Subclinical Lipopolysaccharide from Salmonella Enteritidis Induces Dysregulation of Bioactive Substances from Selected Brain Sections and Glands of Neuroendocrine Axes


{kind=link}
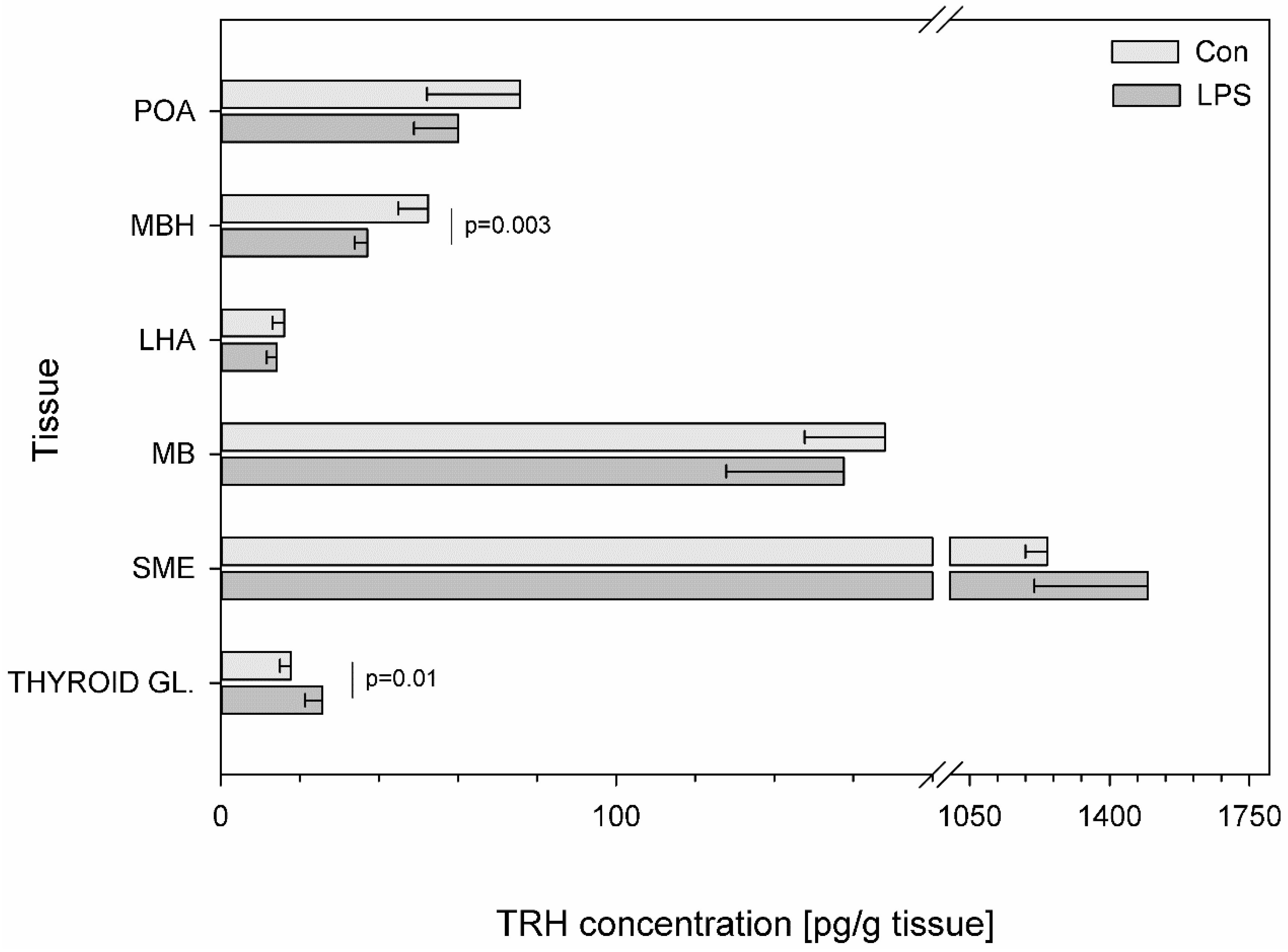{kind=link}
{kind=link}
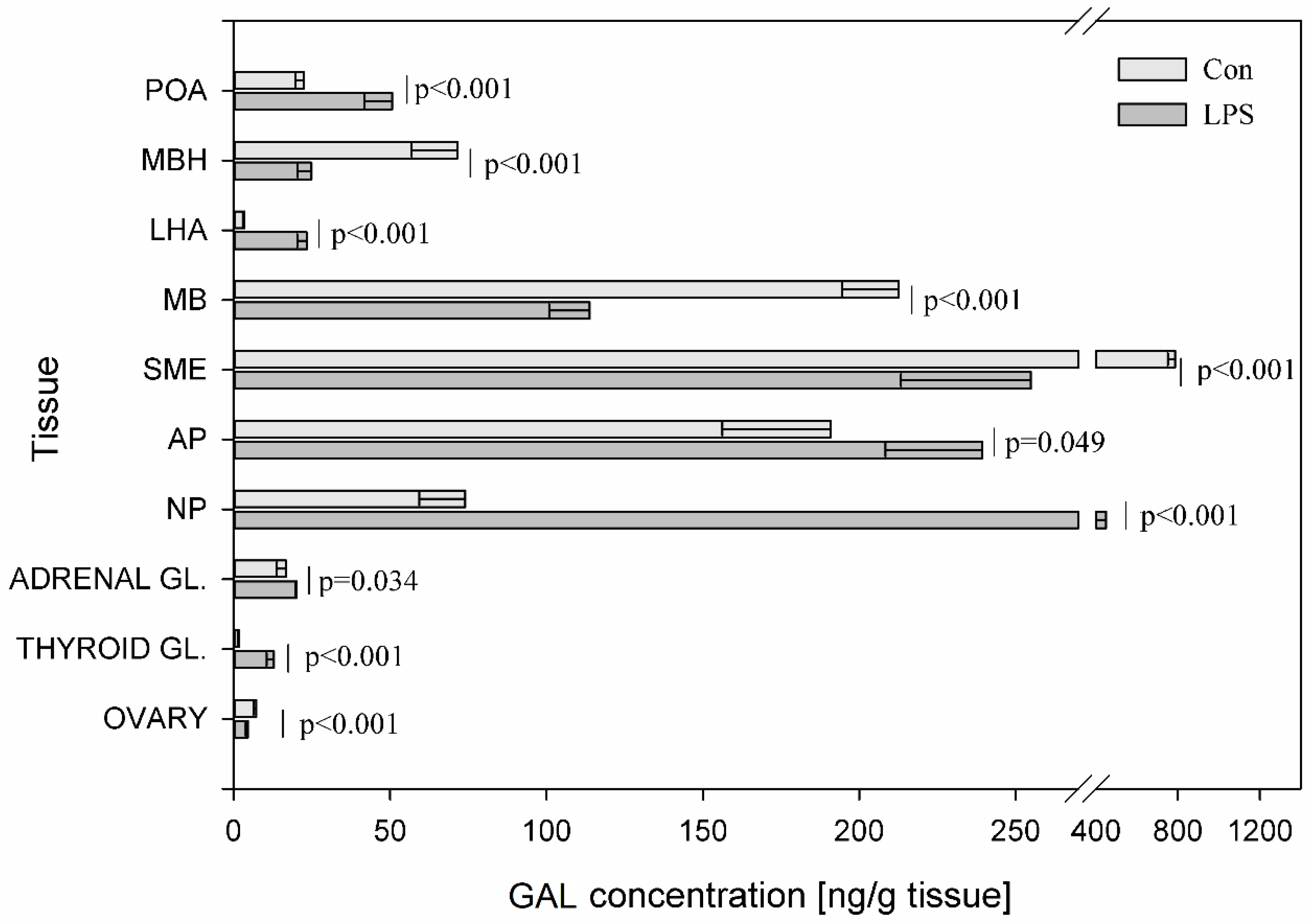{kind=link}
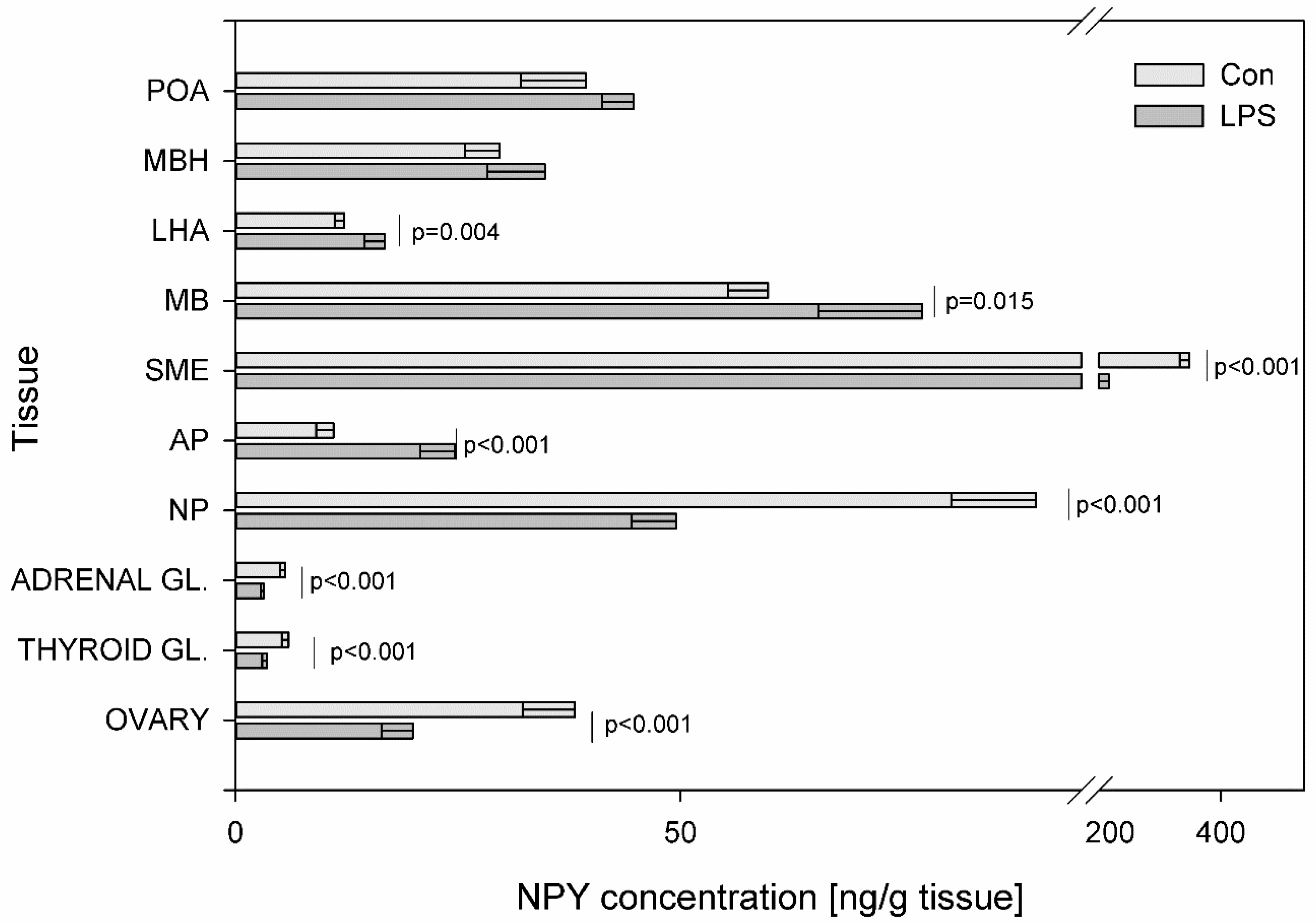{kind=link}
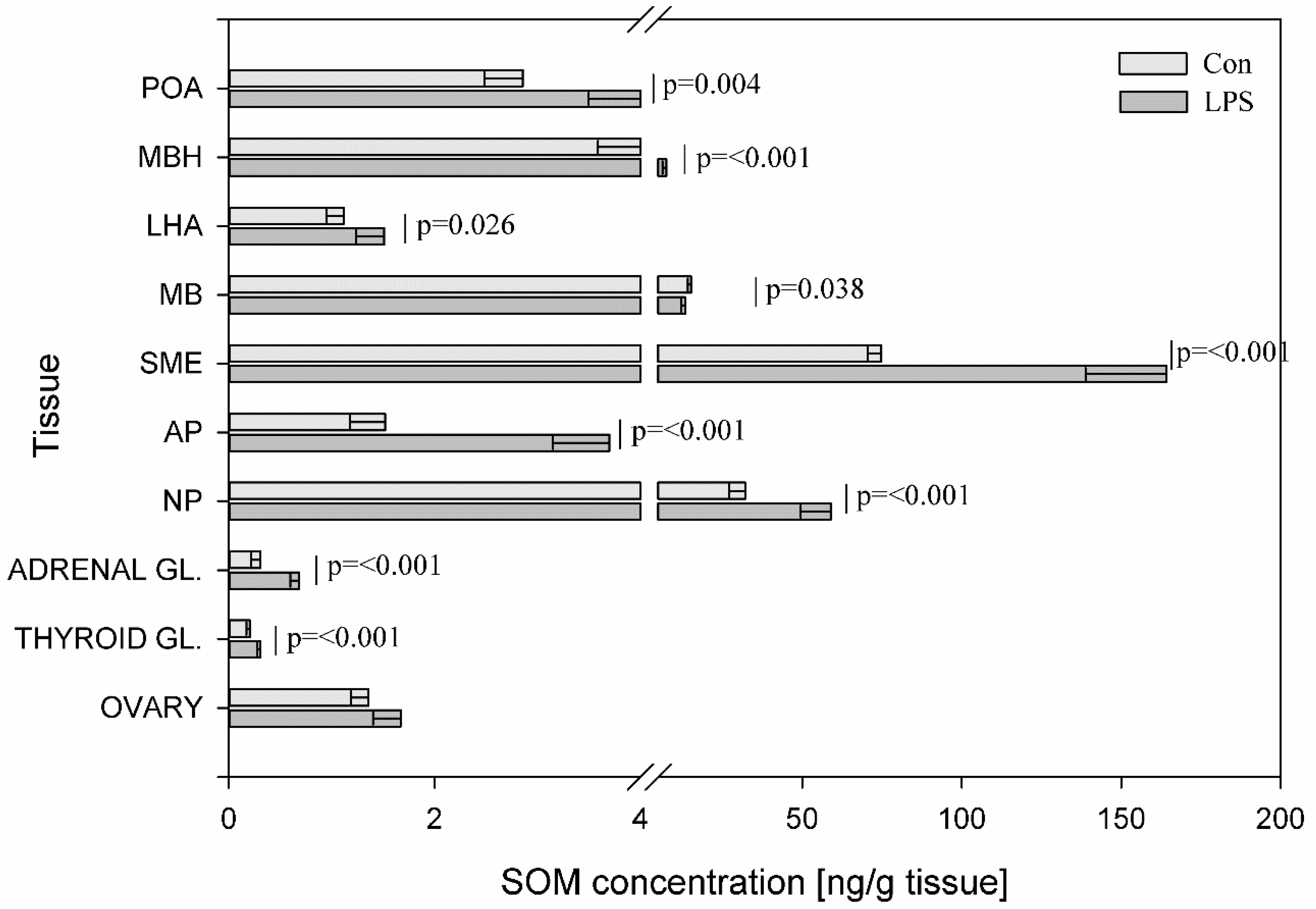{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animal Housing and LPS Administration
5.2. Tissue Collection
5.3. Brain Peptide Extraction from the Tissues
5.3.1. Sample Preparation and High-temperature Extraction
5.3.2. Solid-phase Extraction and Concentration
5.4. Enzyme-Linked Immunosorbent Assay for Quantitative Determination of Selected Peptides (CRH, TRH, GnRH, GAL, NPY, SOM, SP, VIP) in Tissue Extracts
5.5. Statistical Analysis
5.6. Ethics Statement
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Knoll, L.J.; Hogan, D.A.; Leong, J.M.; Heitman, J.; Condit, R.C. Pearls collections: What we can learn about infectious disease and cancer. PLoS Pathog. 2018, 14, e1006915. [Google Scholar] [CrossRef] [PubMed]
- Vedham, V.; Divi, R.L.; Starks, V.L.; Verma, M. Multiple Infections and Cancer: Implications in Epidemiology. Technol. Cancer Res. Treat. 2014, 13, 177–194. [Google Scholar] [CrossRef] [PubMed]
- Mesri, E.A.; Feitelson, M.A.; Munger, K. Human Viral Oncogenesis: A Cancer Hallmarks Analysis. Cell Host. Microbe 2014, 15, 266–282. [Google Scholar] [CrossRef] [PubMed]
- Mikołajczyk, A. Invited Brief Commentary on the Article “Breast Cancer Association with Cytomegalo Virus—A Tertiary Center Case-Control Study” Is Cytomegalo Virus a Breast Cancer Etiologic Risk Factor? J. Investig. Surg. 2017, 20, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Zloza, A. Viruses, bacteria, and parasites—Oh my! a resurgence of interest in microbial-based therapy for cancer. J. Immunother. Cancer 2018, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Garrett, W.S. Cancer and the microbiota. Science 2015, 348, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Matson, V.; Fessler, J.; Bao, R.; Chongsuwat, T.; Zha, Y.; Alegre, M.L.; Luke, J.J.; Gajewski, T.F. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science 2018, 359, 104–108. [Google Scholar] [CrossRef]
- Routy, B.; Le Chatelier, E.; Derosa, L.; Duong, C.P.M.; Alou, M.T.; Daillere, R.; Fluckiger, F.A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut microbiome influences efficacy of P-1-based immunotherapy against epithelial tumors. Science 2018, 359, 97–103. [Google Scholar] [CrossRef]
- Kaimala, S.; Al-Sbiei, A.; Cabral-Marques, O.; Fernandez-Cabezudo, M.J.; Al-Ramadi, B.K. Attenuated Bacteria as immunotherapeutic tools for cancer treatment. Front. Oncol. 2018, 8, 136. [Google Scholar] [CrossRef]
- Kocijancic, D.; Leschner, S.; Felgner, S.; Komoll, R.; Frahm, M.; Pawar, V.; Weiss, S. Therapeutic benefit of Salmonella attributed to LPS and TNF-α is exhaustible and dictated by tumor susceptibility. Oncotarget 2017, 8, 36492–36508. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chu, C.-H.; Qian, L.; Chen, S.-H.; Wilson, B.; Oyarzabal, E.; Jiang, L.; Ali, S.; Robinson, B.; Kim, H.-C.; et al. Substance P Exacerbates Dopaminergic Neurodegeneration through Neurokinin-1 Receptor-Independent Activation of Microglial NADPH Oxidase. J. Neurosci. 2014, 34, 12490–12503. [Google Scholar] [CrossRef] [PubMed]
- Maciel, B.M.; Passos, R.; Sriranganathan, N. Salmonella enterica: Latency. In Current Topics in Salmonella and Salmonellosis; Mares, M., Ed.; InTechOpen: Rijeka, Croatia, 2017; pp. 43–58. [Google Scholar]
- Marzel, A.; Desai, P.T.; Goren, A.; Schorr, Y.I.; Nissan, I.; Porwollik, S.; Valinsky, L.; Mcclelland, M.; Rahav, G.; Gal-Mor, O. Persistent infections by non-typhoidal salmonella in humans: Epidemiology and genetics. Clin. Infect. Dis. 2016, 62, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Mikołajczyk, A.; Radkowski, M. Contamination of Salmonella spp. in slaughter chickens. Med. Wet. 2001, 57, 745–747. [Google Scholar]
- Mikołajczyk, A. An evaluation of the influence of trisodium phosphate and duration of storage upon the survival rate of Salmonella bacteria in turkey carcasses. In Salmonella—Distribution, Adaptation, Control Measures and Molecular Technologies; Chapter 7; Annous, B.A., Gurtler, J.B., Eds.; InTechOpen: Rijeka, Croatia, 2012; pp. 147–158. [Google Scholar]
- Mikołajczyk, A. Evaluation of the effects of a mixture of organic acids and duration of storage on the survival of Salmonella on turkey carcasses. J. Food Prot. 2015, 78, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Suárez, E.J.; Selem-Mojica, N.; Ortiz-López, R.; Gebreyes, W.A.; Allard, M.W.; Barona-Gómez, F.; Rubio-Lozano, M.S. Whole genome sequencing reveals widespread distribution of typhoidal toxin genes and VirB/D4 plasmids in bovine-associated nontyphoidal Salmonella. Sci. Rep. 2018, 8, 9864. [Google Scholar] [CrossRef]
- Iyer, P.; Barreto, S.G.; Sahoo, B.; Chandrani, P.; Ramadwar, M.R.; Shrikhande, S.V.; Dutt, A. Non-typhoidal Salmonella DNA traces in gallbladder cancer. Infect. Agent Cancer 2016, 11, 2–4. [Google Scholar] [CrossRef]
- Steimle, A.; Autenrieth, I.B.; Frick, J.S. Structure and function: Lipid A modifications in commensals and pathogens. Int. J. Med. Microbiol. 2016, 306, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C.; Trent, M.S. Biosynthesis and Export of Bacterial Lipopolysaccharides. Annu. Rev. Biochem. 2014, 83, 99–128. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, E.; Rother, N.; Yanginlar, C.; Hilbrands, L.B.; van der Vlag, J. Neutrophils discriminate between lipopolysaccharides of different bacterial sources and selectively release neutrophil extracellular traps. Front. Immunol. 2016, 7, 484. [Google Scholar] [CrossRef]
- Reyes, R.E.; González, C.; Jiménez, R.C.; Ortiz, M.C.; Andrade, A.A. Mechanisms of O-Antigen Structural Variation of Bacterial Lipopolysaccharide (LPS). In World of Polysaccharides; Karunaratne, D.N., Ed.; IntechOpen: Rijeka, Croatia, 2012; pp. 71–95. [Google Scholar]
- Vatanen, T.; Kostic, A.D.; D’Hennezel, E.; Siljander, H.; Franzosa, E.A.; Yassour, M.; Kolde, R.; Vlamakis, H.; Arthur, T.D.; Hämäläinen, A.M.; et al. Variation in Microbiome LPS Immunogenicity Contributes to Autoimmunity in Humans. Cell 2016, 165, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Bryant, C.E.; Spring, D.R.; Gangloff, M.; Gay, N.J. The molecular basis of the host response to lipopolysaccharide. Nat. Rev. Microbiol. 2010, 8, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Nedrebø, T.; Reed, R.K. Different serotypes of endotoxin (lipopolysaccharide) cause different increases in albumin extravasation in rats. Shock 2002, 18, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B.; Kumar, P.; Cutler, C.W.; Mohamadzadeh, M.; Van Dyke, T.; Banchereau, J. Lipopolysaccharides from Distinct Pathogens Induce Different Classes of Immune Responses In Vivo. J. Immunol. 2001, 167, 5067–5076. [Google Scholar] [CrossRef] [PubMed]
- Mikołajczyk, A.; Kozłowska, A.; Gonkowski, S. Distribution and Neurochemistry of the Porcine Ileocaecal Valve Projecting Sensory Neurons in the Dorsal Root Ganglia and the Influence of Lipopolysaccharide from Different Serotypes of Salmonella spp. on the Chemical Coding of DRG Neurons in the Cell Cultures. Int. J. Mol. Sci. 2018, 19, 2551. [Google Scholar]
- Kang, M.; Edmundson, P.; Araujo-Perez, F.; McCoy, A.N.; Galanko, J.; Keku, T.O. Association of plasma endotoxin, inflammatory cytokines and risk of colorectal adenomas. BMC Cancer 2013, 13, 91. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.K.; Yum, K.S. Association of endotoxins and colon polyp: A case-control study. J. Korean Med. Sci. 2012, 27, 1062–1065. [Google Scholar] [CrossRef]
- Ramachandran, G. Gram-positive and gram-negative bacterial toxins in sepsis: A brief review. Virulence 2014, 5, 213–218. [Google Scholar] [CrossRef]
- Hoban, D.B.; Connaughton, E.; Connaughton, C.; Hogan, G.; Thornton, C.; Mulcahy, P.; Moloney, T.C.; Dowd, E. Further characterisation of the LPS model of Parkinson’s disease: A comparison of intra-nigral and intra-striatal lipopolysaccharide administration on motor function, microgliosis and nigrostriatal neurodegeneration in the rat. Brain Behav. Immun. 2013, 27, 91–100. [Google Scholar] [CrossRef]
- Huang, B.; Liu, J.; Ju, C.; Yang, D.; Chen, G.; Xu, S.; Zeng, Y.; Yan, X.; Wang, W.; Liu, D.; et al. Licochalcone A Prevents the Loss of Dopaminergic Neurons by Inhibiting Microglial Activation in Lipopolysaccharide (LPS)-Induced Parkinson’s Disease Models. Int. J. Mol. Sci. 2017, 18, 2043. [Google Scholar] [CrossRef]
- Sharma, N.; Nehru, B. Characterization of the lipopolysaccharide induced model of Parkinson’s disease: Role of oxidative stress and neuroinflammation. Neurochem. Int. 2015, 87, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.D.; D’Aigle, T.; Gowing, G.; Julien, J.P.; Rivest, S. Exacerbation of motor neuron disease by chronic stimulation of innate immunity in a mouse model of amyotrophic lateral sclerosis. J. Neurosci. 2004, 24, 1340–1349. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Bester, J.; Kell, D.B. A Bacterial Component to Alzheimer’s-Type Dementia Seen via a Systems Biology Approach that Links Iron Dysregulation and Inflammagen Shedding to Disease. J. A. Dis. 2016, 53, 1237–1256. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Stamova, B.; Jin, L.W.; DeCarli, C.; Phinney, B.; Sharp, F.R. Gram-negative bacterial molecules associate with Alzheimer disease pathology. Neurology 2016, 87, 2324–2332. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Lee, Y.K.; Yuk, D.Y.; Choi, D.Y.; Ban, S.B.; Oh, K.W.; Hong, J.T. Neuro-inflammation induced by lipopolysaccharide causes cognitive impairment through enhancement of beta-amyloid generation. J. Neuroinflamm. 2008, 5, 37. [Google Scholar] [CrossRef] [PubMed]
- Lukiw, W.J. Bacteroides fragilis Lipopolysaccharide and Inflammatory Signaling in Alzheimer’s Disease. Front. Microbiol. 2016, 26, 1544. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jaber, V.; Lukiw, W.J. Secretory Products of the Human GI Tract Microbiome and Their Potential Impact on Alzheimer’s Disease (AD): Detection of Lipopolysaccharide (LPS) in AD Hippocampus. Front. Cell Infect. Microbiol. 2017, 7, 318. [Google Scholar] [CrossRef] [PubMed]
- Kallio, K.A.E.; Hätönen, K.A.; Lehto, M.; Salomaa, V.; Männistö, S.; Pussinen, P.J. Endotoxemia, nutrition, and cardiometabolic disorders. Acta Diabetol. 2015, 52, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Gomes, J.M.G.; Costa, J.A.; Alfenas, R.C.G. Metabolic endotoxemia and diabetes mellitus: A systematic review. Metabolism 2017, 68, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Hawkesworth, S.; Moore, S.E.; Fulford, A.J.C.; Barclay, G.R.; Darboe, A.A.; Mark, H.; Nyan, O.A.; Prentice, A.M. Evidence for metabolic endotoxemia in obese and diabetic Gambian women. Nutr. Diabetes 2013, 3, e83. [Google Scholar] [CrossRef]
- Radilla-Vázquez, R.B.; Parra-Rojas, I.; Martínez-Hernández, N.E.; Márquez-Sandoval, Y.F.; Illades-Aguiar, B.; Castro-Alarcón, N. Gut microbiota and metabolic endotoxemia in young obese mexican subjects. Obes. Facts 2016, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Correa, W.; Brandenburg, K.; Zähringer, U.; Ravuri, K.; Khan, T.; von Wintzingerode, F. Biophysical Analysis of Lipopolysaccharide Formulations for an Understanding of the Low Endotoxin Recovery (LER) Phenomenon. Int. J. Mol. Sci. 2017, 18, 2737. [Google Scholar] [CrossRef] [PubMed]
- Niehaus, I. In vivo Radiodetoxification of Salmonella minnesota Lipopolysaccharides with radio-labeled Leucine Enkephalin cures sensory polyneuropathy: A Case report. Niger. Health J. 2010, 10, 26–33. [Google Scholar]
- Gutierrez, K.; Dicks, N.; Glanzner, W.G.; Agellon, L.B.; Bordignon, V. Efficacy of the porcine species in biomedical research. Front. Genet. 2015, 6, 293. [Google Scholar] [CrossRef]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J., Jr.; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Vet. Pathol. 2012, 49, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Caudal, D.; Alvarsson, A.; Björklund, A.; Svenningsson, P. Depressive-like phenotype induced by AAV-mediated overexpression of human α-synuclein in midbrain dopaminergic neurons. Exp. Neurol. 2015, 273, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Tafet, G.E.; Nemeroff, C.B. The Links Between Stress and Depression: Psychoneuroendocrinological, Genetic, and Environmental Interactions. J. Neuropsychiatry Clin. Neurosci. 2016, 28, 77–88. [Google Scholar] [CrossRef]
- Doolin, K.; Farrell, C.; Tozzi, L.; Harkin, A.; Frodl, T.; O’Keane, V. Diurnal hypothalamic-pituitary-adrenal axis measures and inflammatory marker correlates in major depressive disorder. Int. J. Mol. Sci. 2017, 18, 2226. [Google Scholar] [CrossRef] [PubMed]
- Mikołajczyk, A.; Złotkowska, D. Neuroimmunological Implications of Subclinical Lipopolysaccharide from Salmonella Enteritidis. Int. J. Mol. Sci. 2018, 19, 3274. [Google Scholar] [CrossRef]
- Mikołajczyk, A.; Gonkowski, S.; Złotkowska, D. Modulation of the main porcine enteric neuropeptides by a single low-dose of lipopolysaccharide (LPS) Salmonella Enteritidis. Gut Pathog. 2017, 9, 73. [Google Scholar] [CrossRef]
- Mikolajczyk, A.; Makowska, K. Cocaine- And amphetamine-regulated transcript (CART) peptide in the nerve fibres of the porcine gallbladder wall under physiological conditions and after Salmonella Enteritidis lipopolysaccharides administration. Folia Morphol. 2017, 76, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Makowska, K.; Mikolajczyk, A.; Calka, J.; Gonkowski, S. Neurochemical characterization of nerve fibers in the porcine gallbladder wall under physiological conditions and after the administration of Salmonella Enteritidis lipopolysaccharides (LPS). Toxicol. Res. 2018, 7, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Lew, W.Y.W.; Bayna, E.; Molle, E.D.; Dalton, N.D.; Lai, N.C.; Bhargava, V.; Mendiola, V.; Clopton, P.; Tang, T. Recurrent Exposure to Subclinical Lipopolysaccharide Increases Mortality and Induces Cardiac Fibrosis in Mice. PLoS ONE 2013, 8, e61057. [Google Scholar] [CrossRef] [PubMed]
- Calvano, S.E.; Coyle, S.M. Experimental human endotoxemia: A model of the systemic inflammatory response syndrome? Surg. Infect. 2012, 13, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Rittig, N.; Thomsen, H.H.; Bach, E.; Jørgensen, J.O.L.; Møller, N. Hormone and cytokine responses to repeated endotoxin exposures—No evidence of endotoxin tolerance after 5 weeks in humans. Shock 2015, 44, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Freire-Regatillo, A.; Argente-Arizón, P.; Argente, J.; García-Segura, L.M.; Chowen, J.A. Non-neuronal cells in the hypothalamic adaptation to metabolic signals. Front. Endocrinol. 2017, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Xiong, F.; Zhang, L. Role of the hypothalamic-pituitary-adrenal axis in developmental programming of health and disease. Front. Neuroendocrinol. 2013, 34, 27–46. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Taché, Y.F. Activation of brain somatostatin signaling suppresses CRF receptor-mediated stress response. Front. Neurosci. 2017, 11, 231. [Google Scholar] [CrossRef] [PubMed]
- Ait-Ali, D.; Stroth, N.; Sen, J.M.; Eiden, L.E. PACAP-cytokine interactions govern adrenal neuropeptide biosynthesis after systemic administration of LPS. Neuropharmacology 2010, 58, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Incollingo Rodriguez, A.C.; Epel, E.S.; White, M.L.; Standen, E.C.; Seckl, J.R.; Tomiyama, A.J. Hypothalamic-pituitary-adrenal axis dysregulation and cortisol activity in obesity: A systematic review. Psychoneuroendocrinology 2015, 62, 301–318. [Google Scholar] [CrossRef] [PubMed]
- Rutters, F.; La Fleur, S.; Lemmens, S.; Born, J.; Martens, M.; Adam, T. The Hypothalamic-Pituitary-Adrenal Axis, Obesity, and Chronic Stress Exposure: Foods and HPA Axis. Curr. Obes. Rep. 2012, 1, 199–207. [Google Scholar] [CrossRef]
- Sobrino Crespo, C.; PerianesCachero, A.; Puebla Jiménez, L.; Barrios, V.; ArillaFerreiro, E. Peptides and Food Intake. Front. Endocrinol. 2014, 5, 58. [Google Scholar] [CrossRef] [PubMed]
- Boutagy, N.E.; McMillan, R.P.; Frisard, M.I.; Hulver, M.W. Metabolic endotoxemia with obesity: Is it real and is it relevant? Biochimie 2016, 124, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Zhou, J.; Li, D.; Zheng, L.; Xu, Z.; Zhou, S. Long-term over-expression of Neuropeptide Y in hypothalamic paraventricular nucleus contributes to adipose tissue insulin resistance partly via the Y5 receptor. PLoS ONE 2015, 10, e0126714. [Google Scholar] [CrossRef] [PubMed]
- Sergeyev, V.; Broberger, C.; Hökfelt, T. Effect of LPS administration on the expression of POMC, NPY, galanin, CART and MCH mRNAs in the rat hypothalamus. Brain Res. Mol. Brain Res. 2001, 90, 93–100. [Google Scholar] [CrossRef]
- De La Serre, C.B.; de Lartigue, G.; Raybould, H.E. Chronic exposure to low dose bacterial lipopolysaccharide inhibits leptin signaling in vagal afferent neurons. Physiol. Behav. 2015, 139, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Pierpaoli, W. Aging-reversing properties of thyrotropin-releasing hormone. Curr. Aging Sci. 2013, 6, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Boelen, A.; Kwakkel, J.; Thijssen-Timmer, D.C.; Alkemade, A.; Fliers, E.; Wiersinga, W.M. Simultaneous changes in central and peripheral components of the hypothalamus-pituitary-thyroid axis in lipopolysaccharide-induced acute illness in mice. J. Endocrinol. 2004, 182, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Hökfelt, T.; Tatemoto, K. Galanin—25 years with a multitalented neuropeptide. Cell Mol. Life Sci. 2008, 65, 1793–1795. [Google Scholar] [CrossRef] [PubMed]
- Gkonos, P.J.; Tavianini, M.A.; Liu, C.C.; Roos, B.A. Thyrotropin-releasing hormone gene expression in normal thyroid parafollicular cells. Mol. Endocrinol. 1989, 3, 2101–2109. [Google Scholar] [CrossRef] [PubMed]
- Iversen, E.; Laurberg, P. Thyrotrophin-releasing hormone (TRH) and hormone secretion from the follicular and C-cells of perfused dog thyroid lobes. Acta Endocrinol. 1985, 109, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Ahren, B. Regulatory peptides in the thyroid gland—A review on their localization and function. Acta Endocrinol. 1991, 124, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Morillo-Bernal, J.; Fernández-Santos, J.M.; Utrilla, J.C.; de Miguel, M.; García-Marín, R.; Martín-Lacave, I. Functional expression of the thyrotropin receptor in C cells: New insights into their involvement in the hypothalamic-pituitary-thyroid axis. J. Anat. 2009, 215, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Terasawa, E.; Fernandez, D.L. Neurobiological mechanisms of the onset of puberty in primates. Endocr. Rev. 2001, 22, 111–151. [Google Scholar] [CrossRef] [PubMed]
- Manfredi-Lozano, M.; Roa, J.; Tena-Sempere, M. Connecting metabolism and gonadal function: Novel central neuropeptide pathways involved in the metabolic control of puberty and fertility. Front. Neuroendocrinol. 2017, 48, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Constantin, S.; Wray, S. Galanin Activates G Protein Gated Inwardly Rectifying Potassium Channels and Suppresses Kisspeptin-10 Activation of GnRH Neurons. Endocrinology 2016, 157, 3197–3212. [Google Scholar] [CrossRef] [PubMed]
- Splett, C.L.; Scheffen, J.R.; Desotelle, J.A.; Plamann, V.; Bauer-Dantoin, A.C. Galanin enhancement of gonadotropin-releasing hormone-stimulated luteinizing hormone secretion in female rats is estrogen dependent. Endocrinology 2003, 144, 484–490. [Google Scholar] [CrossRef]
- Nuruddin, S.; Syverstad, G.H.E.; Lillehaug, S.; Leergaard, T.B.; Nilsson, L.N.G.; Ropstad, E.; Krogenæs, A.; Haraldsen, I.R.H.; Torp, R. Elevated mRNA-Levels of gonadotropin-releasing hormone and its receptor in plaque-bearing Alzheimer’s Disease transgenic mice. PLoS ONE 2014, 9, e103607. [Google Scholar] [CrossRef]
- Ishii, M.; Iadecola, C. Metabolic and Non-Cognitive Manifestations of Alzheimer’s Disease: The Hypothalamus as Both Culprit and Target of Pathology. Cell Metab. 2015, 22, 761–776. [Google Scholar] [CrossRef]
- Thornton, E.; Vink, R. Substance P and its tachykinin NK1 receptor: A novel neuroprotective target for Parkinson’s disease. Neural Regen. Res. 2015, 10, 1403–1405. [Google Scholar]
- Wang, C.Z.; Kazmierczak, R.A.; Eisenstark, A. Strains, mechanism, and perspective: Salmonella-based cancer therapy. Int. J. Microbiol. 2016, 2016, 5678702. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Qu, C.; Lu, X.; Zhang, S. Activation of microglia by histamine and substance P. Cell Physiol. Biochem. 2014, 34, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.S.; Herrera-Marschitz, M.; Ungerstedt, U. Effects of intranigral substance P and neurokinin A injections on extracellular dopamine levels measured with microdialysis in the striatum and frontoparietal cortex of rats. J. Neurochem. 1991, 57, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Morara, S.; Colangelo, A.M.; Provini, L. Microglia-Induced Maladaptive Plasticity Can Be Modulated by Neuropeptides In Vivo. Neural Plast. 2015, 2015, 135342. [Google Scholar] [CrossRef] [PubMed]
- Carniglia, L.; Ramírez, D.; Durand, D.; Saba, J.; Turati, J.; Caruso, C.; Scimonelli, T.N.; Lasaga, M. Neuropeptides and Microglial Activation in Inflammation, Pain, and Neurodegenerative Diseases. Mediat. Inflamm. 2017, 2017, 5048616. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.; Santos, T.; Cortes, L.; Cochaud, S.; Agasse, F.; Silva, A.P.; Xapelli, S.; Malva, J.O. Neuropeptide y inhibits interleukin-1 beta-induced microglia motility. J. Neurochem. 2012, 120, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.J.; Dong, C.Z.; Li, W.L.; Bu, W.; Wu, J.; Zhao, W.Q. Neuropeptide Y protects cerebral cortical neurons by regulating microglial immune function. Neural Regen. Res. 2014, 9, 959–967. [Google Scholar] [PubMed]
- Counts, S.E.; Perez, S.E.; Ginsberg, S.D.; Mufson, E.J. Neuroprotective role for galanin in Alzheimer’s disease. EXS 2010, 102, 143–162. [Google Scholar]
- Alexandris, A.; Liu, A.K.L.; Chang, R.C.C.; Pearce, R.K.B.; Gentleman, S.M. Differential expression of galanin in the cholinergic basal forebrain of patients with Lewy body disorders. Acta Neuropathol. Commun. 2015, 3, 77. [Google Scholar] [CrossRef]
- Weinshenker, D.; Holmes, P.V. Regulation of neurological and neuropsychiatric phenotypes by locus coeruleus-derived galanin. Brain Res. 2016, 1641 Pt B, 320–337. [Google Scholar] [CrossRef]
- Lundström, L.; Elmquist, A.; Bartfai, T.; Langel, Ü. Galanin and its receptors in neurological disorders. Neuromol. Med. 2005, 7, 157–180. [Google Scholar] [CrossRef]
- Lang, R.; Gundlach, A.L.; Holmes, F.E.; Hobson, S.A.; Wynick, D.; Hokfelt, T.; Kofler, B. Physiology, Signaling, and Pharmacology of Galanin Peptides and Receptors: Three Decades of Emerging Diversity. Pharmacol. Rev. 2014, 67, 118–175. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.; Leceta, J.; Ganea, D. Vasoactive intestinal peptide and pituitary adenylate cyclase- activating polypeptide inhibit the production of inflammatory mediators by activated microglia. J. Leukoc. Biol. 2003, 73, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Xiong, J.-X.; Wang, Y.-Y.; Tang, J.; Zhang, B.; Bai, Y. VIP enhances phagocytosis of fibrillar beta-amyloid by microglia and attenuates amyloid deposition in the brain of APP/PS1 mice. PLoS ONE 2012, 7, e29790. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Zhang, X.; Li, X.; Liu, N.; Lou, F.; Ma, H.; Luo, X.; Ren, Y. Somatostatin prevents lipopolysaccharide-induced neurodegeneration in the rat substantia nigra by inhibiting the activation of microglia. Mol. Med. Rep. 2015, 12, 1002–1008. [Google Scholar] [CrossRef] [PubMed]
- Reubi, J.C.; Schonbrunn, A. Illuminating somatostatin analog action at neuroendocrine tumor receptors. Trends Pharmacol. Sci. 2013, 34, 676–688. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Taratula, O.; Taratula, O.; Schumann, C.; Minko, T. LHRH-Targeted Drug Delivery Systems for Cancer Therapy. Mini Rev. Med. Chem. 2017, 17, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Rojas, A.; Pérez-Solis, M.A.; Maya-Núñez, G. The gonadotropin-releasing hormone system: Perspectives from reproduction to cancer (Review). Int. J. Oncol. 2016, 48, 861–868. [Google Scholar] [CrossRef]
- Szponar, B.; Kraśnik, L.; Hryniewiecki, T.; Gamian, A.; Larsson, L. Distribution of 3-hydroxy fatty acids in tissues after intrapentoneal injection of endotoxin. Clin. Chem. 2003, 49, 1149–1153. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Ding, X. Methods of Endotoxin Detection. J. Lab. Autom. 2015, 20, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Nakasone, M.; Nakaso, K.; Horikoshi, Y.; Hanaki, T.; Kitagawa, Y.; Takahashi, T.; Inagaki, Y.; Matsura, T. Preconditioning by low dose LPS prevents subsequent LPS-induced severe liver injury via Nrf2 activation in mice. Yonago Acta Med. 2016, 59, 223–231. [Google Scholar] [PubMed]
- Hammerich, L.; Binder, A.; Brody, J.D. In situ vaccination: Cancer immunotherapy both personalized and off-the-shelf. Mol. Oncol. 2015, 9, 1966–1981. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, T.; Nakagawa, M.; Kiriyama, Y.; Toyoda, T.; Cao, X. Prevention of gastric cancer: Eradication of Helicobacter pylori and beyond. Int. J. Mol. Sci. 2017, 18, 1699. [Google Scholar] [CrossRef] [PubMed]
- Mikołajczyk, A. Safe and effective anaesthesiological protocols in domestic pig. Ann. Warsaw Univ. Life Sci. SGGW Anim. Sci. 2016, 55, 219–227. [Google Scholar]
- Fu, H.Q.; Yang, T.; Xiao, W.; Fan, L.; Wu, Y.; Terrando, N.; Wang, T.L. Prolonged neuroinflammation after lipopolysaccharide exposure in aged rats. PLoS ONE 2014, 9, e106331. [Google Scholar] [CrossRef] [PubMed]
- Lopes, P.C. LPS and neuroinflammation: A matter of timing. Inflammopharmacology 2016, 24, 291–293. [Google Scholar] [CrossRef] [PubMed]
- Cox, N.M.; Britt, J.H. Effect of estradiol on hypothalamic GnRH and pituitary and serum LH and FSH in ovariectomized pigs. J. Anim. Sci. 1982, 55, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Ettrup, K.S.; Sørensen, J.C.; Bjarkam, C.R. The anatomy of the Göttingen minipig hypothalamus. J. Chem. Neuroanat. 2010, 39, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Sesti, L.A.; Britt, J.H. Relationship of secretion of GnRH in vitro to changes in pituitary concentrations of LH and FSH and serum concentrations of LH during lactation in sows. J. Reprod. Fertil. 1993, 98, 393–400. [Google Scholar] [CrossRef]
- Félix, B.; Léger, M.E.; Albe-Fessard, D.; Marcilloux, J.C.; Rampin, O.; Laplace, J.P. Stereotaxic atlas of the pig brain. Brain Res. Bull. 1999, 49, 1–137. [Google Scholar] [CrossRef]
- Schmidt, V. Comparative Anatomy of the Pig Brain—An Integrative Magnetic Resonance Imaging (MRI) Study of the Porcine Brain with Special Emphasis on the External Morphology of the Cerebral Cortex. Ph.D. Thesis, Giessen University, Vvb Laufersweiler Verlag, Giessen, Germasny, 2014; pp. 1–168. [Google Scholar]
- Conlon, J.M. Purification of naturally occurring peptides by reversed-phase HPLC. Nat. Protoc. 2007, 2, 191–197. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikołajczyk, A.; Złotkowska, D. Subclinical Lipopolysaccharide from Salmonella Enteritidis Induces Dysregulation of Bioactive Substances from Selected Brain Sections and Glands of Neuroendocrine Axes. Toxins 2019, 11, 91. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11020091
Mikołajczyk A, Złotkowska D. Subclinical Lipopolysaccharide from Salmonella Enteritidis Induces Dysregulation of Bioactive Substances from Selected Brain Sections and Glands of Neuroendocrine Axes. Toxins. 2019; 11(2):91. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11020091
Chicago/Turabian StyleMikołajczyk, Anita, and Dagmara Złotkowska. 2019. "Subclinical Lipopolysaccharide from Salmonella Enteritidis Induces Dysregulation of Bioactive Substances from Selected Brain Sections and Glands of Neuroendocrine Axes" Toxins 11, no. 2: 91. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11020091