
Evaluation of the Toxicity and Toxicokinetics of Cereulide from an Emetic Bacillus cereus Strain of Milk Origin

Abstract
:
1. Introduction
2. Results
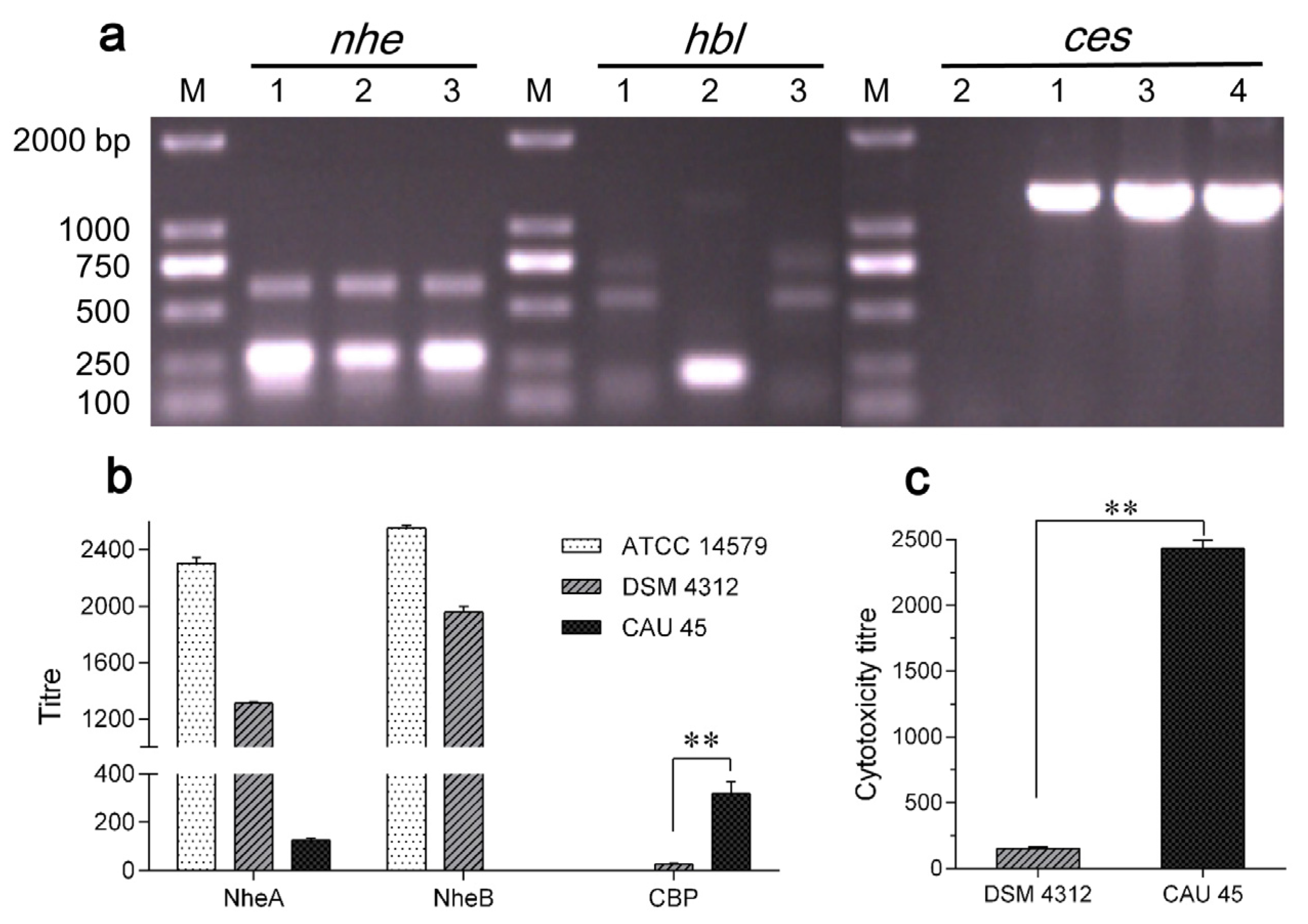
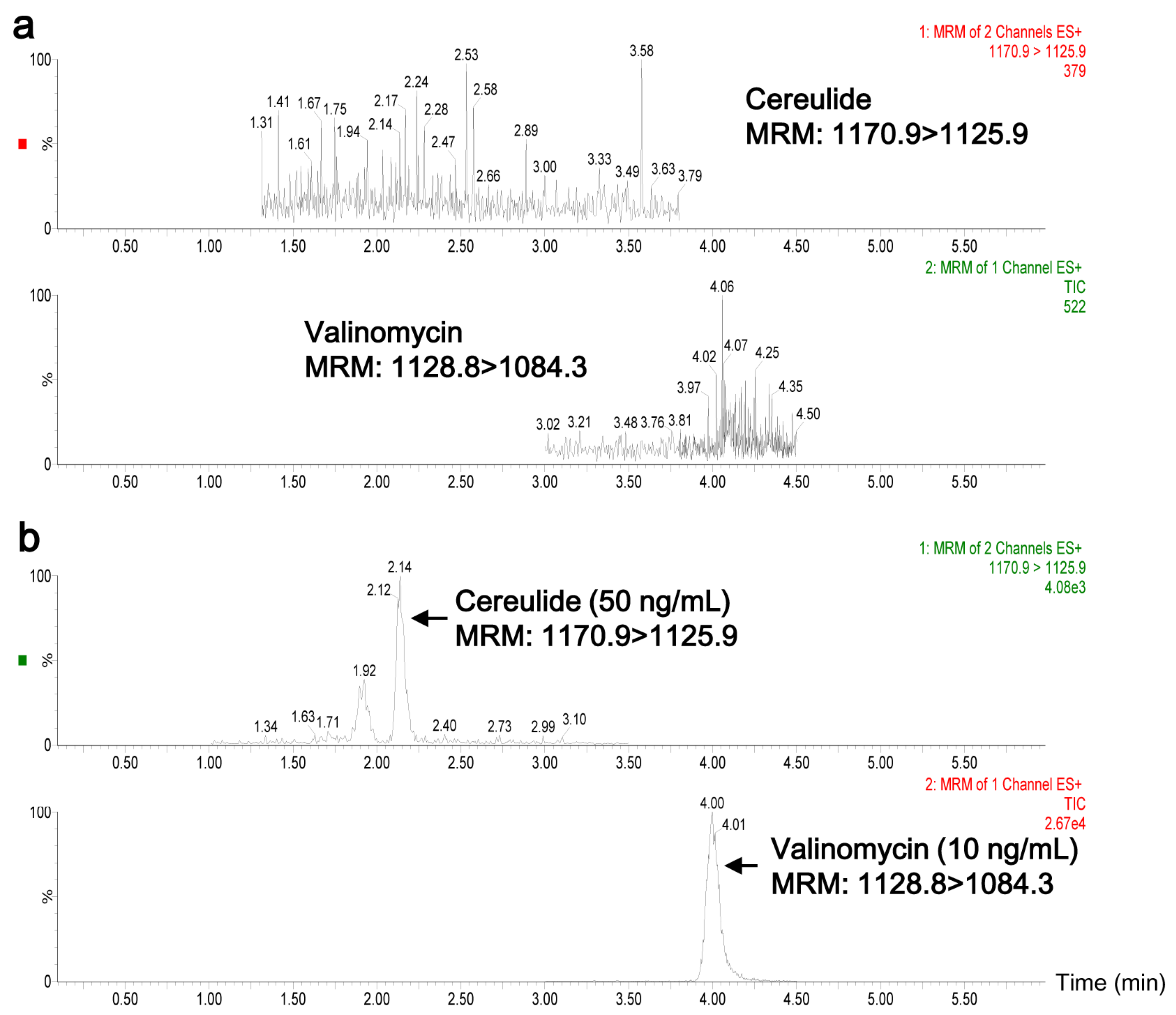
2.1. Characterization of B. cereus CAU45
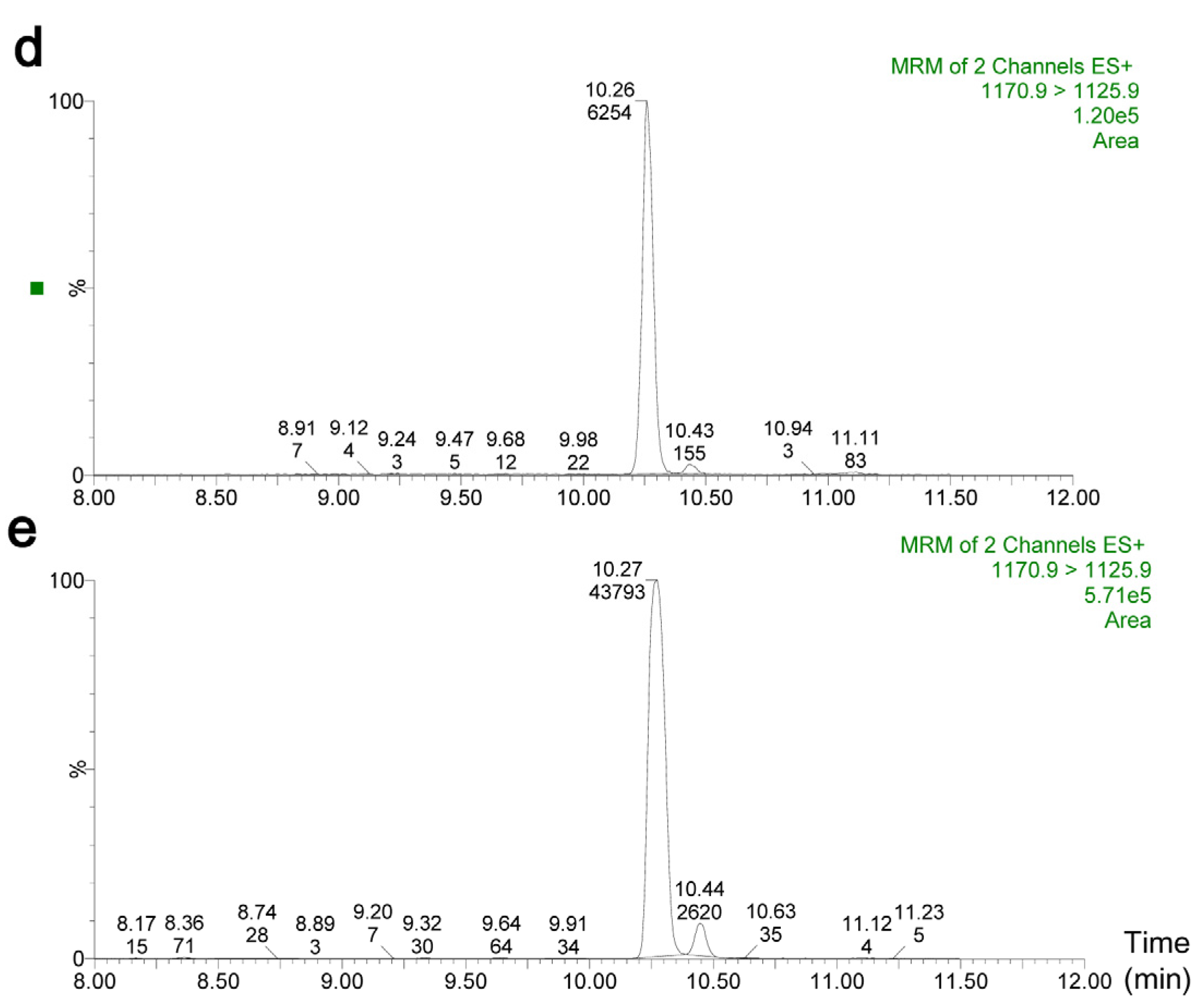
2.2. Quantification of Cereulide in Plasma by UPLC-MS/MS
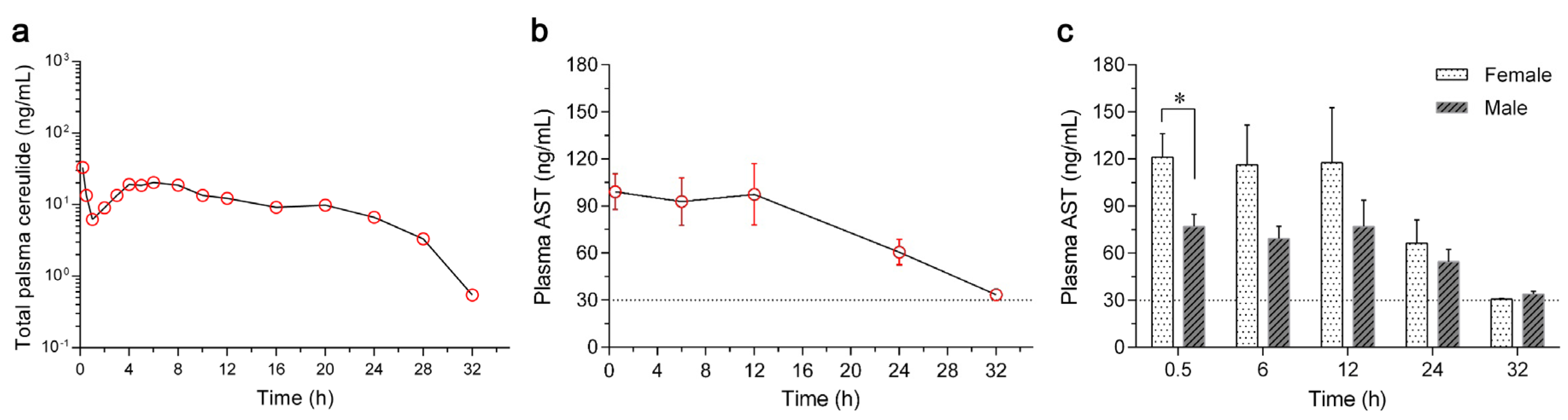
2.3. Toxicokinetics of Cereulide in Rabbits
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Materials
4.2. Isolation and Identification the Emetic B. cereus Strain
4.3. Extraction of Cereulide
4.4. Cereulide Treated Rabbits
4.5. Quantification of Cereulide in Plasma by UPLC-MS/MS
4.6. AST Measurement
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bottone, E.J. Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 2010, 23, 382–398. [Google Scholar] [CrossRef] [PubMed]
- Ceuppens, S.; Boon, N.; Uyttendaele, M. Diversity of Bacillus cereus group strains is reflected in their broad range of pathogenicity and diverse ecological lifestyles. FEMS Microbiol. Ecol. 2013, 84, 433–450. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Acaroez, U.; Maertlbauer, E. A cellular logic circuit for the detection of bacterial pore-forming toxins. Chem. Commun. 2013, 49, 5198–5200. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Didier, A.; Dietrich, R.; Heilkenbrinker, U.; Waltenberger, E.; Jessberger, N.; Martlbauer, E.; Benz, R. Formation of small transmembrane pores: An intermediate stage on the way to Bacillus cereus non-hemolytic enterotoxin (Nhe) full pores in the absence of NheA. Biochem. Biophys. Res. Commun. 2016, 469, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Mahler, H.; Pasi, A.; Kramer, J.M.; Schulte, P.; Scoging, A.C.; Bär, W.; Krähenbühl, S. Fulminant liver failure in association with the emetic toxin of Bacillus cereus. N. Engl. J. Med. 1997, 336, 1142–1148. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, M.; Denayer, S.; Botteldoorn, N.; Delbrassinne, L.; Veys, J.; Waegenaere, J.; Sirtaine, N.; Driesen, R.B.; Sipido, K.R.; Mahillon, J.; et al. Sudden death of a young adult associated with Bacillus cereus food poisoning. J. Clin. Microbiol. 2011, 49, 4379–4381. [Google Scholar] [CrossRef] [PubMed]
- Eurosurveillance Editorial Team. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2011. Eurosurveillance 2013, 18. [Google Scholar] [CrossRef]
- Wang, S.J.; Duan, H.L.; Zhang, W.; Li, J.W. Analysis of bacterial foodborne disease outbreaks in China between 1994 and 2005. FEMS Immunol. Med. Microbiol. 2007, 51, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Rajkovic, A.; Uyttendaele, M.; Vermeulen, A.; Andjelkovic, M.; Fitz-James, I.; in’t Veld, P.; Denon, Q.; Verhe, R.; Debevere, J. Heat resistance of Bacillus cereus emetic toxin, cereulide. Lett. Appl. Microbiol. 2008, 46, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Messelhausser, U.; Frenzel, E.; Blochinger, C.; Zucker, R.; Kampf, P.; Ehling-Schulz, A. Emetic Bacillus cereus are more volatile than thought: Recent foodborne outbreaks and prevalence studies in Bavaria (2007–2013). Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.P.; Liu, H.Z.; He, J.; Yuan, Y.M.; Yuan, Z.M. The occurrence of Bacillus cereus, B. thuringiensis and B. mycoides in Chinese pasteurized full fat milk. Int. J. Food Microbiol. 2008, 121, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.P.; Zheng, D.S.; Dou, L.; Cai, Q.X.; Yuan, Z.M. Occurrence of psychrotolerant Bacillus cereus group strains in ice creams. Int. J. Food Microbiol. 2010, 137, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Marxen, S.; Stark, T.D.; Rutschle, A.; Lucking, G.; Frenzel, E.; Scherer, S.; Ehling-Schulz, M.; Hofmann, T. Depsipeptide intermediates interrogate proposed biosynthesis of cereulide, the emetic toxin of Bacillus cereus. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Makarasen, A.; Yoza, K.; Isobe, M. Higher structure of cereulide, an emetic toxin from Bacillus cereus, and special comparison with valinomycin, an antibiotic from streptomyces fulvissimus. Chem. Asian J. 2009, 4, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Shiota, M.; Saitou, K.; Mizumoto, H.; Matsusaka, M.; Agata, N.; Nakayama, M.; Kage, M.; Tatsumi, S.; Okamoto, A.; Yamaguchi, S.; et al. Rapid detoxification of cereulide in Bacillus cereus food poisoning. Pediatrics 2010, 125, E951–E955. [Google Scholar] [CrossRef] [PubMed]
- Agata, N.; Ohta, M.; Mori, M.; Isobe, M. A novel dodecadepsipeptide, cereulide, is an emetic toxin of Bacillus cereus. FEMS Microbiol. Lett. 1995, 129, 17–19. [Google Scholar] [PubMed]
- Shinagawa, K.; Konuma, H.; Sekita, H.; Sugii, S. Emesis of rhesus monkeys induced by intragastric administration with the HEp-2 vacuolation factor (cereulide) produced by Bacillus cereus. FEMS Microbiol. Lett. 1995, 130, 87–90. [Google Scholar] [PubMed]
- Kumari, S.; Sarkar, P.K. Prevalence and characterization of Bacillus cereus group from various marketed dairy products in India. Dairy Sci. Technol. 2014, 94, 483–497. [Google Scholar] [CrossRef]
- Zhou, G.P.; Bester, K.; Liao, B.; Yang, Z.S.; Jiang, R.R.; Hendriksen, N.B. Characterization of three Bacillus cereus strains involved in a major outbreak of food poisoning after consumption of fermented black beans (Douchi) in Yunan, China. Foodborne Pathog. Dis. 2014, 11, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Karagoz, K.; Adiguzel, G.; Dikbas, N. Emetic and enterotoxin profiles of food-borne Bacillus cereus strains from Turkey. Res. J. Biotechnol. 2015, 10, 44–49. [Google Scholar]
- Ronning, H.T.; Asp, T.N.; Granum, P.E. Determination and quantification of the emetic toxin cereulide from Bacillus cereus in pasta, rice and cream with liquid chromatography-tandem mass spectrometry. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Yim, J.H.; Kim, K.Y.; Chon, J.W.; Kim, D.H.; Kim, H.S.; Choi, D.S.; Choi, I.S.; Seo, K.H. Incidence, antibiotic susceptibility, and toxin profiles of Bacillus cereus sensu lato isolated from Korean fermented soybean products. J. Food Sci. 2015, 80, M1266–M1270. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Frenzel, E.; Gohar, M. Food-bacteria interplay: Pathometabolism of emetic Bacillus cereus. Front. Microbiol. 2015, 6, 704. [Google Scholar] [CrossRef] [PubMed]
- Svensson, B.; Monthan, A.; Shaheen, R.; Andersson, M.A.; Salkinoja-Salonen, M.; Christiansson, A. Occurrence of emetic toxin producing Bacillus cereus in the dairy production chain. Int. Dairy J. 2006, 16, 740–749. [Google Scholar] [CrossRef]
- Kim, J.B.; Jeong, H.R.; Park, Y.B.; Kim, J.M.; Oh, D.H. Food poisoning associated with emetic-type of Bacillus cereus in Korea. Foodborne Pathog. Dis. 2010, 7, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Fricker, M.; Grallert, H.; Rieck, P.; Wagner, M.; Scherer, S. Cereulide synthetase gene cluster from emetic Bacillus cereus: Structure and location on a mega virulence plasmid related to Bacillus anthracis toxin plasmid pXOI. BMC Microbiol. 2006, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Dietrich, R.; Didier, A.; Acar, G.; Maertlbauer, E. Versatile antibody-sensing Boolean logic for the simultaneous detection of multiple bacterial toxins. Chem. Commun. 2013, 49, 9314–9316. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Fricker, M.; Scherer, S. Bacillus cereus, the causative agent of an emetic type of food-borne illness. Mol. Nutr. Food Res. 2004, 48, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, K.; Ito, M.; Agata, N.; Isobe, M.; Shibayama, K.; Horii, T.; Ohta, M. Pathological effect of synthetic cereulide, an emetic toxin of Bacillus cereus, is reversible in mice. FEMS Immunol. Med. Microbiol. 1999, 24, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.F.; Liu, X.Y.; Dietrich, R.; Martlbauer, E.; Cao, J.; Ding, S.Y.; Zhu, Y. Characterization of Bacillus cereus isolates from local dairy farms in China. FEMS Microbiol. Lett. 2016. [Google Scholar] [CrossRef]
- Merzougui, S.; Lkhider, M.; Grosset, N.; Gautier, M.; Cohen, N. Prevalence, PFGE typing, and antibiotic resistance of Bacillus cereus group isolated from food in Morocco. Foodborne Pathog. Dis. 2014, 11, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Wehrle, E.; Moravek, M.; Dietrich, R.; Burk, C.; Didier, A.; Martlbauer, E. Comparison of multiplex PCR, enzyme immunoassay and cell culture methods for the detection of enterotoxinogenic Bacillus cereus. J. Microbiol. Methods 2009, 78, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, R.; Moravek, M.; Burk, C.; Granum, P.E.; Martlbauer, E. Production and characterization of antibodies against each of the three subunits of the Bacillus cereus nonhemolytic enterotoxin complex. Appl. Environ. Microbiol. 2005, 71, 8214–8220. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Vukov, N.; Schulz, A.; Shaheen, R.; Andersson, M.; Martlbauer, E.; Scherer, S. Identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus. Appl. Environ. Microbiol. 2005, 71, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Bauer, T.; Stark, T.; Hofmann, T.; Ehling-Schulz, M. Development of a stable isotope dilution analysis for the quantification of the Bacillus cereus toxin cereulide in foods. J. Agric. Food Chem. 2010, 58, 1420–1428. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxicokinetic Parameters * | Median | Range | Mean ± SD |
|---|---|---|---|
| AUC (h × ng/mL) | 309.3 | 81.2–851.1 | 320.9 ± 240.1 |
| AUC∞ (h × ng/mL) | 330.5 | 126.0–853.9 | 390.3 ± 257.9 |
| Cl (mL/h) | 15.5 | 5.9–39.7 | 19.1 ± 12.8 |
| Cmax (ng/mL) | 33.8 | 13.3–78.1 | 40.8 ± 21.6 |
| HL_Lambda_z (h) | 6.9 | 1.9–24.5 | 10.8 ± 9.1 |
| Lambda_z (1/h) | 0.1 | 0.03–0.4 | 0.1 ± 0.1 |
| MRTlast (h) | 10.6 | 4.5–13.4 | 9.6 ± 2.9 |
| Tmax (h) | 0.2 | 0.2–8.0 | 2.6 ± 3.4 |
| Vd (mL) | 208.1 | 16.3–1254.5 | 320.1 ± 399.6 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Y.; Liu, Y.; Liu, X.; Xia, X.; Ding, S.; Zhu, K. Evaluation of the Toxicity and Toxicokinetics of Cereulide from an Emetic Bacillus cereus Strain of Milk Origin. Toxins 2016, 8, 156. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8060156
Cui Y, Liu Y, Liu X, Xia X, Ding S, Zhu K. Evaluation of the Toxicity and Toxicokinetics of Cereulide from an Emetic Bacillus cereus Strain of Milk Origin. Toxins. 2016; 8(6):156. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8060156
Chicago/Turabian StyleCui, Yifang, Yuan Liu, Xiaoye Liu, Xi Xia, Shuangyang Ding, and Kui Zhu. 2016. "Evaluation of the Toxicity and Toxicokinetics of Cereulide from an Emetic Bacillus cereus Strain of Milk Origin" Toxins 8, no. 6: 156. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8060156