In Vivo Toxicity of Solasonine and Its Effects on cyp450 Family Gene Expression in the Livers of Male Mice from Four Strains

Abstract
:1. Introduction
2. Results
2.1. LD50 Values of Solasonine in Four Different Mouse Strains in the First Week after Treatment
2.2. Body Weight of Mice from Four Different Strains in the First Week after Intraperitoneal Injection of Solasonine
2.3. Ratios of Liver and Spleen Weight to Body Weight in Mice of Four Different Strains Six Days after Solasonine Injection
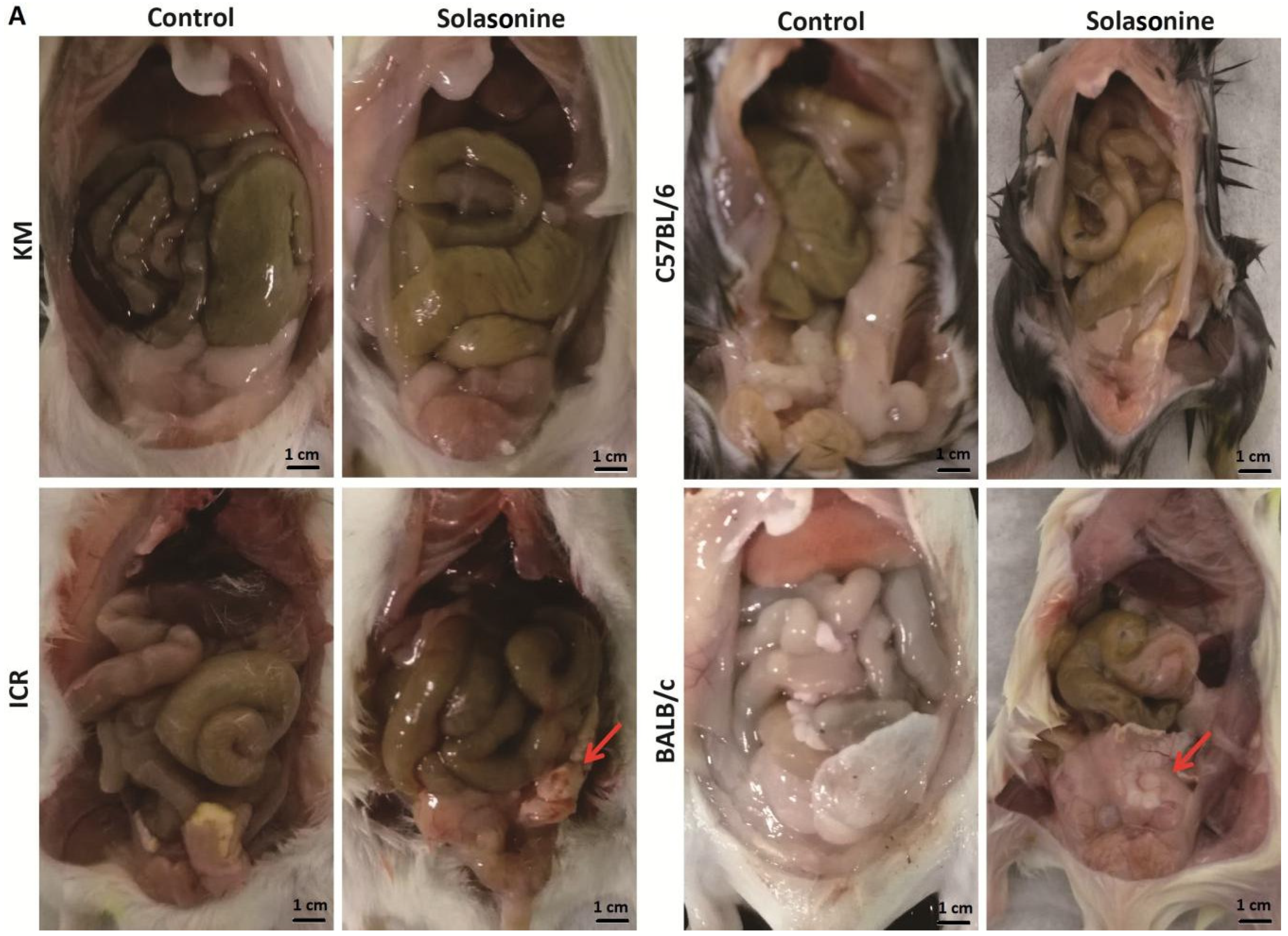
2.4. Pathological Changes in Mice Treated with Solasonine
2.5. mRNA Levels of Genes of the cyp450 Family in the Livers of Each Strain of Mice Six Days after Intraperitoneal Injection with Solasonine at a Dose of 21.5 mg/kg
2.6. Protein Levels of the cyp450 Family Genes in the Livers of Mice from Four Strains Six Days after Intraperitoneal Injection with Solasonine at a Dose of 21.5 mg/kg
3. Discussion
4. Materials and Methods
4.1. Animals
4.1.1. Horn’s Assay (n = 5, 2.15×)
4.1.2. In Vivo Toxicity Experiment
4.2. Haematoxylin–Eosin Staining
4.3. RNA and Real-Time PCR
4.4. Protein and ELISA (Enzyme-Linked Immunosorbent Assay)
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, G.; Xie, Y.; Zhou, Y.; Xiang, C.; Chen, L.; Zhang, C.; Hou, X.; Chen, J.; Zong, H.; Liu, G. p53 pathway is involved in cell competition during mouse embryogenesis. Proc. Natl. Acad. Sci. USA. 2017, 114, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genvigir, F.D.; Salgado, P.C.; Felipe, C.R.; Luo, E.Y.; Alves, C.; Cerda, A.; Tedesco-Silva, H., Jr.; Medina-Pestana, J.O.; Oliveira, N.; Rodrigues, A.C.; et al. Influence of the CYP3A4/5 genetic score and ABCB1 polymorphisms on tacrolimus exposure and renal function in Brazilian kidney transplant patients. Pharmacogenet. Genom. 2016, 26, 462–472. [Google Scholar] [CrossRef] [PubMed]
- Gomez, M.B.; Gorla, D.E. Distribution of Pyrethroid Resistant Populations of Triatoma infestans in the Southern Cone of South America. Plos. Negl. Trop. Dis. 2016, 10, e4561. [Google Scholar]
- Park, M.; Sutherland, J.B.; Kim, J.N.; Rafii, F. Effect of fluoroquinolone resistance selection on the fitness of three strains of Clostridium perfringens. Microb. Drug Resist. 2013, 19, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Dass, S.B.; Heflich, R.H.; Casciano, D.A. The mutagenic response at the ouabain resistance locus in T cells of mice exposed to N-ethyl-N-nitrosourea parallels the response at the Hprt locus and correlates with mutation target size. Carcinogenesis 1997, 18, 2233–2237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oikonomou, E.; Papageorgiou, N.; Papaioannou, S.; Toutouzas, K.; Latsios, G.; Siasos, G.; Tousoulis, D. Genetic testing and antiplatelet treatment: Still way to go? Int. J. Cardiol. 2015, 187, 63–65. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zheng, X.; Liu, D.Y.; Zhao, Q.; Wu, Y.W.; Tan, F.L.; Wang, Y.X.; Jiang, J.; Hu, P. Therapeutic effects and adverse drug reactions are affected by icotinib exposure and CYP2C19 and EGFR genotypes in Chinese non-small cell lung cancer patients. Asian Pac. J. Cancer Prev. 2014, 15, 7195–7200. [Google Scholar] [CrossRef] [PubMed]
- Falck, D.; Rahimi, P.F.; Giera, M.; Honing, M.; Kool, J.; Niessen, W.M. Comparison of (bio-)transformation methods for the generation of metabolite-like compound libraries of p38α MAP kinase inhibitors using high-resolution screening. J. Pharm. Biomed. Anal. 2014, 88, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Ding, X.; Zhang, Q.Y. An update on the role of intestinal cytochrome P450 enzymes in drug disposition. Acta Pharm. Sin. B. 2016, 6, 374–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivares, C.I.; Sierra-Alvarez, R.; Abrell, L.; Chorover, J.; Simonich, M.; Tanguay, R.L.; Field, J.A. Zebrafish embryo toxicity of anaerobic biotransformation products from the insensitive munitions compound 2,4-dinitroanisole (DNAN). Environ. Toxicol. Chem. 2016, 35, 2774–2781. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.Q.; Zhu, Y.P.; Pang, J.; Wang, Y.X.; Song, D.Q.; Kong, W.J.; Jiang, J.D. Tetrandrine potentiates the hypoglycemic efficacy of berberine by inhibiting P-glycoprotein function. Biol. Pharm. Bull. 2013, 36, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, K.; Sui, D.; Ouyang, Z.; Xu, H.; Wei, Y. Effects of tetrahydroberberine and tetrahydropalmatine on hepatic cytochrome P450 expression and their toxicity in mice. Chem. Biol. Interact. 2017, 268, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, Y.; Fan, X.; Tan, H.; Zeng, H.; Wang, Y.; Chen, P.; Huang, M.; Bi, H. Hepato-protective effect of resveratrol against acetaminophen-induced liver injury is associated with inhibition of CYP-mediated bioactivation and regulation of SIRT1-p53 signaling pathways. Toxicol. Lett. 2015, 236, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Fan, X.; Wang, Y.; Tan, H.; Chen, P.; Zeng, H.; Huang, M.; Bi, H. Hepato-protective effects of six schisandra lignans on acetaminophen-induced liver injury are partially associated with the inhibition of CYP-mediated bioactivation. Chem. Biol. Interact. 2015, 231, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R.; Zeldin, D.C.; Hoffman, S.M.; Maltais, L.J.; Wain, H.M.; Nebert, D.W. Comparison of cytochrome P450 (CYP) genes from the mouse and human genomes, including nomenclature recommendations for genes, pseudogenes and alternative-splice variants. Pharmacogenetics 2004, 14, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Cytochrome P450 research and The Journal of Biological Chemistry. J. Biol. Chem. 2018, Jbc. [Google Scholar] [CrossRef] [PubMed]
- Renaud, H.J.; Cui, J.Y.; Khan, M.; Klaassen, C.D. Tissue distribution and gender-divergent expression of 78 cytochrome P450 mRNAs in mice. Toxicol. Sci. 2011, 124, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, Q.; Li, X.; Gong, X.; Ruan, Y.; Zeng, S.; Lu, L.; Qi, X.; Wang, Y.; Hu, M.; Zhu, L.; et al. Tissue Distribution and Gender-Specific Protein Expression of Cytochrome P450 in five Mouse Genotypes with a Background of FVB. Pharm. Res. 2018, 35, 114. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Yang, J.; Liu, H.; Lin, F.Q.; Shi, J.S.; Zhang, F. Effects of tetrahydroxystilbene glucoside on mouse liver cytochrome P450 enzyme expressions. Xenobiotica 2015, 45, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Jarrar, Y.B.; Al-Essa, L.; Kilani, A.; Hasan, M.; Al-Qerem, W. Alterations in the gene expression of drug and arachidonic acid-metabolizing Cyp450 in the livers of controlled and uncontrolled insulin-dependent diabetic mice. Diabetes Metab. Syndr. Obes. 2018, 11, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Maria, K. Psychophysiological stress: A significant parameter in drug pharmacokinetics. Expert Opin. Drug Met. Toxicol. 2013, 9, 1317–1334. [Google Scholar]
- Kwak, H.C.; Kim, H.C.; Oh, S.J.; Kim, S.K. Effects of age increase on hepatic expression and activity of cytochrome P450 in male C57BL/6 mice. Arch. Pharm. Res. 2015, 38, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Zhu, F.; Yang, Y.; Li, M. Purification, antitumor activity in vitro of steroidal glycoalkaloids from black nightshade (Solanum nigrum L.). Food Chem. 2013, 141, 1181–1186. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, S.; Jin, D.; Zhang, J.; Xu, P.; Chen, L. Inhibitive effect on growth of lung cancer cell of solasonine extracted from solanum nigrum Linn, an anticancer used in Traditional Chinese Medicine. Lishizhen Med. Materia Med. Res. 2015, 2, 333–334. [Google Scholar]
- Wang, X.; Zou, S.; Lan, Y.; Xing, J.; Lan, X.; Zhang, B. Solasonine inhibits glioma growth through anti-inflammatory pathways. Am. J. Transl. Res. 2017, 9, 3977–3989. [Google Scholar] [PubMed]
- Munari, C.C.; de Oliveira, P.F.; Campos, J.C.; Martins Sde, P.; Da Costa, J.C.; Bastos, J.K.; Tavares, D.C. Antiproliferative activity of Solanum lycocarpum alkaloidic extract and their constituents, solamargine and solasonine, in tumor cell lines. J. Nat. Med. 2014, 68, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Akter, R.; Uddin, S.J.; Tiralongo, J.; Grice, I.D.; Tiralongo, E. A New Cytotoxic Steroidal Glycoalkaloid from the Methanol Extract of Blumea lacera Leaves. J. Pharm. Sci. 2015, 18, 616–633. [Google Scholar] [CrossRef]
- Huang, W.; Wang, Y.; Zhu, H.; Wu, Y.; Xie, X.; Wang, D. Solasonine-induced Apoptosis in Lung Cancer Cell Line H446 and Its Mechanism. Zhongguo Fei Ai Za Zhi 2015, 18, 416–421. [Google Scholar] [PubMed]
- Yan, Z.; Wang, X.; Xu, T.; An, Z.; Zhang, X.; Zhu, F. Apoptosis-inducing Effects of Solasonine on Human Cholangiocarcinoma Epithelial QBC939 Cells and Its Mechanism. Nat. Prod. Res. Dev. 2017, 4, 559–562. [Google Scholar]
- Chen, L.; Hu, J.; Li, S.-S.; Zhan, J.; Xu, P.; Chen, L. The inhibitory effect of solasonine on melanoma cells. Jiangxi J. Trad. Chin. Med. 2014, 29–30. [Google Scholar]
- Yang, J.; Huang, W.; Tan, W. Solasonine, a Natural Glycoalkaloid Compound, Inhibits Gli-Mediated Transcriptional Activity. Molecules 2016, 21, 1364. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xie, E.; Li, S.; Liu, Z.; Luo, X.; Xu, P. Introduction of mice with spontaneous breast cancer and its application in the evaluation of the anticancer effect of solasonine. Heilongjiang Animal Sci. Vet. Med. 2016, 21, 32–34. [Google Scholar]
- Li, N.; Ma, L. Food Toxicology; China Agricultural University Press: Beijing, China, 2016; pp. 127–129. [Google Scholar]
- Chen, Y.; Zhang, S.; Chen, D.; Zhou, M.; Zheng, J.; Xiang, Z. An UPLC-MS/MS method for determination of solasonine in rat plasma and its application of a pharmacokinetic and bioavailability study. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2015, 985, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Su, Y.; Chang, S.; Sun, Y.; Meng, X.; Shan, A. Vitamin C protects piglet liver against zearalenone-induced oxidative stress by modulating expression of nuclear receptors PXR and CAR and their target genes. Food Funct. 2017, 8, 3675–3687. [Google Scholar] [CrossRef] [PubMed]
- Gorman, N.; Trask, H.S.; Robinson, S.W.; Sinclair, J.F.; Gerhard, G.S.; Smith, A.G.; Sinclair, P.R. Hexachlorobenzene stimulates uroporphyria in low affinity AHR mice without increasing CYP1A2. Toxicol. Appl. Pharmacol. 2007, 221, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, R.S.; Friedrich, K.; De-Oliveira, A.C.A.X.; Suarez-Kurtz, G.; Paumgarttena, F.J.R. Malaria downmodulates mRNA expression and catalytic activities of CYP1A2, 2E1 and 3A11 in mouse liver. Eur. J. Pharmacol. 2009, 616, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Harkitis, P.; Daskalopoulos, E.P.; Malliou, F.; Lang, M.A.; Marselos, M.; Fotopoulos, A.; Albucharali, G.; Konstandi, M. Dopamine D2-Receptor Antagonists Down-Regulate CYP1A1/2 and CYP1B1 in the Rat Liver. PLoS ONE 2015, 10, e0128708. [Google Scholar] [CrossRef] [PubMed]
- Rysz, M.; Bromek, E.; Haduch, A.; Sadakierska-Chudy, A.; Daniel, W.A. Damage to the Brain Serotonergic System Increases the Expression of Liver Cytochrome P450. Drug Metab. Dispos. 2015, 43, 1345–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochańska-Dziurowicz, A.A.; Janikowska, G.; Bijak, A.; Stanjek-Cichoracka, A.; Mazurek, U. The effect of maximal physical exercise on relationships between the growth hormone (GH) and insulin-like growth factor 1 (IGF-1) and transcriptional activity of CYP1A2 in young ice hockey players. J. Sports Med. Phys. Fit. 2015, 55, 158–163. [Google Scholar]
- Kot, M.; Daujat-Chavanieu, M. The impact of serotonergic system dysfunction on the regulation of P4501A isoforms during liver insufficiency and consequences for thyroid hormone homeostasis. Food Chem. Toxicol. 2016, 97, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Hanada, K.; Nakai, K.; Tanaka, H.; Suzuki, F.; Kumada, H.; Ohno, Y.; Ozawa, S.; Ogata, H. Effect of nuclear receptor downregulation on hepatic expression of cytochrome P450 and transporters in chronic hepatitis C in association with fibrosis development. Drug Metab. Pharmacokinet. 2012, 27, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Uno, Y.; Osada, N. CpG site degeneration triggered by the loss of functional constraint created a highly polymorphic macaque drug-metabolizing gene, CYP1A2. BMC Evol. Biol. 2011, 11, 283. [Google Scholar] [CrossRef] [PubMed]
- Ghotbi, R.; Gomez, A.; Milani, L.; Tybring, G.; Syvänen, A.C.; Bertilsson, L.; Ingelmansundberg, M.; Aklillu, E. Allele-specific expression and gene methylation in the control of CYP1A2 mRNA level in human livers. Pharmacogenomics J. 2009, 9, 208–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrzal, R.; Daujat-Chavanieu, M.; Pascussi, J.M.; Ulrichova, J.; Maurel, P.; Dvorak, Z. Microtubules-interfering agents restrict aryl hydrocarbon receptor-mediated CYP1A2 induction in primary cultures of human hepatocytes via c-jun-N-terminal kinase and glucocorticoid receptor. Eur. J. Pharmacol. 2008, 581, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Kakan, X.; Chen, P.; Zhang, J. Clock gene mPer2 functions in diurnal variation of acetaminophen induced hepatotoxicity in mice. Exp. Toxicol. Pathol. 2011, 63, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Sekine, S.; Lan, B.Y.; Bedolli, M.; Feng, S.; Hebrok, M. Liver-specific loss of beta-catenin blocks glutamine synthesis pathway activity and cytochrome p450 expression in mice. Hepatology 2006, 43, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Tomasi, M.L.; Ramani, K.; Ryoo, M.; Cossu, C.; Floris, A.; Murray, B.J.; Iglesias-Ara, A.; Spissu, Y.; Mavila, N. SUMOylation regulates cytochrome P450 2E1 expression and activity in alcoholic liver disease. FASEB J. 2018, 32, 3278–3288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitam, V.O.; Maksymchuk, O.V.; Chashchyn, M.O. The possible mechanisms of CYP2E1 interactions with HSP90 and the influence of ethanol on them. BMC Struct. Biol. 2012, 12, 33. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | KM | ICR | C57BL/6 | BALB/c | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dose (mg/kg) | 100 | 46.4 | 21.5 | 10 | 100 | 46.4 | 21.5 | 10 | 100 | 46.4 | 21.5 | 10 | 100 | 46.4 | 21.5 | 10 |
| Death Rate | 2/5 | 2/5 | 0/5 | 0/5 | 5/5 | 3/5 | 0/5 | 0/5 | 4/5 | 4/5 | 1/5 | 0/5 | 5/5 | 4/5 | 1/5 | 0/5 |
| LD50 (mg/kg) | >43 | 43 | 34.8 | 31.6 | ||||||||||||
| 95% Confidence Limit | N/A | 29.5–62.6 | 19.2–63 | 20.5–48.8 | ||||||||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, Y.; Li, S.; Chen, L.; Liu, Z.; Luo, X.; Xu, P.; Chen, L. In Vivo Toxicity of Solasonine and Its Effects on cyp450 Family Gene Expression in the Livers of Male Mice from Four Strains. Toxins 2018, 10, 487. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10120487
Zhong Y, Li S, Chen L, Liu Z, Luo X, Xu P, Chen L. In Vivo Toxicity of Solasonine and Its Effects on cyp450 Family Gene Expression in the Livers of Male Mice from Four Strains. Toxins. 2018; 10(12):487. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10120487
Chicago/Turabian StyleZhong, Youbao, Shanshan Li, Liling Chen, Zhiyong Liu, Xiaoquan Luo, Peng Xu, and Lai Chen. 2018. "In Vivo Toxicity of Solasonine and Its Effects on cyp450 Family Gene Expression in the Livers of Male Mice from Four Strains" Toxins 10, no. 12: 487. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10120487