
Seasonal and Interannual Fluctuation of the Microbial Soil Community in a Maize Field under Long-Term Conservation Agriculture Management

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and methods
2.1. Sampling Site
2.2. Physical and Chemical Analyses of Soil
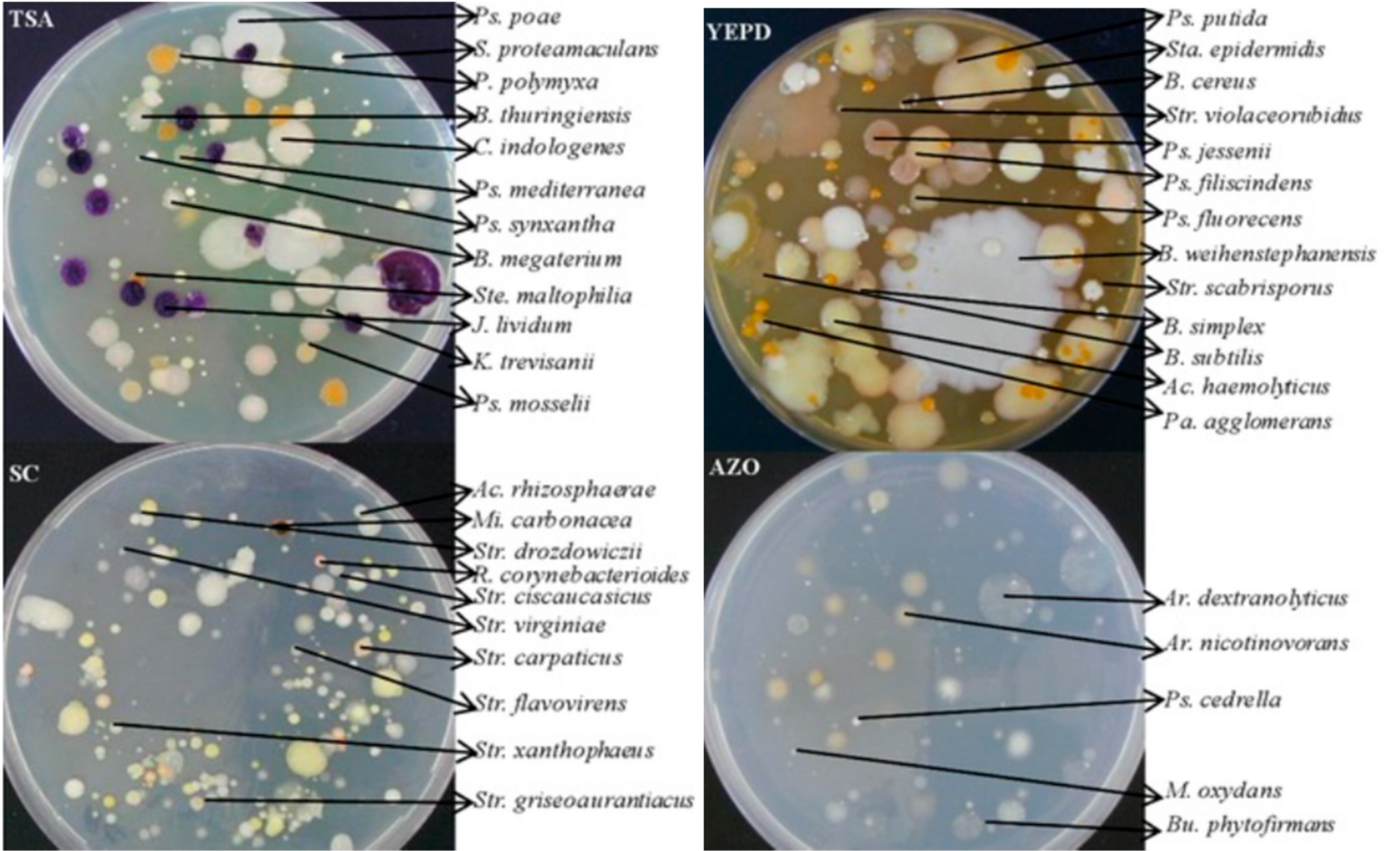
2.3. Plating of Soil Samples and De Visu Analysis of Cultured Microorganisms
2.4. DNA Extraction from Soil and Cultured Bacteria
2.5. PCR Amplification, Sequencing of 16S Ribosomal DNA and Bacteria Identification
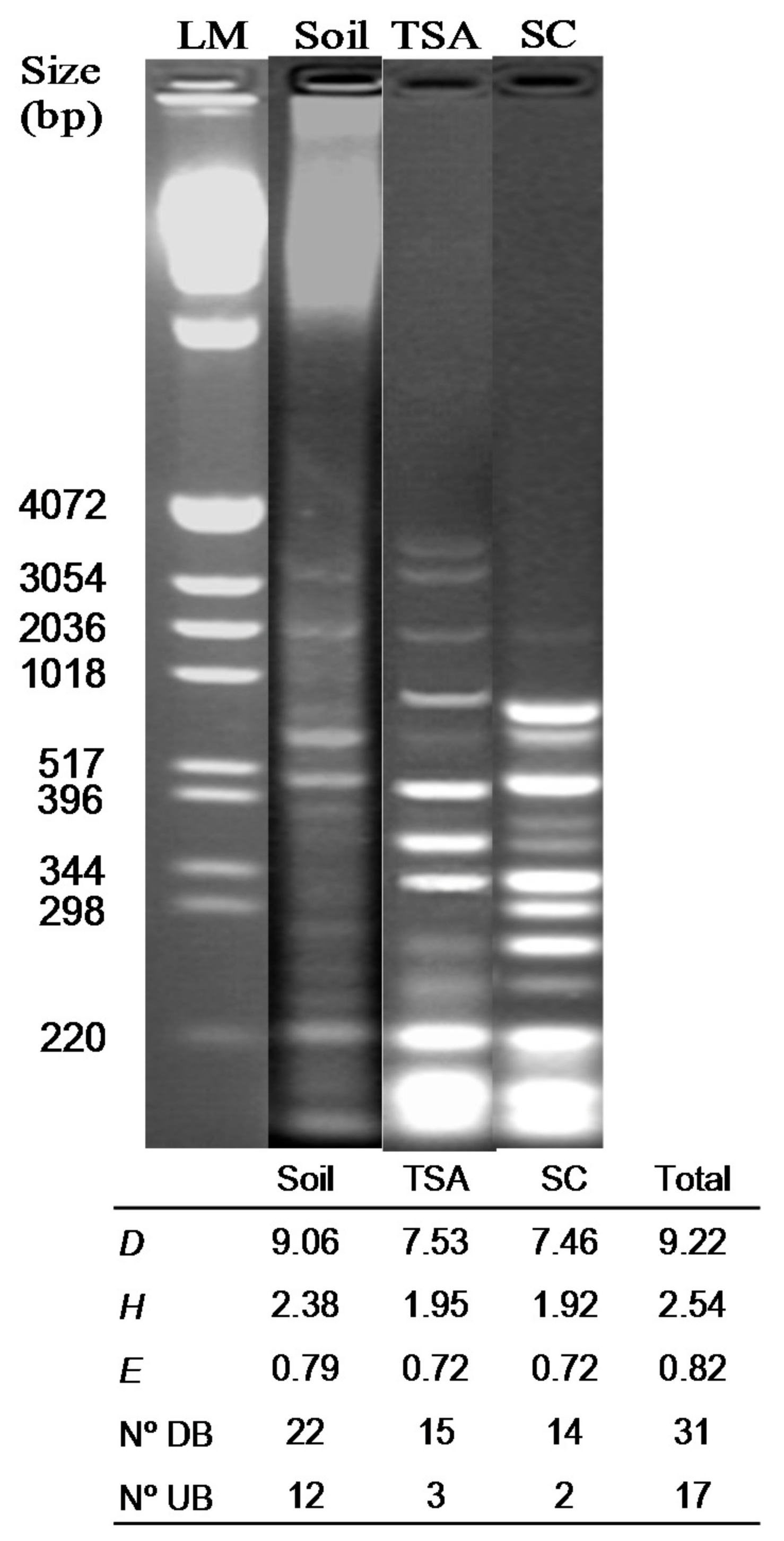
2.6. PCR-Agarose Gel Electrophoresis of the Bacterial Soil Community
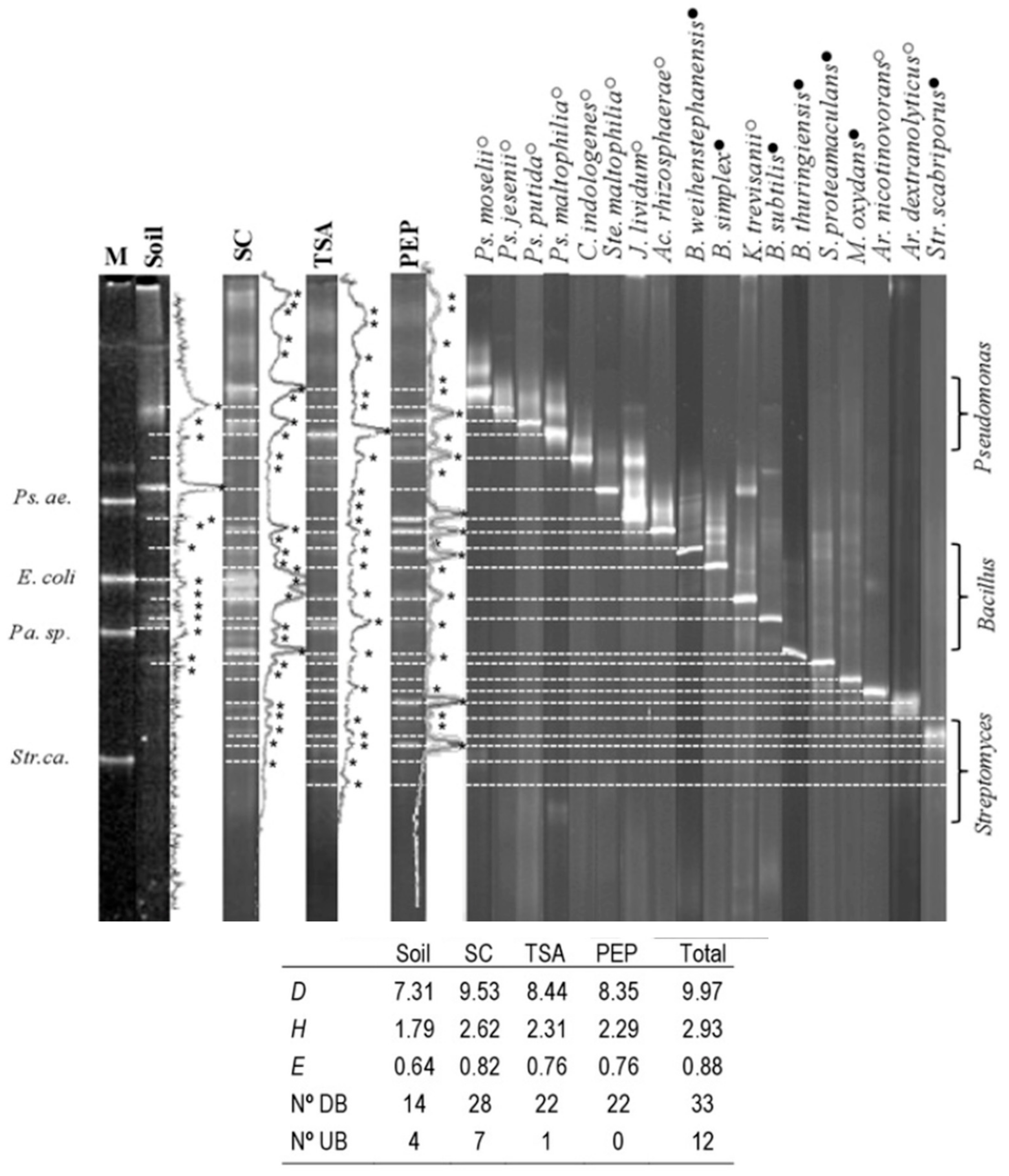
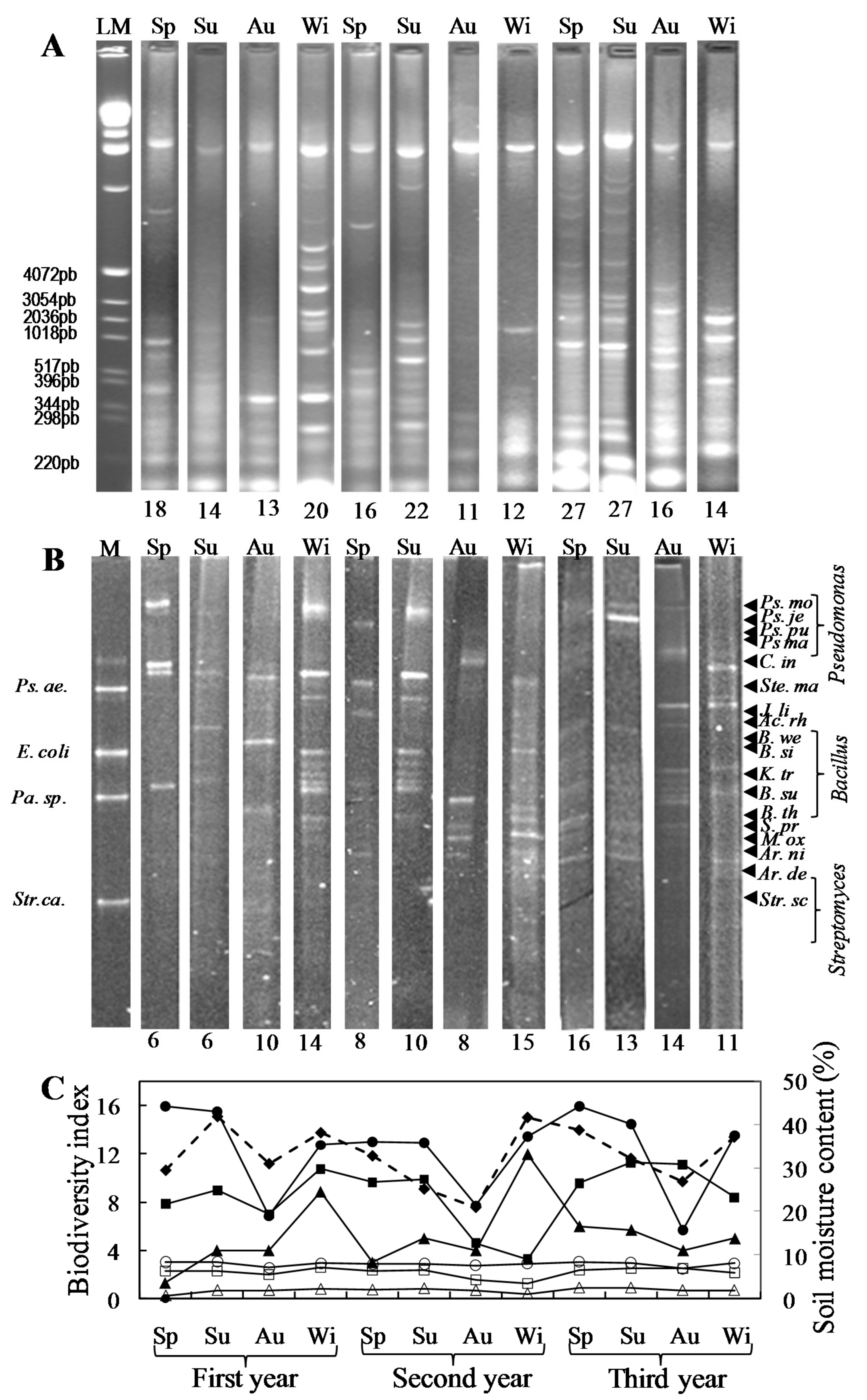
2.7. PCR-DGGE Analysis of the Bacterial Soil Community
2.8. Statistical Data Analysis
3. Results
3.1. Analysis of Cultured Microorganisms
3.2. PCR-Agarose and PCR-DGGE Analysis of Soil and Cultured Bacteria DNA
3.3. Seasonal Fluctuation and Interannual Stability of the Soil Microbial Community in a Long-Term No-Tillage Maize Crop
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Spedding, T.A.; Hamel, C.; Mehuys, G.R.; Madramootoo, C.A. Soil microbial dynamics in maize-growing soil under different tillage and residue management systems. Soil Biol. Biochem. 2004, 36, 499–512. [Google Scholar] [CrossRef]
- De Ploey, J.A.; Imeson, A.; Oldeman, L.R. Soil erosion, soil degradation and climatic change. In Land Use Changes in Europe; Brower, F.M., Thomas, A.J., Chadwick, M.J., Eds.; Kluwer Academic Publisher: London, UK, 1991; pp. 275–292. [Google Scholar]
- Bezdicek, D.F.; Beaver, T.; Granatstein, D. Subsoil ridge tillage and lime effects on soil microbial activity, soil pH, erosion, and wheat and pea yield in the Pacific Northwest, USA. Soil Tillage Res. 2003, 74, 55–63. [Google Scholar] [CrossRef]
- McDaniel, M.D.; Tiemann, L.K.; Grandy, A.S. Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecol. Appl. 2014, 24, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.C.; Smith, K.L. Soil microbial diversity and the sustainability of agricultural soils. In The Significance and Regulation of Soil Biodiversity; Springer: Dordrecht, The Netherlands, 1995; pp. 75–86. [Google Scholar]
- López-Garrido, R.; Madejón, E.; Moreno, F.; Murillo, J.M. Conservation tillage influence on carbon dynamics under Mediterranean conditions. Pedosphere 2014, 24, 65–75. [Google Scholar] [CrossRef]
- Helgason, B.L.; Walley, F.L.; Germida, J.J. Long-term no-till management affects microbial biomass but not community composition in Canadian prairie agroecosytems. Soil Biol. Biochem. 2010, 42, 2192–2202. [Google Scholar] [CrossRef]
- Guo, L.J.; Zhang, Z.S.; Wang, D.D.; Li, C.F.; Cao, C.G. Effects of short-term conservation management practices on soil organic carbon fractions and microbial community composition under a rice-wheat rotation system. Biol. Fertil. Soils 2015, 51, 65–75. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Rice, W.A.; Clayton, G.W. Soil microbial diversity and community structure under wheat as influenced by tillage and crop rotation. Soil Biol. Biochem. 1998, 30, 1733–1741. [Google Scholar] [CrossRef]
- Emmerling, C.; Udelhoven, T.; Schroder, D. Response of soil microbial biomass and activity to agricultural de-intensification over a 10 year period. Soil Biol. Biochem. 2001, 33, 2105–2114. [Google Scholar] [CrossRef]
- Muñoz, A.; López-Piñeiro, A.; Ramírez, M. Soil quality attributes of conservation management regimes in a semi-arid region of south western Spain. Soil Tillage Res. 2007, 95, 255–265. [Google Scholar] [CrossRef]
- Powlson, D.S.; Brookes, P.C.; Christensen, B.T. Measurement of soil microbial biomass provides an early indication of changes in total soil organic matter due to straw incorporation. Soil Biol. Biochem. 1987, 19, 159–164. [Google Scholar] [CrossRef]
- Sparling, G.P. Soil microbial biomass activity and nutrient cycling an indicator of soil health. In Biological Indicators of Soil Health; Pankhurst, C., Doube, B.M., Gupta, V.V.S.R., Eds.; CAB International: Wallingford, UK, 1997; pp. 97–119. [Google Scholar]
- Franzluebbers, A.J.; Langdale, G.W.; Schomberg, H.H. Soil carbon, nitrogen, and aggregation in response to type and frequency of tillage. Soil Sci. Soc. Am. J. 1999, 63, 349–355. [Google Scholar] [CrossRef]
- Avidano, L.; Gamalero, E.; Cossa, G.P.; Carraro, E. Characterization of soil health in an Italian polluted site by using microorganisms as bioindicators. Appl. Soil Ecol. 2005, 30, 21–33. [Google Scholar] [CrossRef]
- Tate, R.L. Soil Microbiology; John Wiley and Sons: New York, NY, USA, 2000; p. 508. [Google Scholar]
- Nielsen, M.N.; Winding, A. Microorganisms as Indicators of Soil Health; Ministry of Environment Technical Report No. 388; National Environmental Research Institute: Aarhus, Denmark, 2002. [Google Scholar]
- Mangalassery, S.; Mooney, S.J.; Sparkes, D.L.; Fraser, W.T.; Sjögersten, S. Impacts of zero tillage on soil enzyme activities, microbial characteristics and organic matter functional chemistry in temperate soils. Eur. J. Soil Biol. 2015, 68, 9–17. [Google Scholar] [CrossRef]
- Bell, C.; Holder-Franklin, M.; Franklin, M. Seasonal fluctuations in river bacteria as measured by multivariate statistical analysis of continuous cultures. Can. J. Microbiol. 1982, 28, 959–975. [Google Scholar] [CrossRef] [PubMed]
- Bloem, J.; Lebbink, G.; Zwart, K.B.; Bouwman, L.A.; Burgers, S.L.G.E.; de Vos, J.A.; de Ruiter, P.C. Dynamics of microorganisms, microbivores and nitrogen mineralisation in winter wheat fields under conventional and integrated management. Agric. Ecosyst. Environ. 1994, 51, 129–143. [Google Scholar] [CrossRef]
- Zogg, G.P.; Zak, D.R.; Ringelberg, D.B.; White, D.C.; MacDonald, N.W.; Pregitzer, K.S. Compositional and functional shifts in microbial communities due to soil warming. Soil Sci. Soc. Am. J. 1997, 61, 475–481. [Google Scholar] [CrossRef]
- Bossio, D.; Scow, K.; Gunapala, N.; Graham, K. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Frenk, S.; Dag, A.; Yermiyahu, U.; Zipori, I.; Hadar, Y.; Minz, D. Seasonal effect and anthropogenic impact on the composition of the active bacterial community in Mediterranean orchard soil. FEMS Microbiol. Ecol. 2015, 91, fiv096. [Google Scholar] [CrossRef] [PubMed]
- Janssen, P.H.; Yates, P.S.; Grinton, B.E.; Taylor, P.M.; Sait, M. Improved culturability of soil bacteria and isolation in pure culture of novel members of the divisions Acidobacteria, Actinobacteria, Proteobacteria, and Verrucomicrobia. Appl. Environ. Microb. 2002, 68, 2391–2396. [Google Scholar] [CrossRef]
- Hugenholtz, P.; Goebel, B.M.; Pace, N.R. Impact of culture-Independent studies on the emerging phylogenetic view of bacterial diversity. J. Bacteriol. 1998, 180, 4765–4774. [Google Scholar] [PubMed]
- Ellis, R.J.; Morgan, P.; Weightman, A.J.; Fry, J.C. Cultivation-dependent and -independent approaches for determining bacterial diversity in heavy-metal-contaminated soil. Appl. Environ. Microb. 2003, 69, 3223–3230. [Google Scholar] [CrossRef]
- Edenborn, S.L.; Sexstone, A.J. DGGE fingerprinting of culturable soil bacterial communities complements culture-independent analyses. Soil Biol. Biochem. 2007, 39, 1570–1579. [Google Scholar] [CrossRef]
- Al-Awadhi, H.; Dashti, N.; Khanafer, M.; Al-Mailem, D.; Ali, N.; Radwan, S. Bias problems in culture-independent analysis of environmental bacterial communities: A representative study on hydrocarbonoclastic bacteria. SpringerPlus 2013, 2, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bent, S.J.; Pierson, J.D.; Forney, L.J.; Danovaro, R.; Luna, G.M.; Dell’Anno, A.; Pietrangeli, B. Measuring species richness based on microbial community fingerprints: The emperor has no clothes. Appl. Environ. Microb. 2007, 73, 2399–2401. [Google Scholar] [CrossRef] [PubMed]
- Danovaro, R.; Luna, G.M.; Dell’Anno, A.; Pietrangeli, B. Comparison of two fingerprinting techniques, terminal restriction fragment length polymorphism and automated ribosomal intergenic spacer analysis, for determination of bacterial diversity in aquatic environments. Appl. Environ. Microb. 2006, 72, 5982–5989. [Google Scholar] [CrossRef] [PubMed]
- Douterelo, I.; Boxall, J.B.; Deines, P.; Sekar, R.; Fish, K.E.; Biggs, C.A. Methodological approaches for studying the microbial ecology of drinking water distribution systems. Water Res. 2014, 65, 134–156. [Google Scholar] [CrossRef] [PubMed]
- Barns, S.M.; Takala, S.L.; Kuske, C.R. Wide distribution and diversity of members of the bacterial kingdom Acidobacterium in the environment. Appl. Environ. Microb. 1999, 65, 1731–1737. [Google Scholar]
- Smit, E.; Leeflang, P.; Gommans, S.; Van den Broek, J.; Van Mil, S.; Wernars, K. Diversity and seasonal fluctuations of the dominant members of the bacterial soil community in a wheat field as determined by cultivation and molecular methods. Appl. Environ. Microb. 2001, 67, 2284–2291. [Google Scholar] [CrossRef] [PubMed]
- Lipson, D.A.; Schmidt, S.K. Seasonal changes in an alpine soil bacterial community in the Colorado Rocky Mountains. Appl. Environ. Microb. 2004, 70, 2867–2879. [Google Scholar] [CrossRef]
- Ibekwe, A.M.; Papiernik, S.K.; Gan, J.; Yates, S.R.; Yang, C.H.; Crowley, D.E. Impact of fumigants on soil microbial communities. Appl. Environ. Microb. 2001, 67, 3245–3257. [Google Scholar] [CrossRef] [PubMed]
- Kisand, V.; Wikner, J. Combining culture-dependent and -independent methodologies for estimation of richness of estuarine bacterioplankton consuming riverine dissolved organic matter. Appl. Environ. Microb. 2003, 69, 3607–3616. [Google Scholar] [CrossRef]
- Liu, B.; Tu, C.; Hu, S.; Gumpertz, M.; Ristaino, J.B. Effect of organic, sustainable, and conventional management strategies in grower fields on soil physical, chemical, and biological factors and the incidence of Southern blight. Appl. Soil Ecol. 2007, 37, 202–214. [Google Scholar] [CrossRef]
- ISSS-ISRIC-FAO (Food and Agriculture Organization of the United Nations). World Reference Base for Soil Resources; Draft; Spaargaren, O., Ed.; FAO: Wageningen, The Netherlands; Rome, Italy, 1994. [Google Scholar]
- United Nations Environment Programme (UNEP). World Map of Desertification, 2nd ed.; Arnold: London, UK, 1997. [Google Scholar]
- Nelson, D.; Sommers, L. Total C, organic C and organic matter. In Methods of Soil Analysis. Soil Science Society of America Book Series No 5, Part 3—Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Summer, M.E., Eds.; America Society of Agronomy–Soil Science Society of America Publisher: Madison, WI, USA, 1996. [Google Scholar]
- Bremner, J.; Mulvaney, C. Nitrogen-Total. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties, Agronomy Monograph; ASA Publisher: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar]
- Hurst, C.J.; Crawford, R.L.; Knudsen, G.R.; McInerney, M.J. Manual of Environmental Microbiology, 2nd ed.; ASM Press: Washington, DC, USA, 2002. [Google Scholar]
- Vieira, F.C.S.; Nahas, E. Comparison of microbial numbers in soils by using various culture media and temperatures. Microbiol. Res. 2005, 160, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Ottow, J.C.; Glathe, H. Rose bengal-malt extract-agar, a simple medium for the simultaneous isolation and enumeration of fungi and actinomycetes from soil. Appl. Microbiol. 1968, 16, 170–171. [Google Scholar] [PubMed]
- Van Schothorst, M.; Renaud, M. Malachite green pre-enrichment medium for improved Salmonella isolation from heavily contaminated samples. J. Appl. Bacteriol. 1985, 59, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Goodfellow, E.S.M., Stackebrant, E., Eds.; John Wiley & Sons Ltd.: London, UK, 1991; pp. 115–175. [Google Scholar]
- González, J.M.; Saiz-Jimenez, C. Microbial diversity in biodeteriorated monuments as studied by denaturing gradient gel electrophoresis. J. Sep. Sci. 2004, 27, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microb. 1993, 59, 695–700. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; The University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Pielou, E. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 166, 688. [Google Scholar] [CrossRef]
- Drijber, R.A.; Doran, J.W.; Parkhurst, A.M.; Lyon, D.J. Changes in soil microbial community structure with tillage under long-term wheat-fallow management. Soil Biol. Biochem. 2000, 32, 1419–1430. [Google Scholar] [CrossRef]
- Yao, H.; He, Z.; Wilson, M.J.; Campbell, C.D. Microbial biomass and community structure in a sequence of soils with increasing fertility and changing land use. Microb. Ecol. 2000, 40, 223–237. [Google Scholar] [PubMed]
- Muñoz, A.; López-Piñeiro, A.; Regodón, J.A.; Ramírez, M. Determination of soil microbial community fluctuations by different techniques in a maize field. In Microorganisms in Industry and Environment, Proceedings of the III International Conference on Environmental, Industrial and Applied Microbiology, Lisbon, Portugal, 2–4 December 2009; Mendez-Vilas, A., Ed.; World Scientific Publishing: Singapore, 2010; pp. 31–35. [Google Scholar]
- More, M.I.; Herrick, J.B.; Silva, M.C.; Ghiorse, W.C.; Madsen, E.L. Quantitative cell lysis of indigenous microorganisms and rapid extraction of microbial DNA from sediment. Appl. Environ. Microb. 1994, 60, 1572–1580. [Google Scholar]
- Lewis, C.; Swindel, B.; Tanner, G. Species diversity and diversity profiles: Concept, measurement, and application to timber and range management. J. Range Manag. 1988, 41, 466–469. [Google Scholar] [CrossRef]
- Aslam, Z.; Yasir, M.; Yoon, H.S.; Jeon, C.O.; Chung, Y.R. Diversity of the bacterial community in the rice rhizosphere managed under conventional and no-tillage practices. J. Microbiol. 2013, 51, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Dell’Amico, E.; Mazzocchi, M.; Cavalca, L.; Allievi, L.; Andreoni, V. Assessment of bacterial community structure in a long-term copper-polluted ex-vineyard soil. Microbiol. Res. 2008, 163, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Lester, E.D.; Satomi, M.; Ponce, A. Microflora of extreme arid Atacama desert soils. Soil Biol. Biochem. 2007, 39, 704–708. [Google Scholar] [CrossRef]
- Muyzer, G.; Smalla, K. Application of denaturing gradient gel electrophoresis (DGGE) and temperature gradient gel electrophoresis (TGGE) in microbial ecology. Antonie Leeuwenhoek 1998, 73, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Nikolausz, M.; Sipos, R.; Révész, S.; Székely, A.; Márialigeti, K. Observation of bias associated with re-amplification of DNA isolated from denaturing gradient gels. FEMS Microbiol. Lett. 2005, 244, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Heuer, H.; Wieland, G.; Schönfeld, J.; Schönwälder, A.; Gomes, N.C.M.; Smalla, K. Bacterial community profiling using DGGE or TGGE analysis. In Environmental Molecular Microbiology: Protocols and Applications; Rouchelle, P., Ed.; Horizon Scientific Press: Wymondham, UK, 2001; pp. 177–190. [Google Scholar]
- Mele, P.M.; Crowley, D.E. Application of self-organizing maps for assessing soil biological quality. Agric. Ecosyst. Environ. 2008, 126, 139–152. [Google Scholar] [CrossRef]
- Xiong, W.; Xie, P.; Wang, S.; Niu, Y.; Yang, X.; Chen, W. Sources of organic matter affect depth-related microbial community composition in sediments of Lake Erhai, Southwest China. J. Limnol. 2015, 74, 310–323. [Google Scholar] [CrossRef]
- Murata, T.; Tanaka, H.; Yasue, S.; Hamada, R.; Sakagami, K.; Kurokawa, Y. Seasonal variations in soil microbial biomass content and soil neutral sugar composition in grassland in the Japanese temperate zone. Appl. Soil Ecol 1999, 11, 253–259. [Google Scholar] [CrossRef]
- Shi, Y.; Lalande, R.; Hamel, C.; Ziadi, N.; Gagnon, B.; Hu, Z. Seasonal variation of microbial biomass, activity, and community structure in soil under different tillage and phosphorus management practices. Biol. Fert. Soils 2013, 49, 803–818. [Google Scholar] [CrossRef]
- Lynch, J.M.; Whipps, J.M. Substrate flow in the rhizosphere. Plant Soil 1990, 129, 1–10. [Google Scholar] [CrossRef]
- Butler, J.L.; Bottomley, P.J.; Griffith, S.M.; Myrold, D.D. Distribution and turnover of recently fixed photosynthate in ryegrass rhizospheres. Soil Biol. Biochem. 2004, 36, 371–382. [Google Scholar] [CrossRef]
- Gotelli, N.; Colwell, R. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- Hill, T.C.J.; Walsh, K.A.; Harris, J.A.; Moffett, B.F. Using ecological diversity measures with bacterial communities. FEMS Microbiol. Ecol. 2003, 43, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Embarcadero-Jiménez, S.; Flor, N.R.-O.; Wang, E.T. Bacterial communities estimated by pyrosequencing in the soils of chinampa, a traditional sustainable agro-ecosystem in Mexico. J. Soils Sediments 2016, 16, 1001–1011. [Google Scholar] [CrossRef]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G. An underground revolution: Biodiversity and soil ecological engineering for agricultural sustainability. Trends Ecol. Evol. 2016, 31, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Keesstra, S.D.; Bouma, J.; Wallinga, J.; Tittonell, P.; Smith, P.; Cerda, A.; Montanarella, L.; Quinton, J.N.; Pachepsky, Y.; van der Putten, W.H.; et al. The significance of soils and soil science towards realization of the United Nations Sustainable Development Goals. Soil 2016, 2, 111. [Google Scholar] [CrossRef]
- Wolff, F.; Timo, K. The UN Convention on biological diversity and soils: Status and future options. In International Yearbook of Soil Law and Policy 2016; Springer: Cham, Switzerland, 2017; pp. 129–148. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus/Type of Microorganism | First Year | Second Year | Third Year | Average | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sp | Su | Au | Wi | Sp | Su | Au | Wi | Sp | Su | Au | Wi | ||
| Azotobacter (1) ○ | 0.3aA | 0.6bA | 0.3aB | 0.4aA | 0.3abA | 0.5cA | 0.2aA | 0.5bcA | 0.3aA | 0.8cA | 0.3aAB | 0.5bA | 0.4 |
| Acinetobacter (2) ○ | 1.3aA | 9.1bB | 11.2bB | 18.2cA | 1.8aA | 9.2bB | 13.3cB | 19.5dA | 4.1abB | 1.2aA | 7.2bA | 15.6cA | 9.3 |
| Burkholderia (1) ○ | 11.7bA | 5.1aA | 4.2aB | 5.3aB | 10.6cA | 8.8bB | 5.8aB | 7.2bB | 12.2cA | 5.8bA | 2.9aA | 2.6aA | 6.9 |
| Chryseobacterium (1) ○ | 2.6cB | 1.5bA | 0.7aA | 1.6bB | 2.0bB | 7.2cB | 0.8aA | 2.2bC | 1.0bA | 0.2aA | 0.9bA | 0.8bA | 1.8 |
| Janthinobacterium (1) ○ | 0.0aA | 0.0aA | 0.0aA | 0.2bA | 0.0aA | 0.0aA | 0.0aA | 0.3A | 0.0aA | 0.0aA | 0.0aA | 0.2bA | 0.1 |
| Klebsiella (1) ○ | 0.2bA | 0.0aA | 0.0aA | 0.0aA | 0.3abA | 0.0aA | 1.1bB | 1.4bB | 0.6bA | 0.4bB | 0.0aA | 0.0aA | 0.3 |
| Pantoea (1) ○ | 1.6cB | 0.7bA | 0.3aA | 0.7bB | 0.9aA | 3.2bB | 0.5aA | 1.0aB | 1.4bB | 0.1aA | 0.4aA | 0.3aA | 0.9 |
| Pseudomonas (9) ○ | 40.1cB | 21.6bA | 12.7aA | 19.5bB | 30.0cA | 16.1aA | 21.3bB | 12.0aA | 42.4bB | 27.6aA | 20.0aB | 24.4aB | 24.0 |
| Serratia (1) ○ | 1.7bB | 0.9bA | 0.1aA | 0.4abA | 0.1aA | 0.4cA | 0.5cA | 0.1bA | 0.1aA | 0.3abA | 0.1aA | 0.6bA | 0.4 |
| Stenotrophomonas (1) ○ | 6.3bA | 2.7aB | 32.9dA | 16.5cA | 5.9aA | 2.4aB | 32.3cA | 12.9bA | 6.4bA | 1.7aA | 38.9dA | 13.9cA | 14.4 |
| Arthrobacter (2) ● | 7.3aA | 18.9bA | 18.4bB | 9.0aA | 22.6cB | 17.0bA | 6.5aA | 11.9abB | 6.6aA | 21.3bA | 7.3aA | 11.1aB | 13.2 |
| Bacillus (6) ● | 8.9aA | 15.9bA | 12.9bB | 8.4aA | 11.6bA | 11.9bA | 7.5aA | 16.4cB | 9.1aA | 18.0cA | 10.0aA | 12.7bB | 11.9 |
| Microbacterium (1) ● | 0.0aA | 0.0aA | 0.0aA | 0.0aA | 0.0aA | 0.0aA | 0.0aA | 0.0aA | 0.2bB | 0.0aA | 0.0aA | 0.0aA | 0.0 |
| Micromonospora (1) ● | 0.4bA | 0.6cA | 0.3aA | 0.7cA | 0.8aB | 2.9bB | 0.7aA | 0.9aA | 0.4bA | 0.1aA | 0.4bA | 0.3bA | 0.7 |
| Paenibacillus (1) ● | 4.7cA | 2.0bA | 0.9aA | 2.1bA | 4.7bA | 2.6aA | 2.3aB | 2.9aA | 4.3cA | 2.3bA | 1.2aA | 2.0bA | 2.7 |
| Rhodococcus (1) ● | 1.3abB | 2.0cB | 1.0aA | 1.5abB | 0.4aA | 0.0aA | 1.8bA | 0.0aA | 0.6aA | 2.4cB | 1.6bA | 1.1abB | 1.1 |
| Staphylococcus (1) ● | 0.0aA | 0.0aA | 0.1bA | 0.4cA | 0.0aA | 0.0aA | 0.4bA | 0.2bA | 0.0aA | 0.0aA | 0.0aA | 0.2bA | 0.1 |
| Streptomyces (11) ● | 11.7bA | 18.5cA | 3.7aA | 14.6bA | 7.9aA | 16.7cA | 5.3aA | 10.3bA | 10.4aA | 18.6cA | 9.1aB | 13.0abA | 11.7 |
| Gram+ bacteria | 34.3 | 57.9 | 37.3 | 36.7 | 48 | 51.1 | 24.5 | 42.6 | 31.6 | 62.7 | 29.6 | 40.4 | 41.4 |
| Gram− bacteria | 65.8 | 42.2 | 62.4 | 62.8 | 51.9 | 47.8 | 75.8 | 57.1 | 68.5 | 38.1 | 70.7 | 58.9 | 58.5 |
| Nº bacteria b | 30.4 | 23.36 | 16.9 | 24.8 | 37.7 | 22.4 | 32.0 | 37.1 | 30.3 | 35.8 | 29.7 | 30.6 | 29.3 |
| Nº yeasts a | 0.0 | 0.0 | 0.0 | 0.0 | 0.3 | 0.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.2 | 0.1 |
| Nº moulds b | 3.86 | 3.81 | 2.59 | 2.86 | 2.66 | 3.16 | 2.49 | 4.37 | 3.13B | 3.66 | 2.0 | 2.91 | 3.1 |
| Nº microorganisms b | 34.3 | 27.1 | 19.5 | 27.7 | 30.4 | 25.5 | 34.5 | 41.5 | 33.4 | 39.5 | 31.7 | 33.5 | 31.6 |
| Nº DB | 33 | 32 | 34 | 33 | 35 | 29 | 34 | 33 | 37 | 33 | 30 | 33 | 33 |
| D | 15.9 | 15.5 | 6.9 | 12.7 | 13.0 | 12.9 | 7.7 | 13.4 | 15.9 | 14.5 | 5.7 | 13.5 | 12.3 |
| H | 3.0 | 3.0 | 2.5 | 2.9 | 2.9 | 2.9 | 2.8 | 2.9 | 3.1 | 3.0 | 2.5 | 3.0 | 2.9 |
| E | 0.8 | 0.8 | 0.7 | 0.8 | 0.8 | 0.8 | 0.7 | 0.8 | 0.8 | 0.8 | 0.7 | 0.8 | 0.8 |
| Annual D | 16.2 | 16.0 | 16.2 | ||||||||||
| Annual H | 3.1 | 3.0 | 3.0 | ||||||||||
| Annual E | 0.8 | 0.8 | 0.8 | ||||||||||
| Microorganism | Nº of Colonies Analysed | Error (%) | |
|---|---|---|---|
| s | d | ||
| Acinetobacter haemolyticus | 2 | 3 | 0 |
| Acinetobacter rhizosphaerae | 1 | 4 | 0 |
| Arthrobacter dextranolyticus | 2 | 4 | 0 |
| Arthrobacter nicotinovorans | 5 | 4 | 0 |
| Bacillus simplex | 1 | 7 | 12.5 |
| Bacillus subtilis | 5 | 3 | 0 |
| Bacillus thuringiensis | 1 | 6 | 14.3 |
| Bacillus weihenstephanensis | 1 | 4 | 0 |
| Chryseobacterium indologenes | 1 | 4 | 0 |
| Janthinobacterium lividum | 1 | 5 | 0 |
| Klebsiella trevisanii | 1 | 4 | 0 |
| Microbacterium oxydans | 1 | 4 | 0 |
| Pseudomonas jessenii | 2 | 6 | 12.5 |
| Pseudomonas mediterranea | 3 | 3 | 0 |
| Pseudomonas mosselii | 2 | 3 | 0 |
| Pseudomonas poae | 1 | 4 | 0 |
| Pseudomonas putida | 4 | 3 | 0 |
| Rhodococcus corynebacterioides | 1 | 6 | 28.6 |
| Serratia proteamaculans | 1 | 5 | 0 |
| Stenotrophomonas maltophilia | 2 | 5 | 14.3 |
| Streptomyces scabrisporus | 1 | 5 | 0 |
| Streptomyces violaceorubidus | 2 | 0 | 0 |
| Streptomyces flavovirens | 2 | 0 | 0 |
| Streptomyces xanthophaeus | 2 | 0 | 0 |
| Streptomyces ciscaucasicus | 2 | 0 | 0 |
| Streptomyces griseoaurantiacus | 2 | 0 | 0 |
| Streptomyces drozdowiczii | 2 | 0 | 0 |
| Streptomyces violascens | 2 | 0 | 0 |
| Streptomyces virginiae | 2 | 0 | 0 |
| Streptomyces carpaticus | 2 | 0 | 0 |
| Total accumulated error | 4.02 | ||
| Genus | Culture Media | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| TSA | SC | YEPD | AZO | MG | ME | PEP | BR | Mean | |
| Arthrobacter (2) ● | 8.6 | 9.9 | 5.4 | 5.7 | 0.0 | 10.6 | 4.4 | 8.3 | 6.6 |
| Bacillus (6) ● | 11.2 | 2.0 | 11.2 | 20.8 | 0.0 | 11.4 | 7.1 | 11.1 | 9.3 |
| Microbacterium (1) ● | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Micromonospora (1) ● | 0.5 | 0.0 | 0.4 | 0.0 | 0.0 | 0.0 | 1.1 | 0.0 | 0.3 |
| Paenibacillus (1) ● | 1.1 | 0.0 | 2.7 | 1.9 | 0.0 | 0.8 | 1.1 | 2.8 | 1.3 |
| Rhodococcus (1) ● | 2.1 | 3.9 | 0.0 | 1.9 | 0.0 | 1.6 | 1.1 | 2.8 | 1.7 |
| Staphylococcus (1) ● | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Streptomyces (11) ● | 4.3 | 44.7 | 1.9 | 0.0 | 0.0 | 1.6 | 2.7 | 11.1 | 8.3 |
| Acinetobacter (2) ○ | 7.0 | 20.4 | 5.4 | 5.7 | 0.0 | 1.6 | 6.0 | 0.0 | 5.8 |
| Azotobacter (1) ○ | 0.0 | 0.0 | 0.0 | 5.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.7 |
| Burkholderia (1) ○ | 3.2 | 0.0 | 1.2 | 22.6 | 0.0 | 3.3 | 2.2 | 0.0 | 4.1 |
| Chryseobacterium (1) ○ | 0.0 | 0.0 | 3.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.4 |
| Janthinobacterium (1) ○ | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Klebsiella (1) ○ | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Pantoea (1) ○ | 0.5 | 0.0 | 0.8 | 0.0 | 0.0 | 0.0 | 0.5 | 0.0 | 0.2 |
| Pseudomonas (9) ○ | 17.1 | 7.2 | 23.2 | 15.1 | 100 | 18.7 | 23.1 | 41.7 | 30.8 |
| Serratia (1) ○ | 0.0 | 0.0 | 0.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Stenotrophomonas (1) ○ | 44.4 | 11.8 | 44.0 | 20.8 | 0.0 | 50.4 | 50.5 | 22.2 | 30.5 |
| Total | |||||||||
| D | 9.57 | 10.2 | 9.41 | 4.61 | 1.34 | 9.32 | 9.37 | 4.42 | 12.53 |
| H | 2.36 | 2.82 | 2.26 | 1.48 | 0.83 | 2.24 | 2.18 | 1.36 | 2.98 |
| E | 0.74 | 0.88 | 0.73 | 0.77 | 0.86 | 0.76 | 0.78 | 0.77 | 0.91 |
| Nº DB | 19 | 27 | 18 | 9 | 2 | 15 | 17 | 6 | 36 |
| Nº UB | 0 | 11 | 2 | 4 | 0 | 0 | 0 | 0 | 17 |
| Variable | Culture Media Combination | ||||
|---|---|---|---|---|---|
| TSA/SC/YEPD | AZO/YEPD/SC | AZO/PEP/SC | TSA/SC/AZO | TSA/SC/PEP | |
| D | 11.6 | 12.2 | 11.7 | 12.0 | 11.4 |
| H | 2.61 | 2.72 | 2.54 | 2.67 | 2.49 |
| E | 0.80 | 0.81 | 0.80 | 0.81 | 0.78 |
| Nº DB | 32 | 36 | 33 | 34 | 30 |
| Nº UB | 13 | 17 | 15 | 15 | 11 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez, M.; López-Piñeiro, A.; Peña, D.; Rato Nunes, J.; Albarrán, Á.; Muñoz, A.; Gama, J.; Loures, L. Seasonal and Interannual Fluctuation of the Microbial Soil Community in a Maize Field under Long-Term Conservation Agriculture Management. Sustainability 2017, 9, 778. https://0-doi-org.brum.beds.ac.uk/10.3390/su9050778
Ramírez M, López-Piñeiro A, Peña D, Rato Nunes J, Albarrán Á, Muñoz A, Gama J, Loures L. Seasonal and Interannual Fluctuation of the Microbial Soil Community in a Maize Field under Long-Term Conservation Agriculture Management. Sustainability. 2017; 9(5):778. https://0-doi-org.brum.beds.ac.uk/10.3390/su9050778
Chicago/Turabian StyleRamírez, Manuel, Antonio López-Piñeiro, David Peña, José Rato Nunes, Ángel Albarrán, Ana Muñoz, José Gama, and Luis Loures. 2017. "Seasonal and Interannual Fluctuation of the Microbial Soil Community in a Maize Field under Long-Term Conservation Agriculture Management" Sustainability 9, no. 5: 778. https://0-doi-org.brum.beds.ac.uk/10.3390/su9050778