance of oxygenated sesquiterpenes and diterpenes in the volatile oil constituents of Lactuca serriola L. revealed antioxidant and allelopathic activity. Chem Biodivers. 2019, 16, e1900278. [Google Scholar] [CrossRef] [PubMed]
- Machan, T.; Korth, J.; Liawruangrath, B.; Liawruangrath, S.; Pyne, S.G. Composition and antituberculosis activity of the volatile oil of Heliotropium indicum Linn. growing in Phitsanulok, Thailand. Flavour Fragr. J. 2006, 21, 265–267. [Google Scholar] [CrossRef]
- Saeedi, M.; Morteza-Semnani, K. Chemical composition and antimicrobial activity of the essential oil of Heliotropium europaeum. Chem. Nat. Compd. 2009, 45, 98–99. [Google Scholar] [CrossRef]
- Abd El-Gawad, A.M.; El-Amier, Y.A. Allelopathy and potential impact of invasive Acacia saligna (Labill.) Wendl. on plant diversity in the Nile Delta coast of Egypt. Int. J. Environ. Res. 2015, 9, 923–932. [Google Scholar]
- Nour, V.; Trandafir, I.; Cosmulescu, S. HPLC determination of phenolic acids, flavonoids and juglone in walnut leaves. J. Chromatogr. Sci. 2012, 51, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Sotubo, S.E.; Lawal, O.A.; Osunsami, A.A.; Ogunwande, I.A. Constituents and insecticidal activity of Deinbollia pinnata essential oil. Nat. Prod. Commun. 2016, 11, 1889–1890. [Google Scholar] [CrossRef]
- Ghori, M.K.; Ghaffari, M.A.; Hussain, S.N.; Manzoor, M.; Aziz, M.; Sarwer, W. Ethnopharmacological, phytochemical and pharmacognostic potential of genus Heliotropium L. Turk. J. Pharm. Sci. 2016, 13, 143–168. [Google Scholar] [CrossRef]
- Bistgani, Z.E.; Hashemi, M.; DaCosta, M.; Craker, L.; Maggi, F.; Morshedloo, M.R. Effect of salinity stress on the physiological characteristics, phenolic compounds and antioxidant activity of Thymus vulgaris L. and Thymus daenensis Celak. Ind. Crops Prod. 2019, 135, 311–320. [Google Scholar] [CrossRef]
- Salem, N.; Msaada, K.; Dhifi, W.; Limam, F.; Marzouk, B. Effect of salinity on plant growth and biological activities of Carthamus tinctorius L. extracts at two flowering stages. Acta Physiol. Plant. 2014, 36, 433–445. [Google Scholar] [CrossRef]
- Abd El-Gawad, A.M. Ecology and allelopathic control of Brassica tournefortii in reclaimed areas of the Nile Delta, Egypt. Turk. J. Bot. 2014, 38, 347–357. [Google Scholar] [CrossRef]
- Abd El-Gawad, A.M.; El-Amier, Y.A.; Bonanomi, G. Allelopathic activity and chemical composition of Rhynchosia minima (L.) DC. essential oil from Egypt. Chem. Biodivers. 2018, 15, e1700438. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Gawad, A.M.; Shehata, H.S. Ecology and development of Mesembryanthemum crystallinum L. in the Deltaic Mediterranean coast of Egypt. Egypt. J. Basic Appl. Sci. 2014, 1, 29–37. [Google Scholar] [CrossRef]
- Radulović, N.; Stojanović, G.; Palić, R. Composition and antimicrobial activity of Equisetum arvense L. essential oil. Phytother. Res. 2006, 20, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Xiangwei, Z.; Xiaodong, W.; Peng, N.; Yang, Z.; JiaKuan, C. Chemical composition and antimicrobial activity of the essential oil of Sagittaria trifolia. Chem. Nat. Compd. 2006, 42, 520–522. [Google Scholar] [CrossRef]
- Venditti, A.; Frezza, C.; Bianco, A.; Serafini, M.; Cianfaglione, K.; Nagy, D.U.; Iannarelli, R.; Caprioli, G.; Maggi, F. Polar constituents, essential oil and antioxidant activity of marsh woundwort (Stachys palustris L.). Chem. Biodivers. 2017, 14, e1600401. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, K.; Kalemba, D.; Komsta, Ł.; Nowak, R. Comparison of the essential oil composition of selected Impatiens species and its antioxidant activities. Molecules 2016, 21, 1162. [Google Scholar] [CrossRef]
- Benmeddour, T.; Laouer, H.; Flamini, G.; Akkal, S. Chemical Composition of essential oil of Limonium bonduellei. Chem. Nat. Compd. 2018, 54, 188–190. [Google Scholar] [CrossRef]
- Balogun, O.S.; Ajayi, O.S.; Adeleke, A.J. Hexahydrofarnesyl acetone-rich extractives from Hildegardia barteri. J. Herbs Spices Med. Plants 2017, 23, 393–400. [Google Scholar] [CrossRef]
- Li, Z.-H.; Wang, Q.; Ruan, X.; Pan, C.-D.; Jiang, D.-A. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef]
- Inderjit. Plant phenolics in allelopathy. Bot. Rev. 1996, 62, 186–202. [Google Scholar] [CrossRef]
- Chou, C.-H.; Leu, L.-L. Allelopathic substances and interactions of Delonix regia (Boj) Raf. J. Chem. Ecol. 1992, 18, 2285–2303. [Google Scholar] [CrossRef] [PubMed]
- Chon, S.U.; Kim, J.D. Biological activity and quantification of suspected allelochemicals from alfalfa plant parts. J. Agron. Crop Sci. 2002, 188, 281–285. [Google Scholar] [CrossRef]
- Mallik, M.; Puchala, R.; Grosz, F. A growth inhibitory factor from lambsquarters (Chenopodium album). J. Chem. Ecol. 1994, 20, 957–967. [Google Scholar] [CrossRef] [PubMed]
- El-Shora, H.M.; Abd El-Gawad, A.M. Evaluation of allelopathic potential of root extract of Rumex dentatus and allelochemicals on Cicer arietinum. J. Stress Physiol. Biochem. 2014, 10, 167–180. [Google Scholar]
- Alam, S.; Shaikh, A. Influence of leaf extract of nettle leaf goosefoot (Chenopodium murale L.) and NaCl salinity on germination and seedling growth of rice (Oryza sativa L.). Pak. J. Bot. 2007, 39, 1695–1699. [Google Scholar]
- Al-Johani, N.S.; Aytah, A.A.; Boutraa, T. Allelopathic impact of two weeds, Chenopodium murale and Malva parviflora on growth and photosynthesis of barley (Hordeum vulgare L.). Pak. J. Bot. 2012, 44, 1865–1872. [Google Scholar]
- Majeed, A.; Chaudhry, Z.; Muhammad, Z. Allelopathic assessment of fresh aqueous extracts of Chenopodium album L. for growth and yield of wheat (Triticum aestivum L.). Pak. J. Bot. 2012, 44, 165–167. [Google Scholar]
- Batish, D.; Lavanya, K.; Pal Singh, H.; Kohli, R. Root-mediated allelopathic interference of nettle-leaved goosefoot (Chenopodium murale) on wheat (Triticum aestivum). J. Agron. Crop Sci. 2007, 193, 37–44. [Google Scholar] [CrossRef]
- Agarwal, S.K. Pesticide Pollution; APH Publishing: New Delhi, India, 2009; Volume 3. [Google Scholar]
- Alam, M.A.; Juraimi, A.; Rafii, M.; Hamid, A.; Aslani, F.; Alam, M. Effects of salinity and salinity-induced augmented bioactive compounds in purslane (Portulaca oleracea L.) for possible economical use. Food Chem. 2015, 169, 439–447. [Google Scholar] [CrossRef]
- Vafadar Shoshtari, Z.; Rahimmalek, M.; Sabzalian, M.R.; Hosseini, H. Essential oil and bioactive compounds variation in myrtle (Myrtus communis L.) as affected by seasonal variation and salt stress. Chem. Biodivers. 2017, 14, e1600365. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.M.; Albu, C.; Matar, S.A.; Litescu, S.-C.; Al Jaber, H.I.; Abualraghib, A.S.; Afifi, F.U. Biological activities of the hydro-alchoholic and aqueous extracts of Achillea biebersteinii Afan.(Asteraceae) grown in Jordan. Afr. J. Pharm. Pharm. 2013, 7, 1686–1694. [Google Scholar] [CrossRef]
- El-Kashak, W.A.; Elshamy, A.I.; Mohamed, T.A.; El Gendy, A.E.-N.G.; Saleh, I.A.; Umeyama, A. Rumpictuside A: Unusual 9, 10-anthraquinone glucoside from Rumex pictus Forssk. Carbohydr. Res. 2017, 448, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Boulos, L. Flora of Egypt: Azollaceae-Oxalidaceae; Al Hadara Publishing: Cairo, Egypt, 1999; Volume 1. [Google Scholar]
- Piper, C.S. Soil and Plant Analysis; Interscience Publishers, Inc.: New York, NY, USA, 1947. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis; Prentic Hall Pvt. Ltd.: New Delhi, India, 1973. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Waste Water, 19th ed.; American Public Health Association, American Water Work Association, Water Pollution Control Federation: Washington, DC, USA, 1998; Volume 2. [Google Scholar]
- Miguel, M.G. Antioxidant activity of medicinal and aromatic plants. Flavour Fragr. J. 2010, 25, 291–312. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Habitat | p-value | |
|---|---|---|---|
| Coastal | Inland | ||
| Sand (%) | 94.16 ± 2.05 | 67.39 ± 2.37 | 0.008 * |
| Silt (%) | 4.20 ± 1.79 | 19.56 ± 2.63 | 0.026 * |
| Clay (%) | 1.65 ± 0.68 | 13.06 ± 1.62 | 0.010 * |
| pH | 7.37 ± 0.49 | 7.17 ± 0.12 | 0.591 |
| OC (g kg−1) | 0.63 ± 0.09 | 1.30 ± 0.13 | 0.032 * |
| EC (mS cm−1) | 1.68 ± 0.21 | 0.57 ± 0.01 | 0.012 * |
| TDP (g kg−1) | 0.46 ± 0.10 | 2.11 ± 0.35 | 0.022 * |
| TN (g kg−1) | 0.33 ± 0.05 | 3.58 ± 1.04 | 0.033 * |
| No | Rt | KIlit [a] | KIexp [b] | Compound | Concentration % [c] | Identification [d] | |
|---|---|---|---|---|---|---|---|
| Coastal | Inland | ||||||
| Monoterpene hydrocarbons | |||||||
| 1 | 4.45 | 936 | 936 | α-Pinene | 0.79 ± 0.03 | 0.4 ± 0.02 | MS, KI |
| 2 | 18.99 | 1375 | 1374 | β-Elemene | -- | 0.32 | MS, KI |
| Oxygenated Monoterpenes | |||||||
| 3 | 18.31 | 1173 | 1174 | Menthol | 0.86 ± 0.03 | 0.42 ± 0.02 | MS, KI |
| 4 | 25.42 | 1426 | 1425 | trans-α-Ionone | -- | 2.05 | MS, KI |
| 5 | 25.57 | 1426 | 1426 | cis-α-Ionone | -- | 2.21 | MS, KI |
| 6 | 40.99 | 1162 | 1164 | trans-p-Menthan-7-ol | 2.34 ± 0.05 | 1.81 ± 0.04 | MS, KI |
| Sesquiterpene hydrocarbons | |||||||
| 7 | 20.38 | 1373 | 1373 | α-Ylangene | -- | 0.27 ± 0.01 | MS, KI |
| 8 | 20.69 | 1376 | 1377 | α-Copaene | -- | 0.51 ± 0.02 | MS, KI |
| 9 | 21.23 | 1351 | 1351 | α-Cubebene | -- | 0.41 ± 0.01 | MS, KI |
| 10 | 21.83 | 1409 | 1410 | α-Gurjunene | 0.65 ± 0.03 | 1.09 ± 0.04 | MS, KI |
| 11 | 22.41 | 1442 | 1441 | (+)-1(10)-Aristolene | -- | 2.92 ± 0.06 | MS, KI |
| 12 | 22.50 | 1417 | 1417 | Isocaryophyllene | 1.35 ± 0.06 | -- | MS, KI |
| 13 | 22.53 | 1430 | 1431 | trans-Caryophyllene | -- | 1.58 ± 0.04 | MS, KI |
| 14 | 24.20 | 1706 | 1705 | Farnesane | 1.51 ± 0.05 | 0.73 ± 0.03 | MS, KI |
| 15 | 24.85 | 1493 | 1493 | β-Muurolene | -- | 1.67 ± 0.04 | MS, KI |
| 16 | 25.89 | 1499 | 1500 | α-Muurolene | 1.59 ± 0.03 | 0.78 ± 0.02 | MS, KI |
| 17 | 33.07 | 1460 | 1361 | Dihydroaromadendrene | 5.21 ± 0.06 | 2.71 ± 0.04 | MS, KI |
| Oxygenated Sesquiterpenes | |||||||
| 18 | 20.99 | 1680 | 1682 | 6-epi-shyobunol | -- | 0.99 ± 0.02 | MS, KI |
| 19 | 21.53 | -- | 1675 | Isocaucalol | -- | 0.38 ± 0.01 | MS |
| 20 | 24.25 | 1675 | 1677 | Cis-Z-α-Bisabolene epoxide | -- | 0.45 ± 0.01 | MS, KI |
| 21 | 24.95 | 1597 | 1578 | Widdrol | -- | 0.43 ± 0.02 | MS, KI |
| 22 | 26.31 | 1632 | 1632 | Nerolidyl acetate | -- | 0.48 ± 0.02 | MS, KI |
| 23 | 26.60 | 1494 | 1496 | 4-epi-cubedol | -- | 0.88 ± 0.03 | MS, KI |
| 24 | 27.32 | 1576 | 1577 | Spathulenol | -- | 0.72 ± 0.01 | MS, KI |
| 25 | 29.20 | 1580 | 1580 | Caryophyllene oxide | 6.41 ± 0.05 | 7.86 ± 0.08 | MS, KI |
| 26 | 30.33 | 1605 | 1606 | Humulene oxide | 5.01 ± 0.04 | 4.02 ± 0.04 | MS, KI |
| 27 | 30.66 | 1584 | 1583 | Isoaromadendrene epoxide | -- | 0.48 ± 0.02 | MS, KI |
| 28 | 30.83 | 1657 | 1658 | Cyclolongifolene oxide, dehydro- | 1.34 ± 0.03 | 0.99 ± 0.01 | MS, KI |
| 29 | 31.09 | 1648 | 1650 | Aromadendrene oxide-(1) | -- | 0.71 ± 0.01 | MS, KI |
| 30 | 33.43 | 1683 | 1681 | α-Bisabolol | -- | 1.42 ± 0.03 | MS, KI |
| 31 | 34.41 | 1563 | 1562 | Hexahydrofarnesol | -- | 1.08 ± 0.03 | MS, KI |
| 32 | 38.72 | 1845 | 1845 | Hexahydrofarnesyl acetone | 50.39 ± 0.11 | 35.82 ± 0.09 | MS, KI |
| 33 | 39.00 | -- | 1863 | Nerolidol epoxy acetate | 3.0 ± 0.07 | 1.64 ± 0.02 | MS |
| 34 | 41.27 | 1918 | 1919 | Farnesyl acetone | -- | 7.17 ± 0.05 | MS, KI |
| 35 | 41.72 | 1921 | 1921 | Farnesyl acetone C | 5.21 ± 0.06 | 0.99 ± 0.02 | MS, KI |
| Diterpene hydrocarbons | |||||||
| 36 | 25.06 | 1942 | 1943 | Phytol | 1.83 ± 0.04 | 1.05 ± 0.03 | MS, KI |
| Oxygenated diterpenes | |||||||
| 37 | 26.92 | 1811 | 1812 | Phytan | 1.07 ± 0.04 | 0.72 ± 0.01 | MS, KI |
| Non-oxygenated hydrocarbons | |||||||
| 38 | 16.28 | 1400 | 1400 | n-Tetradecane | 0.74 ± 0.02 | -- | MS, KI |
| 39 | 16.32 | 1500 | 1498 | n-pentadecane | -- | 0.32 ± 0.01 | MS, KI |
| 40 | 34.70 | 1555 | 1556 | 2,6,10-Trimethyltetradecane | 1.02 ± 0.04 | 1.84 ± 0.05 | MS, KI |
| 41 | 40.28 | 1792 | 1792 | Hexadecane, 2,6,11,15-tetramethyl (Crocetane) | 2.31 ± 0.06 | 0.55 ± 0.02 | MS, KI |
| 42 | 40.48 | 1900 | 1902 | n-Nonadecane | -- | 1.17 ± 0.03 | MS, KI |
| 43 | 43.74 | 2000 | 2001 | n-Eicosane | -- | 0.80 ± 0.02 | MS, KI |
| 44 | 46.88 | 2100 | 2100 | n-Heneicosane | 3.31 ± 0.05 | 2.15 ± 0.06 | MS, KI |
| 45 | 47.07 | 2200 | 2200 | n-Docosane | 0.95 ± 0.02 | -- | MS, KI |
| 47 | 52.86 | 2400 | 2401 | n-Tetracosane | 1.38 ± 0.02 | 0.91 ± 0.01 | MS, KI |
| 46 | 49.84 | 3500 | 3500 | n-Pentatriacontane | -- | 0.61 ± 0.02 | MS, KI |
| Oxygenated hydrocarbons | |||||||
| 48 | 31.53 | 1847 | 1851 | 2-Hexadecanol | 1.07 ± 0.03 | -- | MS, KI |
| 50 | 37.11 | 2760 | 2758 | 2-Hexyl-1-decanol | -- | 0.59 ± 0.02 | MS, KI |
| 51 | 37.47 | 2019 | 2019 | 11- Octadecanal | -- | 0.98 ± 0.02 | MS, KI |
| 52 | 40.59 | 2088 | 2088 | Octadecanol | -- | 1.05 ± 0.03 | MS, KI |
| 53 | 48.07 | 2140 | 2143 | 9, 12-Octadecadienoic acid | 4.43 ± 0.06 | 0.47 ± 0.01 | MS, KI |
| Others | |||||||
| 54 | 17.19 | 1300 | 1302 | Dihydroedulan II | 0.66 ± 0.02 | 0.32 ± 0.01 | MS, KI |
| 55 | 32.07 | -- | 1667 | 2-Tetra-butyl-4-isopropyl-5-methylphenol | -- | 0.55 ± 0.01 | MS |
| Monoterpene hydrocarbons | 0.79 | 0.72 | |||||
| Oxygenated monoterpenes | 3.2 | 6.49 | |||||
| Sesquiterpene hydrocarbons | 10.31 | 12.67 | |||||
| Oxygenated sesquiterpenes | 71.36 | 66.51 | |||||
| Diterpene hydrocarbons | 1.83 | 1.05 | |||||
| Oxygenated diterpenes | 1.07 | 0.72 | |||||
| Non-oxygenated hydrocarbons | 9.71 | 8.35 | |||||
| Oxygenated hydrocarbons | 1.07 | 2.62 | |||||
| Others | 0.66 | 0.87 | |||||
| Total | 100 | 100 | |||||
| No | Retention Time | Compound | Concentration (µg g−1) | |
|---|---|---|---|---|
| Coastal Sample | Inland Sample | |||
| Phenolics | ||||
| 1 | 3.114 | Gallic Acid | 69.66 ± 1.14 | 122.11 ± 1.21 |
| 2 | 3.441 | Chlorogenic acid | 1198.24 ± 2.42 | 190.23 ± 1.32 |
| 3 | 3.893 | Caffeine | 26.03 ± 1.12 | 51.45 ± 1.22 |
| 4 | 4.798 | Caffeic Acid | -- | 452.87 ± 2.67 |
| 5 | 5.195 | Syringic acid | 16.97 ± 0.92 | 96.91 ± 1.18 |
| 6 | 5.496 | Rutin | 728.44 ± 2.92 | 199.63 ± 1.56 |
| 7 | 6.655 | Ellagic acid | 22.84 | -- |
| 8 | 7.453 | Coumaric Acid | -- | 95.36 ± 1.02 |
| 9 | 8.05 | Vanillin | 100.63 ± 2.06 | 1284.61 ± 3.08 |
| 10 | 11.118 | Cinnamic Acid | -- | 163.66 ± 1.13 |
| Flavonoids | ||||
| 11 | 3.678 | Catechin | 36.02 ± 0.82 | 270.76 ± 1.82 |
| 12 | 9.329 | Naringenin | -- | 120.56 ± 1.41 |
| 13 | 10.221 | Propyl gallate | 133.09 ± 1.39 | 233.67 ± 2.17 |
| 14 | 10.442 | 4′,7-Dihydroxyisoflavone | 42.19 ± 0.77 | 721.52 ± 3.16 |
| 15 | 10.628 | Quercetin | -- | 603.63 ± 2.97 |
| Treatment | Conc. (mg mL−1) | DPPH | ABTS | ||
|---|---|---|---|---|---|
| Coastal | Inland | Coastal | Inland | ||
| Essential oil extract | 50 | 72.83 ± 1.63*A | 63.44 ± 3.10B | 76.11 ± 1.38A | 69.30 ± 1.68B |
| 40 | 62.57 ± 2.48B | 47.69 ± 2.87C | 68.49 ± 0.99B | 64.92 ± 1.45C | |
| 30 | 49.12 ± 1.79C | 35.89 ± 1.86D | 61.62 ± 1.53D | 49.41 ± 0.46E | |
| 20 | 33.92 ± 2.02D | 26.12 ± 3.42E | 53.14 ± 1.76F | 38.49 ± 1.22G | |
| 10 | 26.78 ± 1.40E | 20.19 ± 0.47F | 37.62 ± 1.53G | 31.30 ± 1.15H | |
| 5 | 18.99 ± 1.55F | 9.00 ± 1.86G | 23.68 ± 0.92I | 19.78 ± 0.46J | |
| IC50 (mg mL−1) | 30.78 | 40.40 | 22.70 | 29.92 | |
| F value | 173.33 | 444.17 | |||
| MeOH extract | 50 | 73.20 ± 1.54A | 65.27 ± 1.03B | 77.96 ± 1.31A | 70.03 ± 1.46B |
| 40 | 61.17 ± 1.69C | 52.46 ± 1.69D | 66.37 ± 1.24C | 62.62 ± 1.02D | |
| 30 | 53.81 ± 1.98D | 43.39 ± 1.32E | 59.11 ± 1.17E | 57.11 ± 1.24E | |
| 20 | 45.20 ± 1.69E | 32.97 ± 1.25F | 48.51 ± 1.60F | 45.26 ± 0.80G | |
| 10 | 34.11 ± 1.25F | 21.31 ± 1.03H | 37.85 ± 0.80H | 33.42 ± 1.09I | |
| 5 | 25.51 ± 1.10G | 9.59 ± 0.59I | 27.50 ± 0.73J | 21.83 ± 0.58K | |
| IC50 (mg mL−1) | 27.00 | 36.57 | 23.12 | 27.41 | |
| F value | 372.68 | 499.06 | |||
| Ascorbic acid | 20 | 66.63 ± 1.02A | 77.90 ± 1.42A | ||
| 15 | 57.89 ± 0.98B | 60.97 ± 1.12B | |||
| 10 | 46.85 ± 0.65C | 49.52 ± 0.86C | |||
| 5 | 39.86 ± 0.45D | 37.01 ± 0.86D | |||
| 2.5 | 8.99 ± 0.05E | 13.09 ± 0.80E | |||
| 1 | 2.00 ± 0.03F | 4.53 ± 0.28F | |||
| IC50 (mg mL−1) | 12.96 | 11.52 | |||
| F value | 318.15 | 299.25 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd-ElGawad, A.M.; Elshamy, A.I.; Al-Rowaily, S.L.; El-Amier, Y.A. Habitat Affects the Chemical Profile, Allelopathy, and Antioxidant Properties of Essential Oils and Phenolic Enriched Extracts of the Invasive Plant Heliotropium Curassavicum. Plants 2019, 8, 482. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8110482
Abd-ElGawad AM, Elshamy AI, Al-Rowaily SL, El-Amier YA. Habitat Affects the Chemical Profile, Allelopathy, and Antioxidant Properties of Essential Oils and Phenolic Enriched Extracts of the Invasive Plant Heliotropium Curassavicum. Plants. 2019; 8(11):482. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8110482
Chicago/Turabian StyleAbd-ElGawad, Ahmed M., Abdelsamed I. Elshamy, Saud L. Al-Rowaily, and Yasser A. El-Amier. 2019. "Habitat Affects the Chemical Profile, Allelopathy, and Antioxidant Properties of Essential Oils and Phenolic Enriched Extracts of the Invasive Plant Heliotropium Curassavicum" Plants 8, no. 11: 482. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8110482