Metabolic Changes Induced by Silver Ions in Carlina acaulis

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
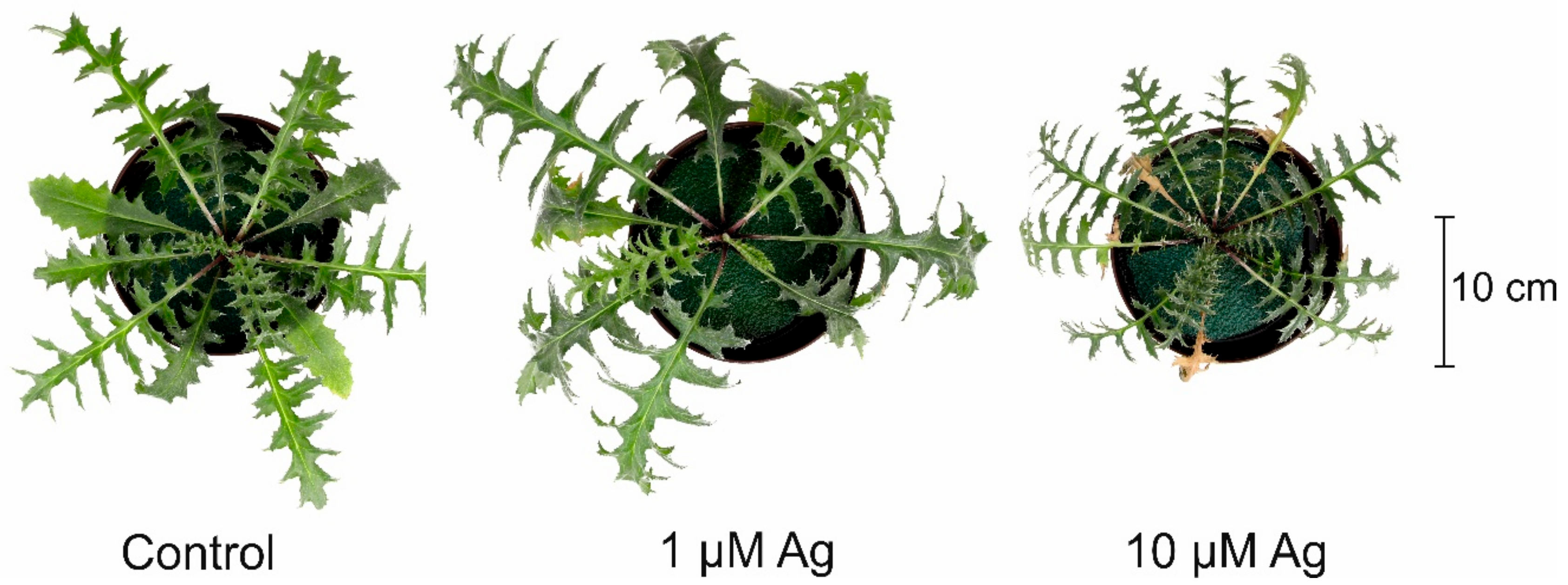
2.1. Impact of Ag(I) on Plant Growth and H2O2 Accumulation
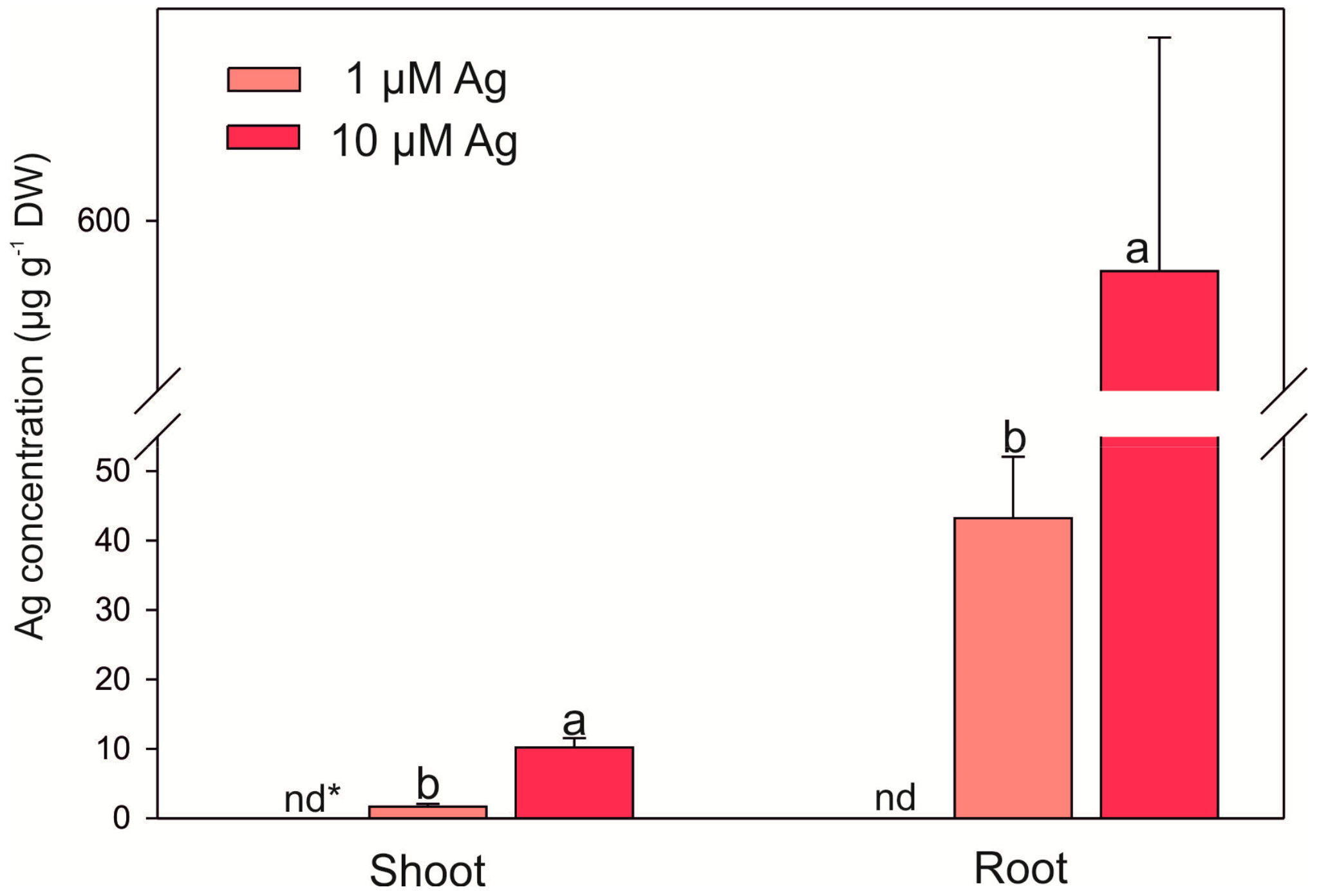
2.2. Bioconcentrations of Ag and Selected Mineral Nutrients
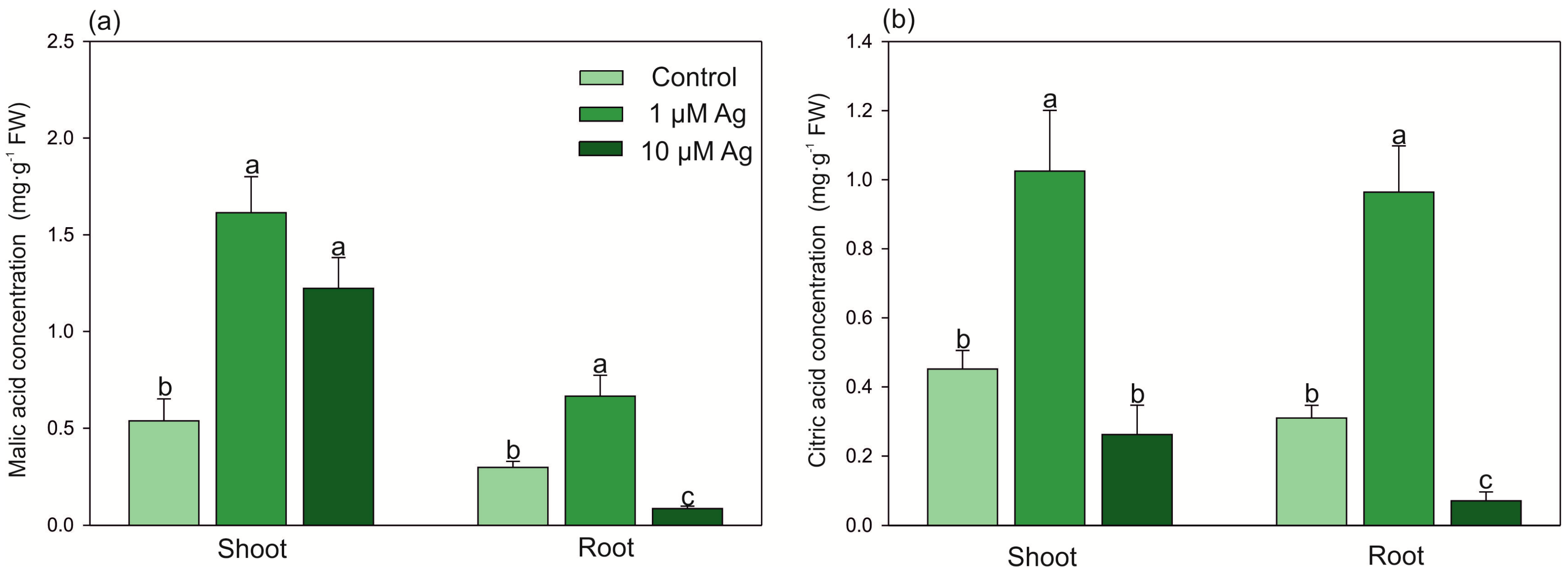
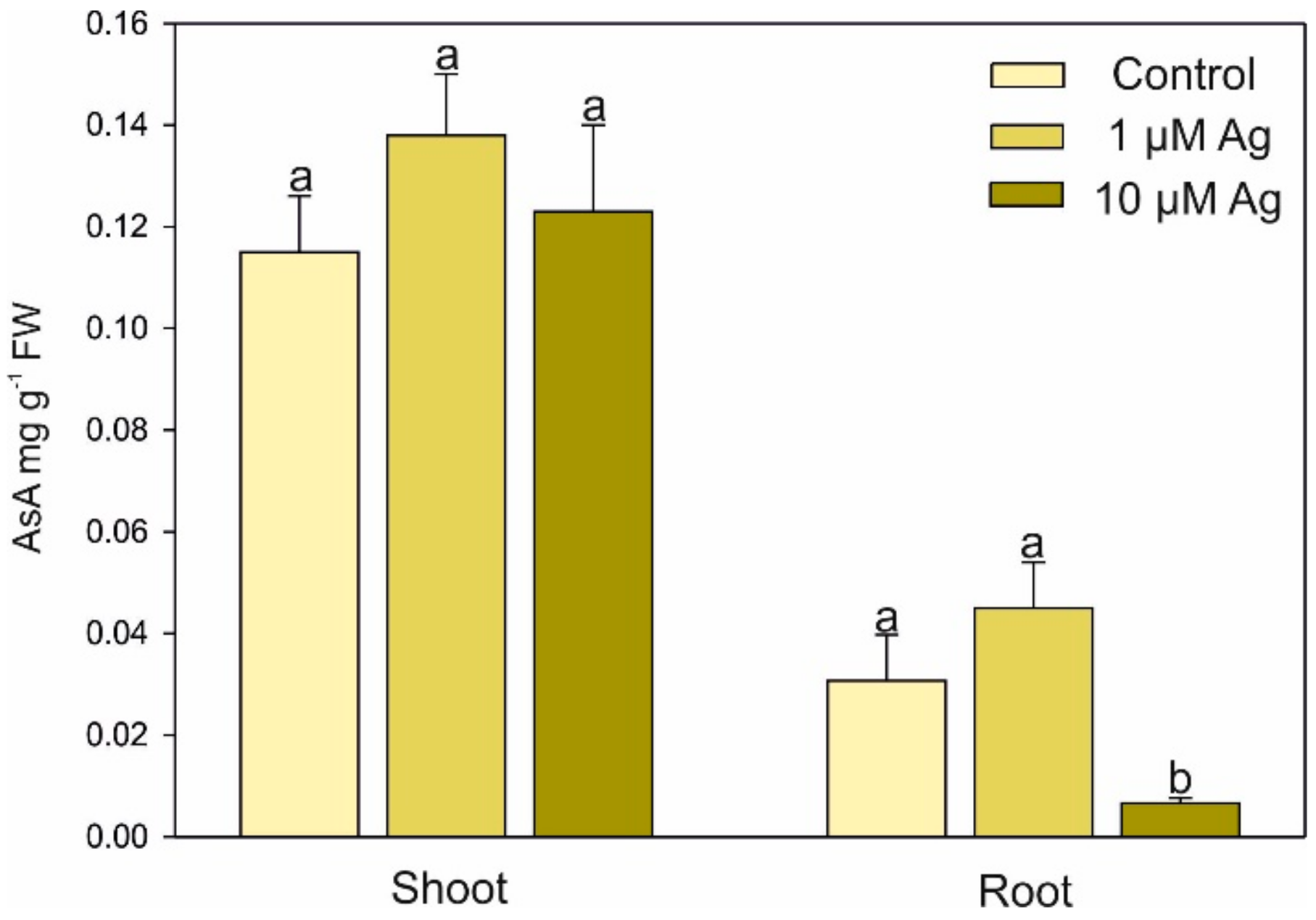
2.3. Changes in Low-Molecular-Weight Organic Acids (LMWOAs), Thiols, and Ascorbic Acid (AsA) Accumulation under Ag(I) Exposure
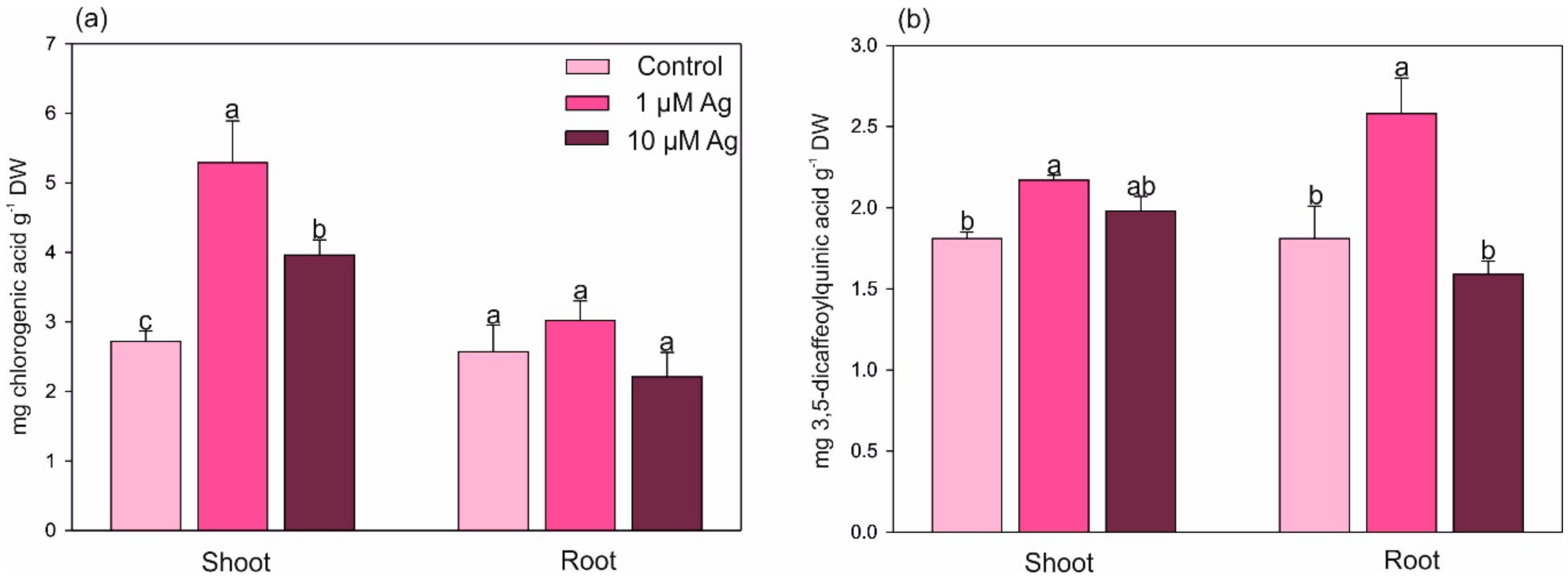
2.4. Changes in the Level of Selected Secondary Metabolites under Ag(I) Exposure
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Determination of Silver and Mineral Nutrients
4.3. Analysis of LMWOAs, Ascorbic Acid, and Thiols
4.4. Quantification of Secondary Metabolites
4.5. Visualization of H2O2
4.6. Statistical Analysis and Experimental Design
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kováčik, J.; Dresler, S.; Wójciak-Kosior, M.; Hladký, J.; Babula, P. Metabolic changes induced by manganese in chamomile. Plant Physiol. Biochem. 2018, 133, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Dresler, S.; Wójciak-Kosior, M.; Sowa, I.; Stanisławski, G.; Bany, I.; Wójcik, M. Effect of short-term Zn/Pb or long-term multi-metal stress on physiological and morphological parameters of metallicolous and nonmetallicolous Echium vulgare L. populations. Plant Physiol. Bioch. 2017, 115, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Dresler, S.; Rutkowska, E.; Bednarek, W.; Stanisławski, G.; Kubrak, T.; Bogucka-Kocka, A.; Wójcik, M. Selected secondary metabolites in Echium vulgare L. populations from nonmetalliferous and metalliferous areas. Phytochemistry 2017, 133, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S. Molecular mechanisms of plant metal tolerance and homeostasis. Planta 2001, 212, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Kováčik, J.; Klejdus, B. Dynamics of phenolic acids and lignin accumulation in metal-treated Matricaria chamomilla roots. Plant Cell Reports 2008, 27, 605–615. [Google Scholar] [CrossRef]
- Sytar, O.; Kumar, A.; Latowski, D.; Kuczynska, P.; Strzałka, K.; Prasad, M.N.V. Heavy metal-induced oxidative damage, defense reactions, and detoxification mechanisms in plants. Acta Physiol. Plant. 2013, 35, 985–999. [Google Scholar] [CrossRef]
- Niggeweg, R.; Michael, A.J.; Martin, C. Engineering plants with increased levels of the antioxidant chlorogenic acid. Nat. Biotechnol. 2004, 22, 746–754. [Google Scholar] [CrossRef]
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Pol. J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Ratte, H.T. Bioaccumulation and toxicity of silver compounds: A review. Environ. Toxicol. Chem. 1999, 18, 89–108. [Google Scholar] [CrossRef]
- Blamey, F.P.C.; Kopittke, P.M.; Wehr, J.B.; Kinraide, T.B.; Menzies, N.W. Rhizotoxic effects of silver in cowpea seedlings. Environ. Toxicol. Chem. 2010, 29, 2072–2078. [Google Scholar] [CrossRef] [PubMed]
- Purcell, T.W.; Peters, J.J. Sources of silver in the environment. Environ. Toxicol. Chem. 1998, 17, 539–546. [Google Scholar] [CrossRef]
- Das, D.; Lakshmi, V.; Das, N.; Vimala, R. Studies on toxicity of ag (I) on plants and microbes. Res. J. Pharm. Biol. Chem. Sci. 2013, 4, 176–188. [Google Scholar]
- Wallace, A.; Mueller, R.T. Recovery from acute silver toxicity by bush beans grown in solution culture. J. Plant Nutr. 1980, 2, 93–95. [Google Scholar] [CrossRef]
- Strzemski, M.; Wójciak-Kosior, M.; Sowa, I.; Załuski, D.; Verpoorte, R. Historical and traditional medical applications of Carlina acaulis L.—A critical ethnopharmacological review. J. of Ethnopharmacol. 2019, 239, 111842. [Google Scholar] [CrossRef]
- Strzemski, M.; Wójciak-Kosior, M.; Sowa, I.; Załuski, D.; Szwerc, W.; Sawicki, J.; Kocjan, R.; Feldo, M.; Dresler, S. Carlina vulgaris L as a source of phytochemicals with antioxidant activity. Oxid. Med. Cell. Longev. 2017, 2017, 1891849. Available online: https://www.hindawi.com/journals/omcl/2017/1891849/ (accessed on 15 April 2019). [CrossRef]
- Jiang, H.-S.; Li, M.; Chang, F.-Y.; Li, W.; Yin, L.-Y. Physiological analysis of silver nanoparticles and AgNO3 toxicity to Spirodela polyrhiza. Environmen. Toxicol. Chem. 2012, 31, 1880–1886. [Google Scholar] [CrossRef]
- Yang, Q.; Shan, W.; Hu, L.; Zhao, Y.; Hou, Y.; Yin, Y.; Liang, Y.; Wang, F.; Cai, Y.; Liu, J.; et al. Uptake and transformation of silver nanoparticles and ions by rice plants revealed by dual stable isotope tracing. Environ. Sci. Technol. 2019, 53, 625–633. [Google Scholar] [CrossRef]
- Wang, J.; Koo, Y.; Alexander, A.; Yang, Y.; Westerhof, S.; Zhang, Q.; Schnoor, J.L.; Colvin, V.L.; Braam, J.; Alvarez, P.J.J. Phytostimulation of poplars and Arabidopsis exposed to silver nanoparticles and Ag+ at sublethal concentrations. Environ. Sci. Technol. 2013, 47, 5442–5449. [Google Scholar] [CrossRef]
- Qian, H.; Peng, X.; Han, X.; Ren, J.; Sun, L.; Fu, Z. Comparison of the toxicity of silver nanoparticles and silver ions on the growth of terrestrial plant model Arabidopsis thaliana. J. Environ. Sci. 2013, 25, 1947–1956. [Google Scholar] [CrossRef]
- Cvjetko, P.; Milošić, A.; Domijan, A.-M.; Vinković Vrček, I.; Tolić, S.; Peharec Štefanić, P.; Letofsky-Papst, I.; Tkalec, M.; Balen, B. Toxicity of silver ions and differently coated silver nanoparticles in Allium cepa roots. Ecotoxicol. Environ. Saf. 2017, 137, 18–28. [Google Scholar] [CrossRef]
- Calabrese, E.J. Evidence that hormesis represents an “overcompensation” response to a disruption in homeostasis. Ecotoxicol. Environ. Saf. 1999, 42, 135–137. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.-H.; Yao, J.-L.; Cohen, D.; Morris, B. Ethylene inhibitors enhance in vitro root formation from apple shoot cultures. Plant Cell Rep. 1998, 17, 211–214. [Google Scholar] [CrossRef]
- Hirsch, M.P. Availability of sludge-borne silver to agricultural crops. Environ. Toxicol. Chem. 1998, 17, 610–616. [Google Scholar] [CrossRef]
- Kováčik, J.; Dresler, S. Calcium availability but not its content modulates metal toxicity in Scenedesmus quadricauda. Ecotoxicol. Environ. Saf. 2018, 147, 664–669. [Google Scholar] [CrossRef] [PubMed]
- Nedjimi, B. Heavy metal tolerance in two Algerian saltbushes: A review on plant responses to cadmium and role of calcium in its mitigation. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer Singapore: Singapore, 2018; pp. 205–220. ISBN 978-981-10-9044-8. [Google Scholar]
- Klíma, P.; Laňková, M.; Vandenbussche, F.; Van Der Straeten, D.; Petrášek, J. Silver ions increase plasma membrane permeability through modulation of intracellular calcium levels in tobacco BY-2 cells. Plant Cell Rep. 2018, 37, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Kutrowska, A.; Szelag, M. Low-molecular weight organic acids and peptides involved in the long-distance transport of trace metals. Acta Physiol. Plant. 2014, 36, 1957–1968. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B.; Dresler, S.; Matraszek, R. Exogenous malic and acetic acids reduce cadmium phytotoxicity and enhance cadmium accumulation in roots of sunflower plants. Plant Physiol. Bioch. 2015, 94, 225–234. [Google Scholar] [CrossRef]
- Zenk, M.H. Heavy metal detoxification in higher plants—A review. Gene 1996, 179, 21–30. [Google Scholar] [CrossRef]
- Mehra, R.K.; Tran, K.; Scott, G.W.; Mulchandani, P.; Saini, S.S. Ag(I)-binding to phytochelatins. J. Inorg. Biochem. 1996, 61, 125–142. [Google Scholar] [CrossRef]
- Grill, E.; Winnacker, E.-L.; Zenk, M.H. Phytochelatins, a class of heavy-metal-binding peptides from plants, are functionally analogous to metallothioneins. Proc. Natl. Acad. Sci. USA 1987, 84, 439–443. [Google Scholar] [CrossRef]
- Figueroa, J.A.L.; Wrobel, K.; Afton, S.; Caruso, J.A.; Felix Gutierrez Corona, J.; Wrobel, K. Effect of some heavy metals and soil humic substances on the phytochelatin production in wild plants from silver mine areas of Guanajuato, Mexico. Chemosphere 2008, 70, 2084–2091. [Google Scholar] [CrossRef] [PubMed]
- Le Faucheur, S.; Schildknecht, F.; Behra, R.; Sigg, L. Thiols in Scenedesmus vacuolatus upon exposure to metals and metalloids. Aquat. Toxicol. 2006, 80, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Barbasz, A.; Kreczmer, B.; Oćwieja, M. Effects of exposure of callus cells of two wheat varieties to silver nanoparticles and silver salt (AgNO3). Acta Physiol. Plant. 2016, 38, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Jozefczak, M.; Remans, T.; Vangronsveld, J.; Cuypers, A. Glutathione is a key player in metal-induced oxidative stress defenses. Int. J. Mol. Sci. 2012, 13, 3145–3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kováčik, J.; Klejdus, B.; Babula, P.; Hedbavny, J. Ascorbic acid affects short-term response of Scenedesmus quadricauda to cadmium excess. Algal Res. 2017, 24, 354–359. [Google Scholar] [CrossRef]
- Dresler, S.; Maksymiec, W. Capillary zone electrophoresis for determination of reduced and oxidised ascorbate and glutathione in roots and leaf segments of Zea mays plants exposed to Cd and Cu. Acta Sci. Pol. Hortorum Cultus 2013, 12, 143–155. [Google Scholar]
- Bagherzadeh Homaee, M.; Ehsanpour, A.A. Silver nanoparticles and silver ions: Oxidative stress responses and toxicity in potato (Solanum tuberosum L.) grown in vitro. Hortic. Environ. Biotechnol. 2016, 57, 544–553. [Google Scholar] [CrossRef]
- Sakihama, Y.; Cohen, M.F.; Grace, S.C.; Yamasaki, H. Plant phenolic antioxidant and prooxidant activities: Phenolics-induced oxidative damage mediated by metals in plants. Toxicology 2002, 177, 67–80. [Google Scholar] [CrossRef]
- Khan, I.; Raza, M.A.; Bin Khalid, M.H.; Awan, S.A.; Raja, N.I.; Zhang, X.; Min, S.; Wu, B.C.; Hassan, M.J.; Huang, L. Physiological and biochemical responses of pearl millet (Pennisetum glaucum L.) seedlings exposed to silver nitrate (AgNO3) and silver nanoparticles (AgNPs). Int. J. Environ. Res. Public Health 2019, 16, 2261. [Google Scholar] [CrossRef] [Green Version]
- Krishnaraj, C.; Jagan, E.G.; Ramachandran, R.; Abirami, S.M.; Mohan, N.; Kalaichelvan, P.T. Effect of biologically synthesized silver nanoparticles on Bacopa monnieri (Linn.) Wettst. plant growth metabolism. Process Biochem. 2012, 47, 651–658. [Google Scholar] [CrossRef]
- Fayez, K.A.; El-Deeb, B.A.; Mostafa, N.Y. Toxicity of biosynthetic silver nanoparticles on the growth, cell ultrastructure and physiological activities of barley plant. Acta Physiol. Plant. 2017, 39, 155. [Google Scholar] [CrossRef]
- Do Nascimento, P.G.G.; Lemos, T.L.G.; Bizerra, A.M.C.; Arriaga, Â.M.C.; Ferreira, D.A.; Santiago, G.M.P.; Braz-Filho, R.; Costa, J.G.M. Antibacterial and antioxidant activities of ursolic acid and derivatives. Molecules 2014, 19, 1317–1327. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Guo, Q.; Wang, Q.; Zhou, L.; Zhang, Z.; Zhang, L.; Huang, T. Effects of lead, copper and cadmium stresses on growth and inherent quality of Prunalla vulgaris. Zhongguo Zhongyao Zazhi 2010, 35, 263–267. (In Chinese) [Google Scholar] [PubMed]
- Wang, Q.J.; Lei, X.Y.; Zheng, L.P.; Wang, J.W. Molecular characterization of an elicitor-responsive 3-hydroxy-3-methylglutaryl coenzyme A reductase gene involved in oleanolic acid production in cell cultures of Achyranthes bidentata. Plant Growth Regul. 2017, 81, 335–343. [Google Scholar] [CrossRef]
- Dresler, S.; Hanaka, A.; Bednarek, W.; Maksymiec, W. Accumulation of low-molecular-weight organic acids in roots and leaf segments of Zea mays plants treated with cadmium and copper. Acta Physiol. Plant. 2014, 36, 1565–1575. [Google Scholar] [CrossRef] [Green Version]
- Dresler, S.; Hawrylak-Nowak, B.; Kováčik, J.; Pochwatka, M.; Hanaka, A.; Strzemski, M.; Sowa, I.; Wójciak-Kosior, M. Allantoin attenuates cadmium-induced toxicity in cucumber plants. Ecotoxicol. Environ. Saf. 2019, 170, 120–126. [Google Scholar] [CrossRef]
- Strzemski, M.; Wójciak-Kosior, M.; Sowa, I.; Rutkowska, E.; Szwerc, W.; Kocjan, R.; Latalski, M. Carlina species as a new source of bioactive pentacyclic triterpenes. Ind. Crop. Prod. 2016, 94, 498–504. [Google Scholar] [CrossRef]
- Sowa, I.; Paduch, R.; Strzemski, M.; Zielińska, S.; Rydzik-Strzemska, E.; Sawicki, J.; Kocjan, R.; Polkowski, J.; Matkowski, A.; Latalski, M.; et al. Proliferative and antioxidant activity of Symphytum officinale root extract. Nat. Prod. Res. 2018, 32, 605–609. [Google Scholar] [CrossRef]
- Strzemski, M.; Wójciak-Kosior, M.; Sowa, I.; Kocjan, R.; Tyszczuk-Rotko, K. Methodological approach to determine carlina oxide—A main volatile constituent of Carlina acaulis L. essential oil. Talanta 2019, 191, 504–508. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B.; Dresler, S.; Rubinowska, K.; Matraszek-Gawron, R.; Woch, W.; Hasanuzzaman, M. Selenium biofortification enhances the growth and alters the physiological response of lamb’s lettuce grown under high temperature stress. Plant Physiol. Bioch. 2018, 127, 446–456. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | K (mg g−1 DW) | P (mg g−1 DW) | ||
|---|---|---|---|---|
| Shoot | Root | Shoot | Root | |
| Control | 55.45 ± 3.83 a | 41.93 ± 2.35 a | 3.48 ± 0.35 a | 5.69 ± 0.37 a |
| 1 µM Ag | 55.01 ± 4.52 a | 40.17 ± 5.22 a | 3.46 ± 0.65 a | 5.79 ± 0.48 a |
| 10 µM Ag | 52.02 ± 1.53 a | 25.19 ± 2.19 b | 2.75 ± 0.23 a | 3.84 ± 0.19 b |
| Ca (mg g−1 DW) | Mg (mg g−1 DW) | |||
| Shoot | Root | Shoot | Root | |
| Control | 12.22 ± 0.73 b | 5.09 ± 0.42 b | 4.26 ± 0.31 a | 2.32 ± 0.09 a |
| 1 µM Ag | 14.37 ± 0.47 a | 5.49 ± 0.47 b | 4.20 ± 0.56 a | 2.74 ± 0.03 a |
| 10 µM Ag | 9.26 ± 0.21 c | 7.67 ± 036 a | 3.75 ± 0.21 a | 2.52 ± 0.04 a |
| Zn (µg g−1 DW) | Cu (µg g−1 DW) | |||
| Shoot | Root | Shoot | Root | |
| Control | 21.57 ± 4.02 a | 58.76 ± 7.82 a | 5.32 ± 0.17 a | 14.06 ± 1.40 a |
| 1 µM Ag | 24.89 ± 2.90 a | 61.48 ± 12.02 a | 3.73 ± 0.41 b | 11.54 ± 1.14 a |
| 10 µM Ag | 20.64 ± 2.86 a | 56.08 ± 9.68 a | 4.54 ± 0.11 ab | 14.92 ± 2.17 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dresler, S.; Hawrylak-Nowak, B.; Strzemski, M.; Wójciak-Kosior, M.; Sowa, I.; Hanaka, A.; Gołoś, I.; Skalska-Kamińska, A.; Cieślak, M.; Kováčik, J. Metabolic Changes Induced by Silver Ions in Carlina acaulis. Plants 2019, 8, 517. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8110517
Dresler S, Hawrylak-Nowak B, Strzemski M, Wójciak-Kosior M, Sowa I, Hanaka A, Gołoś I, Skalska-Kamińska A, Cieślak M, Kováčik J. Metabolic Changes Induced by Silver Ions in Carlina acaulis. Plants. 2019; 8(11):517. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8110517
Chicago/Turabian StyleDresler, Sławomir, Barbara Hawrylak-Nowak, Maciej Strzemski, Magdalena Wójciak-Kosior, Ireneusz Sowa, Agnieszka Hanaka, Iwona Gołoś, Agnieszka Skalska-Kamińska, Małgorzata Cieślak, and Jozef Kováčik. 2019. "Metabolic Changes Induced by Silver Ions in Carlina acaulis" Plants 8, no. 11: 517. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8110517