Forty Years of the Description of Brown Spider Venom Phospholipases-D

 , and
, and
Abstract
:1. Molecular Characteristics of Brown Spider Venoms
2. Nomenclatures and Biochemical Classification of Brown Spider PLDs
3. The History of PLDs in Loxoscelism and the Production of These Toxins as Recombinant Proteins
4. Studies with Loxosceles Venom PLDs Proved Their Participation in the Physiopathology of Envenoming
5. Structural Organization and the Catalytic Mechanisms of Brown Spider PLDs
6. Proteomic Analysis in the Learning about Brown Spider Venom PLDs
7. Cellular Biology of the Brown Spider Venoms PLDs
8. A New Generation of Anti-loxoscelic Sera and Vaccines using Loxosceles Recombinant PLDs and Synthetic Peptides
9. Brown Spider PLDs as Models for the Discovery of Specific Treatments for Loxoscelism and as Tools in New Anti-inflammatory Drugs Description
10. Diagnosis of Loxoscelism Based on the Identification of PLDs or Related Products
11. Biotechnological Applications of Loxosceles PLDs
12. Actions Expected for the Future of Loxosceles PLD Researches
Author Contributions
Funding
Conflicts of Interest
References
- Da Silva, P.H.; Hashimoto, Y.; Dos Santos, F.A.; Mangili, O.C.; Gremski, W.; Veiga, S.S. Hematological cell findings in bone marrow and peripheral blood of rabbits after experimental acute exposure to Loxosceles intermedia (brown spider) venom. Toxicon 2003, 42, 155–161. [Google Scholar] [CrossRef]
- Da Silva, P.H.; Da Silveira, R.B.; Helena Appel, M.; Mangili, O.C.; Gremski, W.; Veiga, S.S. Brown spiders and loxoscelism. Toxicon 2004, 44, 693–709. [Google Scholar] [CrossRef] [PubMed]
- Hogan, C.J.; Barbaro, K.C.; Winkel, K. Loxoscelism: Old obstacles, new directions. Ann. Emerg. Med. 2004, 44, 608–624. [Google Scholar] [CrossRef] [PubMed]
- Barceloux, D.G. Medical Toxicology of Natural Substances: Foods, Fungi, Medicinal Herbs, Plants, and Venomous Animals; John Wiley & Sons: Hoboken, NJ, USA, 2008; ISBN 9780471727613. [Google Scholar]
- Gremski, L.H.; Trevisan-Silva, D.; Ferrer, V.P.; Matsubara, F.H.; Meissner, G.O.; Wille, A.C.M.; Vuitika, L.; Dias-Lopes, C.; Ullah, A.; De Moraes, F.R.; et al. Recent advances in the understanding of brown spider venoms: From the biology of spiders to the molecular mechanisms of toxins. Toxicon 2014, 83, 91–120. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Moreira, D.; Senff-Ribeiro, A.; Wille, A.C.M.; Gremski, L.H.; Chaim, O.M.; Veiga, S.S. Highlights in the knowledge of brown spider toxins. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sams, H.H.; Dunnick, C.A.; Smith, M.L.; King, L.E. Necrotic arachnidism. J. Am. Acad. Dermatol. 2001, 44, 561–576. [Google Scholar] [CrossRef]
- Gremski, L.H.; Da Silveira, R.B.; Chaim, O.M.; Probst, C.M.A.; Ferrer, V.P.; Nowatzki, J.; Weinschutz, H.C.; Madeira, H.M.I.; Gremski, W.; Nader, H.B.; et al. A novel expression profile of the Loxosceles intermedia spider venomous gland revealed by transcriptome analysis. Mol. Biosyst. 2010, 6, 2403–2416. [Google Scholar] [CrossRef]
- Fernandes-Pedrosa, M.; de, F.; Junqueira-de-Azevedo, I.D.L.M.; Gonçalves-de-Andrade, R.M.; Kobashi, L.S.; Almeida, D.D.; Ho, P.L.; Tambourgi, D.V. Transcriptome analysis of Loxosceles laeta (Araneae, Sicariidae) spider venomous gland using expressed sequence tags. BMC Genomics 2008, 9, 279. [Google Scholar] [CrossRef] [Green Version]
- Dantas, A.E.; Carmo, A.O.; Horta, C.C.R.; Leal, H.G.; Oliveira-Mendes, B.B.R.; Martins, A.P.V.; Chávez-Olórtegui, C.; Kalapothakis, E. Description of Loxtox protein family and identification of a new group of Phospholipases D from Loxosceles similis venom gland. Toxicon 2016, 120, 97–106. [Google Scholar] [CrossRef]
- Segura-Ramírez, P.J.; Silva Júnior, P.I. Loxosceles gaucho spider venom: An untapped source of antimicrobial agents. Toxins 2018, 10, 522. [Google Scholar] [CrossRef] [Green Version]
- Zobel-Thropp, P.A.; Kerins, A.E.; Binford, G.J. Sphingomyelinase D in sicariid spider venom is a potent insecticidal toxin. Toxicon 2012, 60, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira, R.B.; Pigozzo, R.B.; Chaim, O.M.; Appel, M.H.; Dreyfuss, J.L.; Toma, L.; Mangili, O.C.; Gremski, W.; Dietrich, C.P.; Nader, H.B.; et al. Molecular cloning and functional characterization of two isoforms of dermonecrotic toxin from Loxosceles intermedia (Brown spider) venom gland. Biochimie 2006, 88, 1241–1253. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira, R.B.; Pigozzo, R.B.; Chaim, O.M.; Appel, M.H.; Silva, D.T.; Dreyfuss, J.L.; Toma, L.; Dietrich, C.P.; Nader, H.B.; Veiga, S.S.; et al. Two novel dermonecrotic toxins LiRecDT4 and LiRecDT5 from Brown spider (Loxosceles intermedia) venom: From cloning to functional characterization. Biochimie 2007, 89, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.O.S.; Chaim, O.M.; da Silveira, R.B.; Gremski, L.H.; Sade, Y.B.; Paludo, K.S.; Senff-Ribeiro, A.; de Moura, J.; Chávez-Olórtegui, C.; Gremski, W.; et al. Biological and structural comparison of recombinant phospholipase D toxins from Loxosceles intermedia (brown spider) venom. Toxicon 2007, 50, 1162–1174. [Google Scholar] [CrossRef] [PubMed]
- Vuitika, L.; Gremski, L.H.; Belisário-Ferrari, M.R.; Chaves-Moreira, D.; Ferrer, V.P.; Senff-Ribeiro, A.; Chaim, O.M.; Veiga, S.S. Brown spider phospholipase-D containing a conservative mutation (D233E) in the catalytic site: Identification and functional characterization. J. Cell. Biochem. 2013, 114, 2479–2492. [Google Scholar] [CrossRef] [PubMed]
- Ospedal, K.Z.; Appel, M.H.; Neto, J.F.; Mangili, O.C.; Veiga, S.S.; Gremski, W. Histopathological findings in rabbits after experimental acute exposure to the Loxosceles intermedia (brown spider) venom. Int. J. Exp. Pathol. 2002, 83, 287–294. [Google Scholar] [CrossRef]
- De Santi Ferrara, G.I.; Fernandes-Pedrosa, M.; de, F.; Junqueira-de-Azevedo, I.; de, L.M.; Gonçalves-de-Andrade, R.M.; Portaro, F.C.V.; Manzoni-de-Almeida, D.; Murakami, M.T.; Arni, R.K.; et al. SMase II, a new sphingomyelinase D from Loxosceles laeta venom gland: Molecular cloning, expression, function and structural analysis. Toxicon 2009, 53, 743–753. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Futrell, J.M. Loxoscelism. Am. J. Med. Sci. 1992, 304, 261–267. [Google Scholar] [CrossRef]
- Atkins, J.A.; Wingo, C.W.; Sodeman, W.A. Probable cause of necrotic spider bite in the Midwest. Science 1957, 126, 73. [Google Scholar] [CrossRef]
- Atkins, J.A.; Wingo, C.W.; Sodeman, W.A.; Flynn, J.E. Necrotic arachnidism. Am. J. Trop. Med. Hyg. 1958, 7, 165–184. [Google Scholar] [CrossRef] [PubMed]
- Forrester, L.J.; Barrett, J.T.; Campbell, B.J. Red blood cell lysis induced by the venom of the brown recluse spider. The role of sphingomyelinase D. Arch. Biochem. Biophys. 1978, 187, 355–365. [Google Scholar] [CrossRef]
- Kurpiewski, G.; Forrester, L.J.; Barrett, J.T.; Campbell, B.J. Platelet aggregation and sphingomyelinase D activity of a purified toxin from the venom of loxosceles reclusa. BBA-Gen. Subj. 1981, 678, 467–476. [Google Scholar] [CrossRef]
- Gates, C.A.; Rees, R.S. Serum amyloid P component: Its role in platelet activation stimulated by sphingomyelinase D purified from the venom of the brown recluse spider (Loxosceles reclusa). Toxicon 1990, 28, 1303–1315. [Google Scholar] [CrossRef]
- Machado, L.F.; Laugesen, S.; Botelho, E.D.; Ricart, C.A.O.; Fontes, W.; Barbaro, K.C.; Roepstorff, P.; Valle De Sousa, M. Proteome analysis of brown spider venom: Identification of loxnecrogin isoforms in Loxosceles gaucho venom. Proteomics 2005, 5, 2167–2176. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lynch, K.R. Brown recluse spider (Loxosceles reclusa) venom phospholipase D (PLD) generates lysophosphatidic acid (LPA). Biochem. J. 2006, 391, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Chaim, O.M.; Da Silveira, R.B.; Trevisan-Silva, D.; Ferrer, V.P.; Sade, Y.B.; Bóia-Ferreira, M.; Gremski, L.H.; Gremski, W.; Senff-Ribeiro, A.; Takahashi, H.K.; et al. Phospholipase-D activity and inflammatory response induced by brown spider dermonecrotic toxin: Endothelial cell membrane phospholipids as targets for toxicity. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2011, 1811, 84–96. [Google Scholar] [CrossRef] [Green Version]
- Chaves-Moreira, D.; Souza, F.N.; Fogaça, R.T.H.; Mangili, O.C.; Gremski, W.; Senff-Ribeiro, A.; Chaim, O.M.; Veiga, S.S. The relationship between calcium and the metabolism of plasma membrane phospholipids in hemolysis induced by brown spider venom phospholipase-D toxin. J. Cell. Biochem. 2011, 112, 2529–2540. [Google Scholar] [CrossRef]
- Lajoie, D.M.; Roberts, S.A.; Zobel-Thropp, P.A.; Delahaye, J.L.; Bandarian, V.; Binford, G.J.; Cordes, M.H.J. Variable substrate preference among phospholipase D toxins from sicariid spiders. J. Biol. Chem. 2015, 290, 10994–11007. [Google Scholar] [CrossRef] [Green Version]
- Kalapothakis, E.; Chatzaki, M.; Gonçalves-Dornelas, H.; de Castro, C.S.; Silvestre, F.G.; Laborne, F.V.; de Moura, J.F.; Veiga, S.S.; Chávez-Olórtegui, C.; Granier, C.; et al. The Loxtox protein family in Loxosceles intermedia (Mello-Leitão) venom. Toxicon 2007, 50, 938–946. [Google Scholar] [CrossRef]
- Binford, G.J.; Cordes, M.H.J.; Wells, M.A. Sphingomyelinase D from venoms of Loxosceles spiders: Evolutionary insights from cDNA sequences and gene structure. Toxicon 2005, 45, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Binford, G.J.; Callahan, M.S.; Bodner, M.R.; Rynerson, M.R.; Núñez, P.B.; Ellison, C.E.; Duncan, R.P. Phylogenetic relationships of Loxosceles and Sicarius spiders are consistent with Western Gondwanan vicariance. Mol. Phylogenet. Evol. 2008, 49, 538–553. [Google Scholar] [CrossRef] [PubMed]
- Binford, G.J.; Bodner, M.R.; Cordes, M.H.J.; Baldwin, K.L.; Rynerson, M.R.; Burns, S.N.; Zobel-Thropp, P.A. Molecular evolution, functional variation, and proposed nomenclature of the gene family that includes sphingomyelinase D in sicariid spider venoms. Mol. Biol. Evol. 2009, 26, 547–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denny, W.F.; Dillaha, C.J.; Morgan, P.N. Hemotoxic effect of Loxosceles reclusus venom: In vivo and in vitro studies. J. Lab. Clin. Med. 1964, 64, 291–298. [Google Scholar] [PubMed]
- Morgan, P.N.; Felton, W.W. Utilization of mammalian cell cultures in spider venom studies. Bact. Proc. 1965, 133, 276. [Google Scholar]
- Smith, C.W.; Micks, D.W. The role of polymorphonuclear leukocytes in the lesion caused by the venom of the brown spider, Loxosceles reclusa. Lab. Investig. 1970, 22, 90–93. [Google Scholar] [PubMed]
- Majeski, J.A.; Stinnett, J.D.; Alexander, J.W.; Durst, G.G. Action of venom from the brown recluse spider (Loxosceles reclusa) on human neutrophils. Toxicon 1977, 15, 423–427. [Google Scholar] [CrossRef]
- Kniker, W.T.; Morgan, P.N.; Flanigan, W.J.; Reagan, P.W.; Dillaha, C.J. An inhibitor of complement in the venom of the brown recluse spider, Loxosceles reclusa. Proc. Soc. Exp. Biol. Med. 1969, 131, 1432–1434. [Google Scholar] [CrossRef]
- Patel, K.D.; Modur, V.; Zimmerman, G.A.; Prescott, S.M.; McIntyre, T.M. The necrotic venom of the brown recluse spider induces dysregulated endothelial cell-dependent neutrophil activation. Differential induction of GM-CSF, IL-8, and E-selectin expression. J. Clin. Investig. 1994, 94, 631–642. [Google Scholar] [CrossRef]
- Barbaro, K.C.; Cardoso, J.L.C.; Eickstedt, V.R.D.; Mota, I. Dermonecrotic and lethal components of Loxosceles gaucho spider venom. Toxicon 1992, 30, 331–338. [Google Scholar] [CrossRef]
- Tambourgi, D.V.; Magnoli, F.C.; von Eickstedt, V.R.D.; Benedetti, Z.C.; Petricevich, V.L.; Da Silva, W.D. Incorporation of a 35-Kilodalton Purified Protein from Loxosceles intermedia Spider Venom Transforms Human Erythrocytes into Activators of Autologous Complement Alternative Pathway. J. Immunol. 1995, 155, 4459–4466. [Google Scholar] [PubMed]
- Barbaro, K.C.; Ferreira, M.L.; Cardoso, D.F.; Eickstedt, V.R.D.; Mota, I. Identification and neutralization of biological activities in the venoms of Loxosceles spiders. Braz. J. Med. Biol. Res. 1996, 29, 1491–1497. [Google Scholar] [PubMed]
- Tambourgi, D.V.; Magnoli, F.C.; Van Den Berg, C.W.; Morgan, B.P.; De Araujo, P.S.; Alves, E.W.; Da Silva, W.D. Sphingomyelinases in the venom of the spider Loxosceles intermedia are responsible for both dermonecrosis and complement-dependent hemolysis. Biochem. Biophys. Res. Commun. 1998, 251, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Kalapothakis, E.; Araujo, S.C.; De Castro, C.S.; Mendes, T.M.; Gomez, M.V.; Mangili, O.C.; Gubert, I.C.; Chávez-Olórtegui, C. Molecular cloning, expression and immunological properties of LiD1, a protein from the dermonecrotic family of Loxosceles intermedia spider venom. Toxicon 2002, 40, 1691–1699. [Google Scholar] [CrossRef]
- Fernandes Pedrosa, M.D.F.; Junqueira de Azevedo, I.D.L.M.; Gonçalves-de-Andrade, R.M.; Van Den Berg, C.W.; Ramos, C.R.R.; Lee Ho, P.; Tambourgi, D.V. Molecular cloning and expression of a functional dermonecrotic and haemolytic factor from Loxosceles laeta venom. Biochem. Biophys. Res. Commun. 2002, 298, 638–645. [Google Scholar] [CrossRef]
- Chaim, O.M.; Sade, Y.B.; Da Silveira, R.B.; Toma, L.; Kalapothakis, E.; Chávez-Olórtegui, C.; Mangili, O.C.; Gremski, W.; Von Dietrich, C.P.; Nader, H.B.; et al. Brown spider dermonecrotic toxin directly induces nephrotoxicity. Toxicol. Appl. Pharmacol. 2006, 211, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Appel, M.H.; da Silveira, R.B.; Chaim, O.M.; Paludo, K.S.; Silva, D.T.; Chaves, D.M.; da Silva, P.H.; Mangili, O.C.; Senff-Ribeiro, A.; Gremski, W.; et al. Identification, cloning and functional characterization of a novel dermonecrotic toxin (phospholipase D) from brown spider (Loxosceles intermedia) venom. Biochim. Biophys. Acta Gen. Subj. 2008, 1780, 167–178. [Google Scholar] [CrossRef]
- Wille, A.C.M.; Chaves-Moreira, D.; Trevisan-Silva, D.; Magnoni, M.G.; Boia-Ferreira, M.; Gremski, L.H.; Gremski, W.; Chaim, O.M.; Senff-Ribeiro, A.; Veiga, S.S. Modulation of membrane phospholipids, the cytosolic calcium influx and cell proliferation following treatment of B16-F10 cells with recombinant phospholipase-D from Loxosceles intermedia (brown spider) venom. Toxicon 2013, 67, 17–30. [Google Scholar] [CrossRef]
- Dias-Lopes, C.; Felicori, L.; Rubrecht, L.; Cobo, S.; Molina, L.; Nguyen, C.; Galéa, P.; Granier, C.; Molina, F.; Chávez-Olortegui, C. Generation and molecular characterization of a monoclonal antibody reactive with conserved epitope in sphingomyelinases D from Loxosceles spider venoms. Vaccine 2014, 32, 2086–2092. [Google Scholar] [CrossRef] [Green Version]
- Olvera, A.; Ramos-Cerrillo, B.; Estévez, J.; Clement, H.; de Roodt, A.; Paniagua-Solís, J.; Vázquez, H.; Zavaleta, A.; Salas Arruz, M.; Stock, R.P.; et al. North and South American Loxosceles spiders: Development of a polyvalent antivenom with recombinant sphingomyelinases D as antigens. Toxicon 2006, 48, 64–74. [Google Scholar] [CrossRef]
- Catalán, A.; Cortes, W.; Sagua, H.; González, J.; Araya, J.E. Two new phospholipase D isoforms of Loxosceles laeta: Cloning, heterologous expression, functional characterization, and potential biotechnological application. J. Biochem. Mol. Toxicol. 2011, 25, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, G.S.; Caporrino, M.C.; Della-Casa, M.S.; Kimura, L.F.; Prezotto-Neto, J.P.; Fukuda, D.A.; Portes-Junior, J.A.; Neves-Ferreira, A.G.C.; Santoro, M.L.; Barbaro, K.C. Cloning, expression and characterization of a phospholipase D from Loxosceles gaucho venom gland. Biochimie 2013, 95, 1773–1783. [Google Scholar] [CrossRef] [PubMed]
- Tambourgi, D.V.; Pedrosa, M.D.F.F.; Van Den Berg, C.W.; Gonçalves-De-Andrade, R.M.; Ferracini, M.; Paixão-Cavalcante, D.; Morgan, B.P.; Rushmere, N.K. Molecular cloning, expression, function and immunoreactivities of members of a gene family of sphingomyelinases from Loxosceles venom glands. Mol. Immunol. 2004, 41, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Felicori, L.; Araujo, S.C.; Machado de Ávila, R.A.; Sanchez, E.F.; Granier, C.; Kalapothakis, E.; Chávez-Olórtegui, C. Functional characterization and epitope analysis of a recombinant dermonecrotic protein from Loxosceles intermedia spider. Toxicon 2006, 48, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Dias-Lopes, C.; Guimarães, G.; Felicori, L.; Fernandes, P.; Emery, L.; Kalapothakis, E.; Nguyen, C.; Molina, F.; Granier, C.; Chávez-Olórtegui, C. A protective immune response against lethal, dermonecrotic and hemorrhagic effects of Loxosceles intermedia venom elicited by a 27-residue peptide. Toxicon 2010, 55, 481–487. [Google Scholar] [CrossRef]
- Dias-Lopes, C.; Felicori, L.; Guimarães, G.; Gomes, E.R.M.; Roman-Campos, D.; Duarte, H.; Damasceno, D.; Martins, M.; Kalapothakis, E.; Almeida, A.P.; et al. Cardiotoxic effects of Loxosceles intermedia spider venom and the recombinant venom toxin rLiD1. Toxicon 2010, 56, 1426–1435. [Google Scholar] [CrossRef]
- Rojas, J.M.; Arán-Sekul, T.; Cortés, E.; Jaldín, R.; Ordenes, K.; Orrego, P.R.; González, J.; Araya, J.E.; Catalán, A. Phospholipase d from loxosceles laeta spider venom induces IL-6, IL-8, CXCL1/GRO-α, and CCL2/MCP-1 production in human skin fibroblasts and stimulates monocytes migration. Toxins 2017, 9, 125. [Google Scholar] [CrossRef]
- Kusma, J.; Chaim, O.M.; Wille, A.C.M.; Ferrer, V.P.; Sade, Y.B.; Donatti, L.; Gremski, W.; Mangili, O.C.; Veiga, S.S. Nephrotoxicity caused by brown spider venom phospholipase-D (dermonecrotic toxin) depends on catalytic activity. Biochimie 2008, 90, 1722–1736. [Google Scholar] [CrossRef]
- Oliveira, C.F.B.; Vilela, A.; Coura, L.A.M.; Rodrigues, F.T.G.; Nagem, R.A.P.; Chávez-Olortegui, C.; Maioli, T.U.; Felicori, L.F. Protective antibodies against a sphingomyelinase D from Loxosceles intermedia spider venom elicited in mice with different genetic background. Vaccine 2016, 34, 3828–3834. [Google Scholar] [CrossRef]
- Vuitika, L.; Chaves-Moreira, D.; Caruso, I.; Lima, M.A.; Matsubara, F.H.; Murakami, M.T.; Takahashi, H.K.; Toledo, M.S.; Coronado, M.A.; Nader, H.B.; et al. Active site mapping of Loxosceles phospholipases D: Biochemical and biological features. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 970–979. [Google Scholar] [CrossRef]
- Murakami, M.T.; Fernandes-Pedrosa, M.F.; Tambourgi, D.V.; Arni, R.K. Structural basis for metal ion coordination and the catalytic mechanism of sphingomyelinases D. J. Biol. Chem. 2005, 280, 13658–13664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Giuseppe, P.O.; Ullah, A.; Silva, D.T.; Gremski, L.H.; Wille, A.C.M.; Chaves Moreira, D.; Ribeiro, A.S.; Chaim, O.M.; Murakami, M.T.; Veiga, S.S.; et al. Structure of a novel class II phospholipase D: Catalytic cleft is modified by a disulphide bridge. Biochem. Biophys. Res. Commun. 2011, 409, 622–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, A.; Magalhães, G.S.; Masood, R.; Mariutti, R.B.; Coronado, M.A.; Murakami, M.T.; Barbaro, K.C.; Arni, R.K. Crystallization and preliminary X-ray diffraction analysis of a novel sphingomyelinase D from Loxosceles gaucho venom. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2014, 70, 1418–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masood, R.; Ullah, K.; Ali, H.; Ali, I.; Betzel, C.; Ullah, A. Spider’s venom phospholipases D: A structural review. Int. J. Biol. Macromol. 2018, 107, 1054–1065. [Google Scholar] [CrossRef]
- Andrade, S.A.D.; Murakami, M.T.; Cavalcante, D.P.; Arni, R.K.; Tambourgi, D.V. Kinetic and mechanistic characterization of the Sphingomyelinases D from Loxosceles intermedia spider venom. Toxicon 2006, 47, 380–386. [Google Scholar] [CrossRef]
- Lajoie, D.M.; Zobel-Thropp, P.A.; Kumirov, V.K.; Bandarian, V.; Binford, G.J.; Cordes, M.H.J. Phospholipase D Toxins of Brown Spider Venom Convert Lysophosphatidylcholine and Sphingomyelin to Cyclic Phosphates. PLoS ONE 2013, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Catalán, A.; Cortés, W.; Muñoz, C.; Araya, J.E. Tryptophan and aspartic acid residues present in the glycerophosphoryl diester phosphodiesterase (GDPD) domain of the Loxosceles laeta phospholipase D are essential for substrate recognition. Toxicon 2014, 81, 43–47. [Google Scholar] [CrossRef]
- Cheng, J.; Goldstein, R.; Gershenson, A.; Stec, B.; Roberts, M.F. The cation-π box is a specific phosphatidylcholine membrane targeting motif. J. Biol. Chem. 2013, 288, 14863–14873. [Google Scholar] [CrossRef] [Green Version]
- Gorelik, A.; Liu, F.; Illes, K.; Nagar, B. Crystal structure of the human alkaline sphingomyelinase provides insights into substrate recognition. J. Biol. Chem. 2017, 292, 7087–7094. [Google Scholar] [CrossRef] [Green Version]
- Cunha, R.B.; Barbaro, K.C.; Muramatsu, D.; Vieira Portaro, F.C.; Fontes, W.; De Sousa, M.V. Purification and Characterization of Loxnecrogin, a Dermonecrotic Toxin from Loxosceles gaucho Brown Spider Venom. J. Protein Chem. 2003, 22, 135–146. [Google Scholar] [CrossRef]
- Binford, G.J.; Wells, M.A. The phylogenetic distribution of sphingomyelinase D activity in venoms of Haplogyne spiders. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 135, 25–33. [Google Scholar] [CrossRef]
- Luciano, M.N.; Da Silva, P.H.; Chaim, O.M.; Dos Santos, V.L.P.; Franco, C.R.C.; Soares, M.F.S.; Zanata, S.M.; Mangili, O.C.; Gremski, W.; Veiga, S.S. Experimental Evidence for a Direct Cytotoxicity of Loxosceles intermedia (Brown Spider) Venom in Renal Tissue. J. Histochem. Cytochem. 2004, 52, 455–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pretel, F.; Gonçalves-De-Andrade, R.M.; Magnoli, F.C.; Da Silva, M.E.R.; Ferreira, J.M.C.; Van Den Berg, C.W.; Tambourgi, D.V. Analysis of the toxic potential of venom from Loxosceles adelaida, a Brazilian brown spider from karstic areas. Toxicon 2005, 45, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, F.G.; Castro, C.S.D.; Moura, J.F.D.; Giusta, M.S.; De Maria, M.; Álvares, É.S.S.; Lobato, F.C.F.; Assis, R.A.; Gonçalves, L.A.; Gubert, I.C.; et al. Characterization of the venom from the Brazilian Brown Spider Loxosceles similis Moenkhaus, 1898 (Araneae, Sicariidae). Toxicon 2005, 46, 927–936. [Google Scholar] [CrossRef]
- Dos Santos, L.D.; Dias, N.B.; Roberto, J.; Pinto, A.S.; Palma, M.S. Brown recluse spider venom: Proteomic analysis and proposal of a putative mechanism of action. Protein Pept. Lett. 2009, 16, 933–943. [Google Scholar] [CrossRef]
- Trevisan-Silva, D.; Bednaski, A.V.; Fischer, J.S.G.; Veiga, S.S.; Bandeira, N.; Guthals, A.; Marchini, F.K.; Leprevost, F.V.; Barbosa, V.C.; Senff-Ribeiro, A.; et al. A multi-protease, multi-dissociation, bottom-up-to-top-down proteomic view of the Loxosceles intermedia venom. Sci. Data 2017, 4, 1–7. [Google Scholar] [CrossRef]
- Gomez-Muñoz, A.; Gangoiti, P.; Arana, L.; Ouro, A.; Rivera, I.G.; Ordoñez, M.; Trueba, M. New insights on the role of ceramide 1-phosphate in inflammation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2013, 1831, 1060–1066. [Google Scholar] [CrossRef]
- MacEyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nature 2014, 510, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Dragulev, B.; Bao, Y.; Ramos-Cerrillo, B.; Vazquez, H.; Olvera, A.; Stock, R.; Algaron, A.; Fox, J.W. Upregulation of IL-6, IL-8, CXCL1, and CXCL2 dominates gene expression in human fibroblast cells exposed to Loxosceles reclusa sphingomyelinase D: Insights into spider venom dermonecrosis [2]. J. Investig. Dermatol. 2007, 127, 1264–1266. [Google Scholar] [CrossRef]
- Horta, C.C.R.; Oliveira-Mendes, B.B.R.; Do Carmo, A.O.; Siqueira, F.F.; Barroca, T.M.; Dos Santos Nassif Lacerda, S.M.; De Almeida Campos, P.H.; De França, L.R.; Ferreira, R.L.; Kalapothakis, E. Lysophosphatidic acid mediates the release of cytokines and chemokines by human fibroblasts treated with loxosceles spider venom. J. Investig. Dermatol. 2013, 133, 1682–1685. [Google Scholar] [CrossRef] [Green Version]
- Chaves-Moreira, D.; Chaim, O.M.; Sade, Y.B.; Paludo, K.S.; Gremski, L.H.; Donatti, L.; De Moura, J.; Mangili, O.C.; Gremski, W.; Da Silveira, R.B.; et al. Identification of a direct hemolytic effect dependent on the catalytic activity induced by phospholipase-D (dermonecrotic toxin) from brown spider venom. J. Cell. Biochem. 2009, 107, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Stock, R.P.; Brewer, J.; Wagner, K.; Ramos-Cerrillo, B.; Duelund, L.; Jernshøj, K.D.; Olsen, L.F.; Bagatolli, L.A. Sphingomyelinase D activity in model membranes: Structural effects of in situ generation of ceramide-1-phosphate. PLoS ONE 2012, 7, e36003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, R.J.; Campoli, J.; Johar, S.K.; Schumacher, K.A.; Allison, E.J. Suspected brown recluse envenomation: A case report and review of different treatment modalities. J. Emerg. Med. 2011, 41, e31–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isbister, G.K.; Fan, H.W. Spider bite. Lancet 2011, 378, 2039–2047. [Google Scholar] [CrossRef]
- Gomez, H.F.; Greenfield, D.M.; Miller, M.J.; Warren, J.S. Direct Correlation between Diffusion of Loxosceles recluse venom and extent of dermal inflammation. Acad. Emerg. Med. 2001, 8, 309–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauli, I.; Minozzo, J.C.; Henrique da Silva, P.; Chaim, O.M.; Veiga, S.S. Analysis of therapeutic benefits of antivenin at different time intervals after experimental envenomation in rabbits by venom of the brown spider (Loxosceles intermedia). Toxicon 2009, 53, 660–671. [Google Scholar] [CrossRef]
- Floriano, R.S.; Schezaro-Ramos, R.; Silva, N.J.; Bucaretchi, F.; Rowan, E.G.; Hyslop, S. Neurotoxicity of Micrurus lemniscatus lemniscatus (South American coralsnake) venom in vertebrate neuromuscular preparations in vitro and neutralization by antivenom. Arch. Toxicol. 2019, 93, 2065–2086. [Google Scholar] [CrossRef]
- Barbaro, K.C.; Eickstedt, V.R.D.; Mota, I. Antigenic cross-reactivity of venoms from medically important Loxosceles (araneae) species in Brazil. Toxicon 1994, 32, 113–120. [Google Scholar] [CrossRef]
- De Roodt, A.R.; Litwin, S.; Dokmetjian, J.C.; Vidal, J.C. A reduced immunization scheme to obtain an experimental anti-Loxosceles laeta (“Violinist spider”) venom. J. Nat. Toxins 2002, 11, 193–203. [Google Scholar]
- Guilherme, P.; Fernandes, I.; Barbaro, K.C. Neutralization of dermonecrotic and lethal activities and differences among 32-35 kDa toxins of medically important Loxosceles spider venoms in Brazil revealed by monoclonal antibodies. Toxicon 2001, 39, 1333–1342. [Google Scholar] [CrossRef]
- Feitosa, L.; Gremski, W.; Veiga, S.S.; Elias, M.C.Q.B.; Graner, E.; Mangili, O.C.; Brentani, R.R. Detection and characterization of metalloproteinases with gelatinolytic, fibronectinolytic and fibrinogenolytic activities in Brown spider (Loxosceles intermedia) venom. Toxicon 1998, 36, 1039–1051. [Google Scholar] [CrossRef]
- Da Silveira, R.B.; Wille, A.C.M.; Chaim, O.M.; Appel, M.H.; Silva, D.T.; Franco, C.R.C.; Toma, L.; Mangili, O.C.; Gremski, W.; Dietrich, C.P.; et al. Identification, cloning, expression and functional characterization of an astacin-like metalloprotease toxin from Loxosceles intermedia (brown spider) venom. Biochem. J. 2007, 406, 355–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevisan-Silva, D.; Gremski, L.H.; Chaim, O.M.; da Silveira, R.B.; Meissner, G.O.; Mangili, O.C.; Barbaro, K.C.; Gremski, W.; Veiga, S.S.; Senff-Ribeiro, A. Astacin-like metalloproteases are a gene family of toxins present in the venom of different species of the brown spider (genus Loxosceles). Biochimie 2010, 92, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Trevisan-Silva, D.; Gremski, L.H.; Chaim, O.M.; Senff-Ribeiro, A.; Veiga, S.S. Loxosceles Astacin-Like Proteases (LALPs), an Astacin Metalloprotease Family from the Brown Spider Venom. In Handbook of Proteolytic Enzymes; Elsevier: Amsterdam, The Netherlands, 2013; Volume Voume 1, pp. 1081–1086. ISBN 9780123822192. [Google Scholar]
- Sade, Y.B.; Bóia-Ferreira, M.; Gremski, L.H.; Da Silveira, R.B.; Gremski, W.; Senff-Ribeiro, A.; Chaim, O.M.; Veiga, S.S. Molecular cloning, heterologous expression and functional characterization of a novel translationally-controlled tumor protein (TCTP) family member from Loxosceles intermedia (brown spider) venom. Int. J. Biochem. Cell Biol. 2012, 44, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Buch, D.R.; Souza, F.N.; Meissner, G.O.; Morgon, A.M.; Gremski, L.H.; Ferrer, V.P.; Trevisan-Silva, D.; Matsubara, F.H.; Boia-Ferreira, M.; Sade, Y.B.; et al. Brown spider (Loxosceles genus) venom toxins: Evaluation of biological conservation by immune cross-reactivity. Toxicon 2015, 108, 154–166. [Google Scholar] [CrossRef]
- Barbaro, K.C.; Knysak, I.; Martins, R.; Hogan, C.; Winkel, K. Enzymatic characterization, antigenic cross-reactivity and neutralization of dermonecrotic activity of five Loxosceles spider venoms of medical importance in the Americas. Toxicon 2005, 45, 489–499. [Google Scholar] [CrossRef]
- Da Silveira, R.B.; Chaim, O.M.; Mangili, O.C.; Gremski, W.; Dietrich, C.P.; Nader, H.B.; Veiga, S.S. Hyaluronidases in Loxosceles intermedia (Brown spider) venom are endo-β-N-acetyl-d-hexosaminidases hydrolases. Toxicon 2007, 49, 758–768. [Google Scholar] [CrossRef]
- Ferrer, V.P.; de Mari, T.L.; Gremski, L.H.; Trevisan Silva, D.; da Silveira, R.B.; Gremski, W.; Chaim, O.M.; Senff-Ribeiro, A.; Nader, H.B.; Veiga, S.S. A Novel Hyaluronidase from Brown Spider (Loxosceles intermedia) Venom (Dietrich’s Hyaluronidase): From Cloning to Functional Characterization. PLoS Negl. Trop. Dis. 2013, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Coronado, M.A.; Ullah, A.; Silva, L.S.; Chaves-Moreira, D.; Vuitika, L.; Chaim, O.M.; Veiga, S.S.; Chahine, J.; Murakami, M.T.; Arni, R.K. Structural Insights into Substrate Binding of Brown Spider Venom Class II Phospholipases D. Curr. Protein Pept. Sci. 2015, 16, 768–774. [Google Scholar] [CrossRef]
- Mariutti, R.B.; Chaves-Moreira, D.; Vuitika, L.; Caruso, Í.P.; Coronado, M.A.; Azevedo, V.A.; Murakami, M.T.; Veiga, S.S.; Arni, R.K. Bacterial and Arachnid Sphingomyelinases D: Comparison of Biophysical and Pathological Activities. J. Cell. Biochem. 2017, 118, 2053–2063. [Google Scholar] [CrossRef]
- Pace, L.B.; Vetter, R.S. Brown recluse spider (Loxosceles reclusa) envenomation in small animals. J. Vet. Emerg. Crit. Care 2009, 19, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Felicori, L.; Fernandes, P.B.; Giusta, M.S.; Duarte, C.G.; Kalapothakis, E.; Nguyen, C.; Molina, F.; Granier, C.; Chávez-Olórtegui, C. An in vivo protective response against toxic effects of the dermonecrotic protein from Loxosceles intermedia spider venom elicited by synthetic epitopes. Vaccine 2009, 27, 4201–4208. [Google Scholar] [CrossRef] [PubMed]
- de Moura, J.; Felicori, L.; Moreau, V.; Guimarães, G.; Dias-Lopes, C.; Molina, L.; Alvarenga, L.M.; Fernandes, P.; Frézard, F.; Ribeiro, R.R.; et al. Protection against the toxic effects of Loxosceles intermedia spider venom elicited by mimotope peptides. Vaccine 2011, 29, 7992–8001. [Google Scholar] [CrossRef] [PubMed]
- Ramada, J.S.; Becker-Finco, A.; Minozzo, J.C.; Felicori, L.F.; Machado de Avila, R.A.; Molina, F.; Nguyen, C.; de Moura, J.; Chávez-Olórtegui, C.; Alvarenga, L.M. Synthetic peptides for invitro evaluation of the neutralizing potency of Loxosceles antivenoms. Toxicon 2013, 73, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.D.A.; Guerra-Duarte, C.; Costal-Oliveira, F.; Mendes, T.M.; Luís, L.F.; Oliveira, D.; de Avila, R.A.M.; Ferrer, V.P.; Trevisan-Silva, D.; Veiga, S.S.; et al. Recombinant protein containing B-cell epitopes of different Loxosceles spider toxins generates neutralizing antibodies in immunized rabbits. Front. Immunol. 2018, 9, 653. [Google Scholar] [CrossRef] [Green Version]
- Bermúdez-Méndez, E.; Fuglsang-Madsen, A.; Føns, S.; Lomonte, B.; Gutiérrez, J.M.; Laustsen, A.H. Innovative immunization strategies for antivenom development. Toxins 2018, 10, 452. [Google Scholar] [CrossRef] [Green Version]
- Van Meeteren, L.A.; Stortelers, C.; Moolenaar, W.H. Upregulation of cytokine expression in fibroblasts exposed to Loxosceles sphingomyelinase D: What is the trigger? J. Investig. Dermatol. 2007, 127, 1266–1267. [Google Scholar] [CrossRef] [Green Version]
- Anliker, B.; Chun, J. Lysophospholipid G protein-coupled receptors. J. Biol. Chem. 2004, 279, 20555–20558. [Google Scholar] [CrossRef] [Green Version]
- Chalfant, C.E.; Spiegel, S. Sphingosine 1-phosphate and ceramide 1-phosphate: Expanding roles in cell signaling. J. Cell Sci. 2005, 118, 4605–4612. [Google Scholar] [CrossRef] [Green Version]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef]
- Chaves-Moreira, D.; de Moraes, F.R.; Caruso, Í.P.; Chaim, O.M.; Senff-Ribeiro, A.; Ullah, A.; da Silva, L.S.; Chahine, J.; Arni, R.K.; Veiga, S.S. Potential Implications for Designing Drugs Against the Brown Spider Venom Phospholipase-D. J. Cell. Biochem. 2017, 118, 726–738. [Google Scholar] [CrossRef]
- Paixão-Cavalcante, D.; Van Den Berg, C.W.; Gonçalves-De-Andrade, R.M.; Fernandes-Pedrosa, M.D.F.; Okamoto, C.K.; Tambourgi, D.V. Tetracycline protects against dermonecrosis induced by Loxosceles spider venom. J. Investig. Dermatol. 2007, 127, 1410–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, C.K.; van den Berg, C.W.; Masashi, M.; Gonçalves-de-Andrade, R.M.; Tambourgi, D.V. Tetracycline reduces kidney damage induced by loxosceles spider venom. Toxins 2017, 9, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Medeiros, C.R.; Brioschi, M.L.; de Souza, S.N.; Teixeira, M.J. Infrared thermography to diagnose and manage venomous animal bites and stings. Rev. Soc. Bras. Med. Trop. 2017, 50, 260–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, S.M.; Romine-Jenkins, M.; Blick, K.E. Passive hemagglutination inhibition test for diagnosis of brown recluse spider bite envenomation. Clin. Chem. 1993, 39, 2104–2107. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Olórtegui, C.; Zanetti, V.C.; Ferreira, A.P.; Minozzo, J.C.; Mangili, O.C.; Gubert, I.C. ELISA for the detection of venom antigens in experimental and clinical envenoming by Loxosceles intermedia spiders. Toxicon 1998, 36, 563–569. [Google Scholar] [CrossRef]
- Miller, M.J.; Gomez, H.F.; Snider, R.J.; Stephens, E.L.; Czop, R.M.; Warren, J.S. Detection of Loxosceles venom in lesional hair shafts and skin: Application of a specific immunoassay to identify dermonecrotic arachnidism. Am. J. Emerg. Med. 2000, 18, 626–628. [Google Scholar] [CrossRef]
- Gomez, H.F.; Krywko, D.M.; Stoecker, W.V. A new assay for the detection of Loxosceles species (brown recluse) spider venom. Ann. Emerg. Med. 2002, 39, 469–474. [Google Scholar] [CrossRef] [Green Version]
- Krywko, D.M.; Gomez, H.F. Detection of Loxosceles species venom in dermal lesions: A comparison of 4 venom recovery methods. Ann. Emerg. Med. 2002, 39, 475–480. [Google Scholar] [CrossRef]
- Stoecker, W.V.; Green, J.A.; Gomez, H.F. Diagnosis of loxoscelism in a child confirmed with an enzyme-linked immunosorbent assay and noninvasive tissue sampling. J. Am. Acad. Dermatol. 2006, 55, 888–890. [Google Scholar] [CrossRef] [Green Version]
- Akdeniz, S.; Green, J.A.; Stoecker, W.V.; Gomez, H.F.; Keklikçi, S.U. Diagnosis of loxoscelism in two Turkish patients confirmed with an enzyme-linked immunosorbent assay (ELISA) and non-invasive tissue sampling. Dermatol. Online J. 2007, 13, 11. [Google Scholar]
- Stoecker, W.V.; Wasserman, G.S.; Calcara, D.A.; Green, J.A.; Larkin, K. Systemic loxoscelism confirmation by bite-site skin surface: ELISA. Mo. Med. 2009, 106, 425. [Google Scholar] [PubMed]
- McGlasson, D.L.; Green, J.A.; Stoecker, W.V.; Babcock, J.L.; Calcara, D.A. Duration of Loxosceles reclusa venom detection by ELISA from swabs. Clin. Lab. Sci. 2009, 22, 216–222. [Google Scholar] [PubMed]
- Dias-Lopes, C.; Paiva, A.L.; Guerra-Duarte, C.; Molina, F.; Felicori, L. Venomous arachnid diagnostic assays, lessons from past attempts. Toxins 2018, 10, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arán-Sekul, T.; Rojas, J.M.; Subiabre, M.; Cruz, V.; Cortés, W.; Osorio, L.; González, J.; Araya, J.E.; Catalán, A. Heterophilic antibodies in sera from individuals without loxoscelism cross-react with phospholipase D from the venom of Loxosceles and Sicarius spiders. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 18. [Google Scholar] [CrossRef] [PubMed]
- Jiacomini, I.; Silva, S.K.; Aubrey, N.; Muzard, J.; Chavez-Olortegui, C.; De Moura, J.; Billiald, P.; Alvarenga, L.M. Immunodetection of the “brown” spider (Loxosceles intermedia) dermonecrotoxin with an scFv-alkaline phosphatase fusion protein. Immunol. Lett. 2016, 173, 1–6. [Google Scholar] [CrossRef]
- Dos Santos, V.L.P.; Franco, C.R.C.; Viggiano, R.L.L.; Da Silveira, R.B.; Cantão, M.P.; Mangili, O.C.; Veiga, S.S.; Gremski, W. Structural and ultrastructural description of the venom gland of Loxosceles intermedia (brown spider). Toxicon 2000, 38, 265–285. [Google Scholar] [CrossRef]
- Sapag, A.; Salinas-Luypaert, C.; Constenla-Muñoz, C. First report of in vitro selection of RNA aptamers targeted to recombinant Loxosceles laeta spider toxins. Biol. Res. 2014, 47, 2. [Google Scholar] [CrossRef] [Green Version]
- Marsh, N.; Williams, V. Practical applications of snake venom toxins in haemostasis. Toxicon 2005, 45, 1171–1181. [Google Scholar] [CrossRef]
- Heine, R.J.; Van Gaal, L.F.; Johns, D.; Mihm, M.J.; Widel, M.H.; Brodows, R.G. Exenatide versus insulin glargine in patients with suboptimally controlled type 2 diabetes: A randomized trial. Ann. Intern. Med. 2005, 143, 559–569. [Google Scholar] [CrossRef]
- Koh, D.C.I.; Armugam, A.; Jeyaseelan, K. Snake venom components and their applications in biomedicine. Cell. Mol. Life Sci. 2006, 63, 3030–3041. [Google Scholar] [CrossRef] [PubMed]
- Mirshafiey, A. Venom therapy in multiple sclerosis. Neuropharmacology 2007, 53, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Schmidtko, A.; Lötsch, J.; Freynhagen, R.; Geisslinger, G. Ziconotide for treatment of severe chronic pain. Lancet 2010, 375, 1569–1577. [Google Scholar] [CrossRef]
- Chaim, O.M.; Trevisan-Silva, D.; Chaves-Moreira, D.; Wille, A.C.M.; Ferrer, V.P.; Matsubara, F.H.; Mangili, O.C.; da Silveira, R.B.; Gremski, L.H.; Gremski, W.; et al. Brown spider (Loxosceles genus) venom toxins: Tools for biological purposes. Toxins 2011, 3, 309–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norton, R.; Pennington, M.; Wulff, H. Potassium Channel Blockade by the Sea Anemone Toxin ShK for the Treatment of Multiple Sclerosis and Other Autoimmune Diseases. Curr. Med. Chem. 2012, 11, 3041–3052. [Google Scholar] [CrossRef]
- Tambourgi, D.V.; De Sousa Da Silva, M.; Billington, S.J.; Gonçalves De Andrade, R.M.; Magnoli, F.C.; Songer, J.G.; Van Den Berg, C.W. Mechanism of induction of complement susceptibility of erythrocytes by spider and bacterial sphingomyelinases. Immunology 2002, 107, 93–101. [Google Scholar] [CrossRef]
- Murph, M.; Tanaka, T.; Liu, S.; Mills, G.B. Of spiders and crabs: The emergence of lysophospholipids and their metabolic pathways as targets for therapy in cancer. Clin. Cancer Res. 2006, 12, 6598–6602. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.H.; Han, J.S. Phospholipase D and its essential role in cancer. Mol. Cells 2017, 40, 805–813. [Google Scholar] [PubMed]
- Houben, A.J.S.; Moolenaar, W.H. Autotaxin and LPA receptor signaling in cancer. Cancer Metastasis Rev. 2011, 30, 557–565. [Google Scholar] [CrossRef]
- World Spider Catalog (2020). World Spider Catalog. Version 21.0. Natural History Museum Bern. Available online: http://wsc.nmbe.ch (accessed on 10 February 2020). [CrossRef]
- Vetter, R.S. The distribution of brown recluse spiders in the southeastern quadrant of the United States in relation to loxoscelism diagnoses. South. Med. J. 2009, 102, 518–522. [Google Scholar] [CrossRef]
- Cordeiro, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Arantes, E.C. Arachnids of medical importance in Brazil: Main active compounds present in scorpion and spider venoms and tick saliva. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]





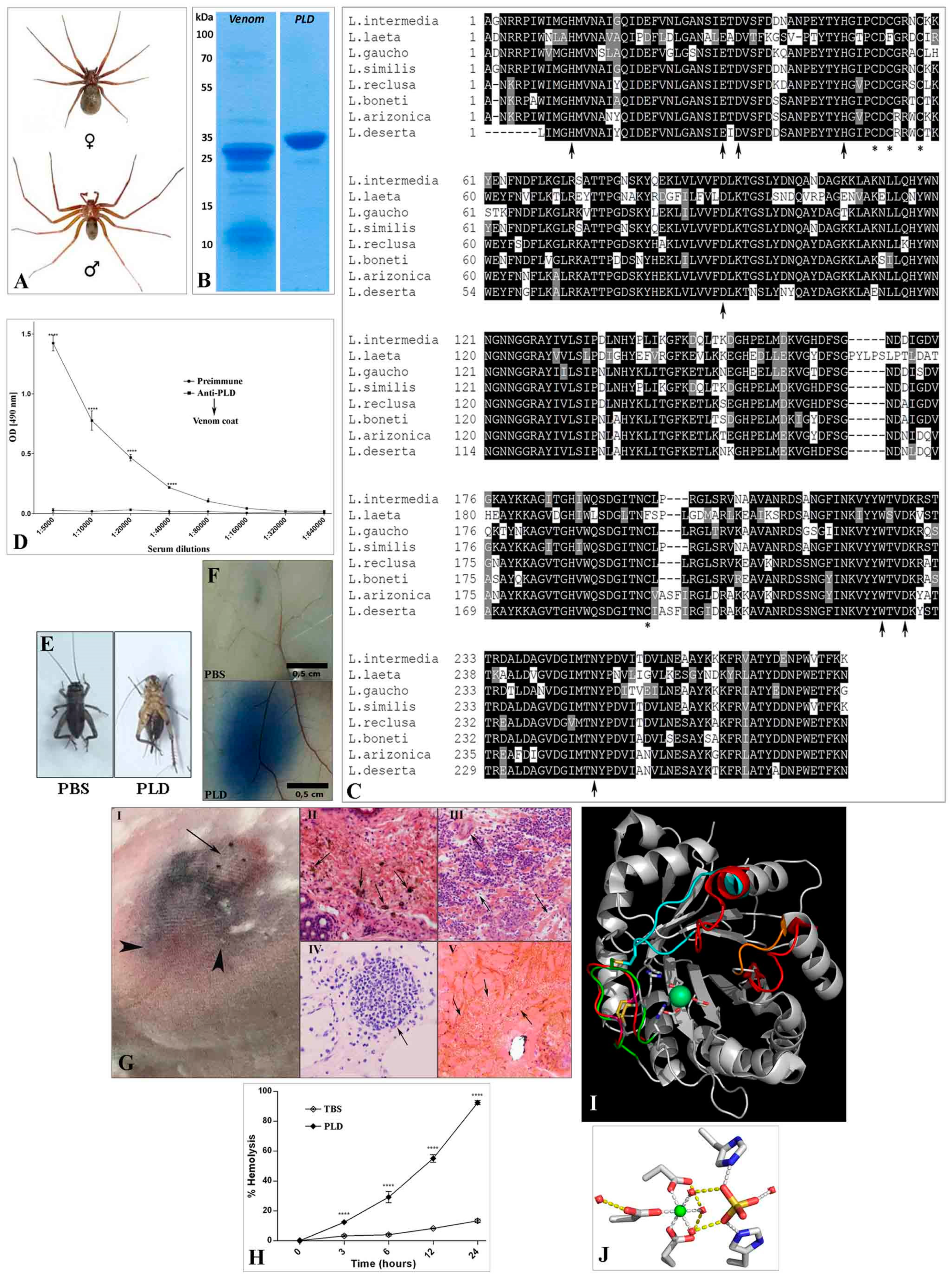{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recombinant Loxosceles PLDs | Known Activity | References | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Name | GenBank Acession Number | Dermonecrosis | Hemolysis | Sphingo-myelinase | In vitro Platelet Aggregation | Vasc. Perm. Increase | Inflammat. | Lethality | Nephrotox | Edema | Cardiotox | Insecticidal | |
| L. intermedia | LiRecDT1; LiSicTox-alphaIA1a | ABA62021, P0CE80 | Yes | Yes (Direct/C-dep) | Yes | Yes | Yes | Yes | Yes | Yes | Yes | − | Yes | [9,47,53,54], This work |
| LiD1 | AAQ16123 | Yes | Yes (C-dep) | Yes | Yes | − | − | − | − | Yes | Yes | − | [45,55,56,57] | |
| LiRecDT2; LiSicTox-alphaIA2aii | ABB69098, P0CE83 | Yes | Yes (Direct) | Yes | Yes | Yes | Yes | Yes | − | Yes | − | − | [13,15,54] | |
| LiRecDT3 | ABB71184 | No | No | No | No | No | ± | No | − | No | − | − | [13,15] | |
| LiRecDT4 | ABD91846 | Yes | − | Yes | ± | Yes | Yes | − | − | Yes | − | − | [14] | |
| LiRecDT5 | ABD91847 | No | − | No | No | No | ± | − | − | Yes | − | − | [14] | |
| LiRecDT6 | ABO87656 | Yes | − | Yes | Yes | Yes | Yes | Yes | − | Yes | − | − | [48] | |
| LiRecDT7 | AGN52903 | Yes | Yes (Direct) | Yes | − | Yes | Yes | − | − | − | − | [16] | ||
| L. laeta | Smase I/ LlSicTox-alphaIII1i | AAM21154 | Yes | Yes (C-dep) | Yes | − | − | Yes | − | − | Yes | − | − | [18,46] |
| Ll1/LlSicTox-alphaIII1ii | ABD15447 | − | − | Yes | − | − | − | Yes | − | − | − | − | [51] | |
| LlPLD1 | ADP00408 | − | Yes (C-dep) | Yes | − | − | Yes | − | − | − | − | − | [52,58] | |
| LlPLD2 | ADP00409 | − | No | No | − | − | Yes | − | − | − | − | − | [52,58] | |
| Ll2/LlSicTox-alphaIII2 | ABD15448 | − | − | Yes | − | − | − | Yes | − | − | − | − | [51] | |
| Smase II/ LlSicTox-betaIA1 | AAM21156 | ± | Yes (C-dep) | Yes | − | − | Yes | − | − | Yes | − | − | [18,46] | |
| L. reclusa | Smase D | AAW56831 | − | Yes (C-dep) | Yes | − | − | − | − | − | − | − | − | [27] |
| Lr1 | AAT66075 | − | − | Yes | − | − | − | Yes | − | − | − | − | [51] | |
| L. boneti | Lb1 | AAT66073 | − | − | Yes | − | − | − | Yes | − | − | − | − | [51] |
| Lb3 | AAT66074 | − | − | No | − | − | − | − | − | − | − | − | [51] | |
| L. arizonica | LarSicTox-alphaIB2bi | Q4ZFU2 | − | − | Yes | − | − | − | − | − | − | − | Yes | [12] |
| LarSicTox-betaID1 | AJV88487 | − | − | Yes | − | − | − | − | − | − | − | − | [30] | |
| L. gaucho | LgRec1 | AFY98967 | Yes | Yes (Direct) | Yes | Yes | − | − | − | − | Yes | − | − | [53] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gremski, L.H.; da Justa, H.C.; da Silva, T.P.; Polli, N.L.C.; Antunes, B.C.; Minozzo, J.C.; Wille, A.C.M.; Senff-Ribeiro, A.; Arni, R.K.; Veiga, S.S. Forty Years of the Description of Brown Spider Venom Phospholipases-D. Toxins 2020, 12, 164. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030164
Gremski LH, da Justa HC, da Silva TP, Polli NLC, Antunes BC, Minozzo JC, Wille ACM, Senff-Ribeiro A, Arni RK, Veiga SS. Forty Years of the Description of Brown Spider Venom Phospholipases-D. Toxins. 2020; 12(3):164. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030164
Chicago/Turabian StyleGremski, Luiza Helena, Hanna Câmara da Justa, Thaís Pereira da Silva, Nayanne Louise Costacurta Polli, Bruno César Antunes, João Carlos Minozzo, Ana Carolina Martins Wille, Andrea Senff-Ribeiro, Raghuvir Krishnaswamy Arni, and Silvio Sanches Veiga. 2020. "Forty Years of the Description of Brown Spider Venom Phospholipases-D" Toxins 12, no. 3: 164. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030164