

Abstract
Objective. Ankylosing spondylitis (AS) is characterized by chronic inflammation of the axial and peripheral joints and ligamentous attachments. Gut immunity is thought to be involved in AS, because a prominent coexistence of gut and joint inflammation has been observed in patients with AS. Mucosal-associated invariant T (MAIT) cells are preferentially located in the gut lamina propria and produce inflammatory cytokines such as interleukin 17 (IL-17) and tumor necrosis factor-α (TNF-α), which are therapeutic targets for AS. This study aimed to investigate the involvement of MAIT cells in AS.
Methods. The frequency of MAIT cells and their cytokine production were determined in patients with AS and healthy controls (HC). The expression of a MAIT cell activation marker (CD69) was analyzed in patients with AS by using flow cytometry.
Results. The frequency of MAIT cells in the peripheral blood was lower in patients with AS compared with HC. The levels of IL-17 produced by MAIT cells after activation were higher in patients with AS than in the HC. CD69 expression on MAIT cells correlated with the Ankylosing Spondylitis Disease Activity Score in patients with AS.
Conclusion. These results suggest the involvement of MAIT cells in the pathogenesis of AS.
Ankylosing spondylitis (AS) is characterized by chronic inflammation of the axial and peripheral joints and ligamentous attachments and the absence of rheumatoid factor. Although the pathogenesis of AS is poorly understood, both genetic and environmental factors are thought to be involved1. Gut immunity was suggested to be involved in AS because a prominent coexistence of gut and joint inflammation was present in patients with AS2,3,4. In addition, the genetic predisposition of AS is similar to that of Crohn disease (CD)5. Thus, mucosal immunity might have an important role in the development and progression of AS.
Mucosal-associated invariant T (MAIT) cells are preferentially located in the gut lamina propria and produce inflammatory cytokines such as interleukin 17 (IL-17) and tumor necrosis factor-α (TNF-α), which are therapeutic targets for AS6,7,8,9,10,11. Therefore, researchers have been increasingly interested in the function of MAIT cells in infection and inflammation12–21,22,23. MAIT cells express an invariant T cell receptor α (TCRα) chain: Vα7.2-Jα33 in humans and Vα19-Jα33 in mice. The invariant TCRα chain is associated with a limited set of Vβ chains6,7. MAIT cells are restricted by MHC-related molecule 1 (MR1), a nonpolymorphic MHC class 1b molecule, and their selection in the thymus is MR1-dependent7. MAIT cells require B cells as well as commensal microbiota for their peripheral expansion7. Vitamin derivatives that originate from the bacterial riboflavin biosynthetic pathway were identified as antigens for MAIT cells24. We previously reported that MAIT cells play a pathogenic role in murine models of arthritis, including collagen-induced arthritis and antibody-mediated arthritis14. In a study by Cho, et al, MAIT cells were found to be involved in rheumatoid arthritis (RA)15.
In our current study, we showed that the frequency of MAIT cells was low in the peripheral blood of patients with AS compared with healthy controls (HC). The production of IL-17 from MAIT cells after activation was higher in patients with AS than in the HC. The expression of CD69 on MAIT cells correlated with the Ankylosing Spondylitis Disease Activity Score (ASDAS) in patients with AS. The results of our study suggest that MAIT cells are involved in the pathogenesis of AS.
MATERIALS AND METHODS
Subjects
Our study included 30 patients diagnosed with AS [28 men and 2 women, median age (interquartile range; IQR) 34.5 yrs (29.8–46.0)] and 21 HC [20 men and 1 woman, median age 34.0 yrs (31.5–37.0)]. The diagnosis of AS was established according to the modified New York criteria25. The HC did not have a history of any autoimmune disease and had never received immunosuppressive therapy. Informed consent was obtained from all patients with AS and HC according to the ethical guidelines for human subject research. The characteristics of the study participants are shown in Table 1. We used the ASDAS to evaluate the activity of AS26.
Clinical characteristics and laboratory test results of patients with AS and HC. Values are n or median (interquartile range).
Flow cytometry
Fresh peripheral blood mononuclear cells (PBMC) were isolated from heparinized blood by density-gradient centrifugation using Lymphoprep (Axis-Shield PoC AS). The cells were then stained with the combinations of the following monoclonal antibodies against human cell-surface antigens and a tetramer for 30 min on ice: anti-CD3-V500 (BD Biosciences), anti-CD4-APC-H7 (BD Biosciences), anti-CD8b-ECD (Beckman Coulter), anti-TCR-γδ-FITC (Beckman Coulter), anti-CD161-Brilliant violet-421 (BioLegend), anti-Vα7.2-PE (BioLegend), anti-CD69- Brilliant violet 605 (BioLegend), and CD1d tetramer loaded with PBS-57 (NIH Tetramer Core Facility). MAIT cells were CD3+γδTCR− Vα7.2TCR+CD161high, γδT cells were CD3+γδTCR+, invariant CD1d-restricted natural killer (NK) T cells were CD1d/PBS-57 tetramer+CD3+, and NK cells were CD3–CD56+. For intracellular active caspase-3 staining, PBMC were stained using surface markers. Thereafter, intracellular active caspase-3 staining was performed using the Active Caspase-3 Apoptosis Kit (BD Biosciences). Data were acquired on FACS LSR Fortessa (BD Biosciences) and the percentages of each cell population and mean fluorescence intensity (MFI) were analyzed using FlowJo software (Tree Star).
Intracellular cytokine staining
PBMC (2 × 106 cells/well in 96-well culture plates) were stimulated with 50 ng/ml phorbol-myristate-acetate (PMA; Sigma) and 500 ng/ml ionomycin (IM; Sigma) for 3 h at 37°C in 5% CO2. Golgi Plug Protein Transport Inhibitor (BD Biosciences) was added for the last 2 h of the culture at a final concentration of 1.0 μg/ml. After staining of the cell-surface antigens, intracellular cytokines were stained using the BD Cytofix/Cytoperm Fixation/Permeabilization Solution Kit (BD Biosciences) and Alexa Fluor 700 anti-IL-17A, PE-Cy7 anti-interferon-γ (IFN-γ), Alexa Fluor 700 anti-IL-2, PE-Cy7-anti-TNF-α, or their isotype control antibodies. Cells were analyzed on a FACS Fortessa flow cytometer (BD Biosciences) and data were analyzed using FlowJo software (Tree Star).
Statistical analysis
All data were analyzed using GraphPad Prism (GraphPad), and differences between the groups were analyzed using the Mann-Whitney U test. The significance level was set at p < 0.05. Associations between 2 variables were analyzed using Spearman correlation.
RESULTS
Reduced MAIT cell frequency in the peripheral blood of patients with AS
AS is chronic inflammatory arthritis characterized by the absence of specific autoantibodies and the contribution of innate immune cells to its pathogenesis. To reveal the involvement of innate-like lymphocytes (abundant in humans) in AS, we first examined the frequency of MAIT cells, γδT cells, and NK cells in the peripheral blood of patients with AS and HC by using flow cytometry. MAIT cells were identified as CD3+γδTCR–Vα7.2TCR+CD161high cells, γδT cells as CD3+γδTCR+ cells, and NK cells as CD3–CD56+ cells. In peripheral blood, the frequency of MAIT cells was significantly lower in patients with AS (median 1.62%, IQR 0.70–3.96) compared with HC (median 5.27%, IQR 3.18–7.49; Table 2 and Figure 1B). Among MAIT cells, there was no difference in the populations of CD4−CD8− double-negative (DN) cells or CD8+ cells between patients with AS and HC (Figure 1C). In addition, the frequency (median, IQR) of γδT cells (HC 2.40%, 1.60–4.32 vs AS 2.02%, 1.39–3.74) and NK cells (HC 12.10%, 9.46–17.55 vs AS 8.64%, 6.87–14.18) among total lymphocytes was not significantly different between patients with AS and HC (Table 2 and Figure 1D, 1E). These results indicated that the frequency of MAIT cells, but not γδT cells and NK cells, was lower in the peripheral blood of patients with AS compared with HC.

Frequency of innate-like lymphocytes in the peripheral blood of patients with AS and the HC. The frequency of innate-like lymphocytes in HC (n = 21) and patients with AS (n = 30) was analyzed by flow cytometry. A. Representative Vα7.2 TCR and CD161 staining gated on CD3+γδTCR– cells from 1 HC and 1 patient with AS. MAIT cells were identified as CD3+γδTCR–Vα7.2TCR+CD161high cells. Values in graphs indicate percentages of MAIT cells among αβT cells. B. Frequency of MAIT cells among αβT cells. C. Frequency of CD8+ and DN cells among MAIT cells. D. Frequency of γδT cells among lymphocytes. E. Frequency of NK cells among lymphocytes. Each symbol represents the value of 1 individual. Box indicates the first and third quartile. Line in the box indicates the median. Whiskers indicate the minimum and maximum. *** p < 0.001 (Mann-Whitney U test). AS: ankylosing spondylitis; HC: healthy controls; TCR: T cell receptor; MAIT: mucosal-associated invariant T; DN: double negative; NK: natural killer.
Frequency of innate-like lymphocytes. Values are median (interquartile range) unless otherwise specified.
Active caspase-3–positive MAIT cells are not increased in the peripheral blood of patients with AS
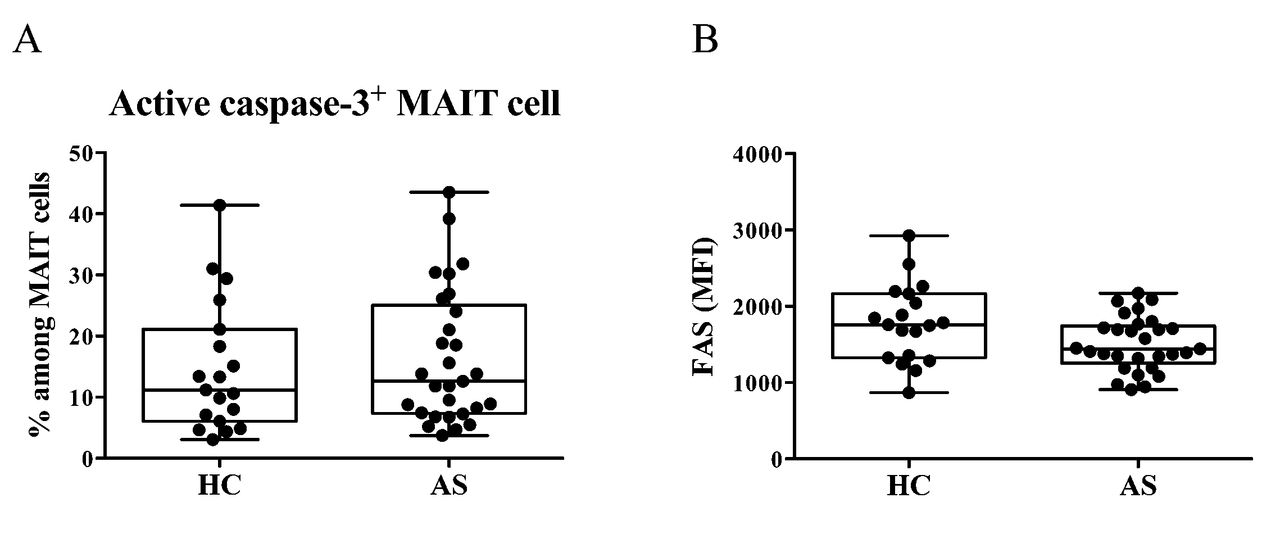
To determine the cause of the reduced number of MAIT cells in the peripheral blood of patients with AS, we examined whether MAIT cells underwent cell death in vivo. Peripheral blood MAIT cells were stained for intracellular active caspase-3. There was no difference in the frequency of active caspase-3–positive MAIT cells between HC and patients with AS (HC 11.20%, 6.05–21.10 vs AS 12.60%, 7.36–25.05; Appendix 1A). In addition, the expression level of FAS on MAIT cells was similar between HC and patients with AS (MFI: HC 1759, 1324–2166 vs AS 1442, 1255–1742; Appendix 1B).
Enhanced cytokine production in MAIT cells from patients with AS
Because MAIT cells efficiently produce high levels of inflammatory cytokines, we next assessed whether cytokine production from MAIT cells was altered in patients with AS. We performed intracellular staining of cytokines, including IL-17, IL-2, TNF-α, and IFN-γ in MAIT cells after activation with PMA/IM. Although the frequency of MAIT cells that produced IL-2, TNF-α, and IFN-γ did not differ between patients with AS and HC, the population of MAIT cells that produced IL-17 was higher in patients with AS than in the HC (Figure 2). These results suggest a functional difference in MAIT cells between HC and patients with AS.
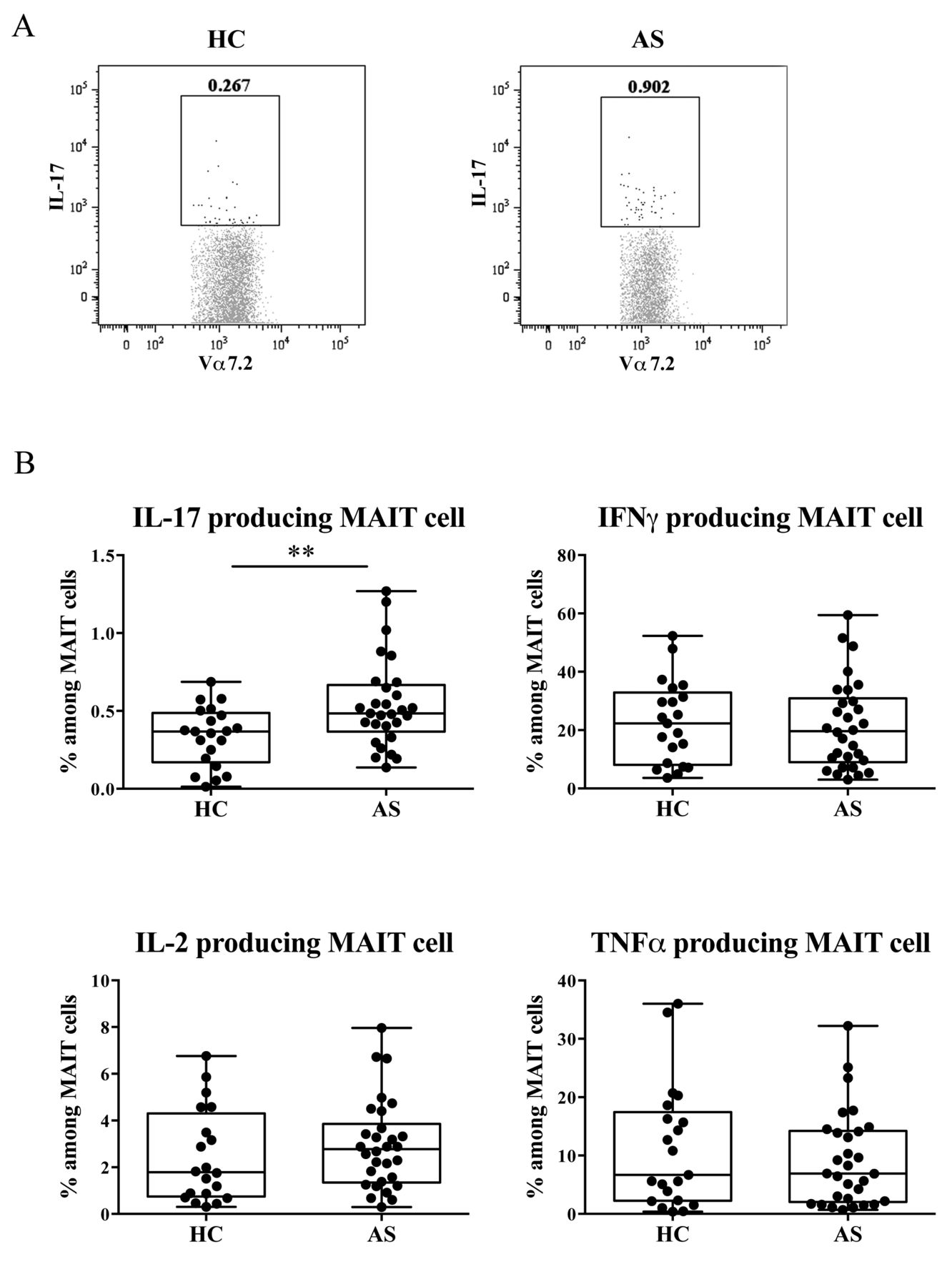
Frequency of cytokine-producing MAIT cells in patients with AS and HC. A. Representative staining profiles of IL-17 by MAIT cells from 1 HC and 1 patient with AS. B. The frequency of cytokine-producing MAIT cells was analyzed by flow cytometry in the HC (n = 21) and patients with AS (n = 30). MAIT cells producing IL-17 (IL-17+IFN-γ−), IFN-γ (IL-17−IFN-γ+), TNF-α (TNF-α+IL-2−), and IL-2 (TNF-α-IL-2+) are shown. Each symbol represents the value of 1 individual. Box indicates the first and third quartile. Line in the box indicates the median. Whiskers indicate the minimum and maximum. ** p < 0.01 (Mann-Whitney U test). MAIT: mucosal-associated invariant T; AS: ankylosing spondylitis; HC: healthy controls; IL-17: interleukin 17; IFN-γ: interferon-γ; TNF-α: tumor necrosis factor-α.
MAIT cell activation reflects AS disease activity
Because enhanced cytokine production from MAIT cells suggests that they are activated in patients with AS, we next examined the surface expression of CD69 on MAIT cells. The frequency of CD69+ MAIT cells and the MFI of CD69 on MAIT cells were associated with the ASDAS (Figure 3). These results indicated that MAIT cells were activated in vivo, and that the activation status reflected the disease activity in patients with AS.
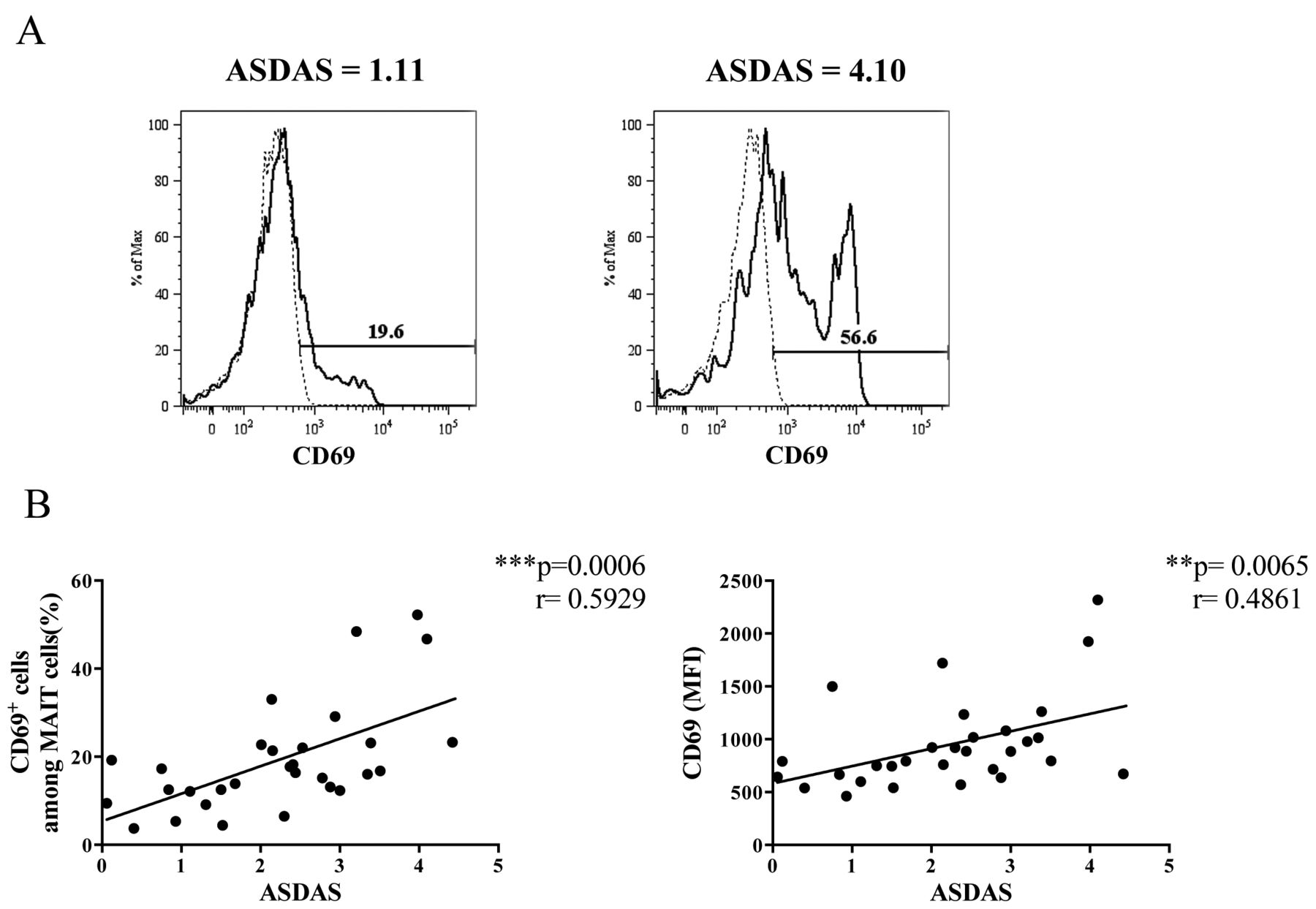
The correlation of activation status of MAIT cells with ASDAS. A. Representative staining and gating strategy of CD69 on MAIT cells from 1 patient with AS with ASDAS, 1.11 (left panel), and another patient with ASDAS, 4.10 (right panel). Dashed lines indicate mouse IgG1κ isotype control staining and black lines indicate CD69 staining on MAIT cells. The percentages of CD69+ cells among MAIT cells are shown. B. The correlation of ASDAS with the frequency of CD69-positive MAIT cells (left panel) and with MFI of CD69 expression on MAIT cells (right panel) was examined using Spearman correlation. MFI of CD69 on MAIT cells was measured using FlowJo software. Each symbol represents the value of 1 individual. MAIT: mucosal-associated invariant T; ASDAS: Ankylosing Spondylitis Disease Activity Score; MFI: mean fluorescence intensity.
Minimal effect of anti-TNF-α therapy on MAIT cell frequency and IL-17 production in patients with AS
In our study, 50% of patients with AS received anti-TNF-α therapy. To determine whether the findings observed in MAIT cells from patients with AS were caused by the anti-TNF-α therapy, we compared the frequency and cytokine-producing capacity of MAIT cells from patients with AS, shown in the experiments for Figure 1, Figure 2, and Figure 3, grouped by the administration or lack of anti-TNF-α therapy. As shown in Figure 4A, the frequencies of MAIT cells, CD8+MAIT cells, and DN MAIT cells were comparable between patients with AS regardless of whether they received anti-TNF-α therapy. The production of cytokines by MAIT cells including IL-17, IL-2, and IFN-γ were also comparable between patient groups, except for TNF-α (Figure 4B). We also investigated the relationship between CD69 expression on MAIT cells and disease activity. There was a tendency for a positive correlation between the activation status of MAIT cells and disease activity (Figure 4C). These findings suggest that numerical and functional changes and altered activation status of MAIT cells might be caused by AS pathology rather than anti-TNF-α therapy.

Effects of anti-TNF-α therapy on MAIT cells in patients with AS. (A) Frequency of MAIT cells among αβT cells and (B) cytokine-producing MAIT cells in patients with AS receiving (n = 15) and not receiving anti-TNF-α therapy (n = 15). Box indicates the first and third quartile. Line in the box indicates the median. Whiskers indicate the minimum and maximum. * p < 0.05 (Mann-Whitney U test). (C) The correlation of ASDAS with the frequency of CD69-positive MAIT cells (upper panels) and with MFI of CD69 expression on MAIT cells (lower panels) in patients with AS receiving (right panels) and not receiving (left panels) anti-TNF-α therapy was examined using Spearman correlation. Each symbol represents the value of 1 individual. ** p < 0.01. *** p < 0.001. Anti-TNF-α: antitumor necrosis factor-α; MAIT: mucosal-associated invariant T; AS: ankylosing spondylitis; ASDAS: Ankylosing Spondylitis Disease Activity Score; MFI: mean fluorescence intensity; DN: double negative; IL-17: interleukin 17; IL-2: interleukin 2; IFN-γ: interferon-γ.
DISCUSSION
In our study, we found that the frequency of MAIT cells in the peripheral blood of patients with AS was lower than that in HC. Only the frequency of MAIT cells, among all innate-like lymphocytes abundantly present in the peripheral blood, was significantly reduced in patients with AS. A study by Cho, et al reported that the reduction of MAIT cell frequency was not significant in patients with AS15. Another study showed a tendency for decreased MAIT cell frequency in patients with spondyloarthritis (SpA), including AS27. Although the latter study demonstrated an increase of CD8+ cells and a decrease of DN cells among MAIT cells in patients with SpA, we did not observe these alterations in the frequencies of MAIT cell subpopulations in patients with AS. The reason for these discrepancies among the studies remains unclear. Differences in the population groups and the disease duration might be responsible for the inconsistent findings among these groups. A reduction in the number of MAIT cells in the peripheral blood was also observed in patients with RA as well as other autoimmune disorders, such as multiple sclerosis (MS) and systemic lupus erythematosus13,15. In addition, reduced numbers of peripheral blood MAIT cells were observed in inflammatory, metabolic, or infectious diseases, such as inflammatory bowel diseases (IBD), obesity, type 2 diabetes, human immunodeficiency virus infection, and tuberculosis17,18,19,21,28,29,30. These findings suggest that MAIT cells are involved in various inflammatory conditions. Further, MAIT cells are present in inflamed tissues in diseases such as MS, IBD, and tuberculosis17,18,19,21,31,32.
Previously, we demonstrated that MAIT cells are reduced in the peripheral blood, but accumulate in the inflamed mucosa of patients with ulcerative colitis (UC)19. Because of the limited accessibility of the affected axial skeleton and intestinal tissue for analyses, we were unable to determine whether MAIT cells accumulated in the inflamed tissues in patients with AS. Alternatively, we examined whether the apoptosis of MAIT cells was enhanced. Based on the comparable expression of active caspase-3 and FAS by MAIT cells between patients with AS and HC, the reduced peripheral blood frequency of MAIT cells in patients with AS is unlikely to be caused by enhanced cell death. Rather, the reduction of MAIT cell frequency in the peripheral blood might be caused by their accumulation in the inflamed tissues of patients with AS. However, because apoptotic cells are cleared and do not remain in circulation, MAIT cells might be lost from the peripheral blood because of cell death. To uncover the involvement of MAIT cells and the cause of their reduced frequency, MAIT cells should be investigated in the inflamed tissues of patients with AS.
We previously reported that MAIT cells are effector cells in murine models of arthritis14. To determine whether the effector functions of these cells differ between arthritis and AS, we examined cytokine production from MAIT cells in response to stimulation with PMA/IM. Because other groups in addition to ours have shown that human MAIT cells produce inflammatory cytokines and not antiinflammatory cytokines, we investigated the production of these cytokines by MAIT cells in patients with AS11,13,17,20. The number of IL-17–producing cells was increased significantly in patients with AS. The production of IL-17 by MAIT cells was also enhanced in CD and RA, indicating that the augmentation of IL-17 production represents the activation status of MAIT cells15,17. Moreover, the expression of the activation marker CD69 on MAIT cells correlated with AS disease activity. Human peripheral blood MAIT cells display an effector memory phenotype (CD45RA–CD45RO+ CD62Llo-CD122intCD127hi), and they upregulate the expression of the early activation marker CD69 upon activation11,21,24,29,30,31. We and others reported that the increased expression of CD69 on MAIT cells is associated with elevated cytokine production in patients with UC and obesity19,28. The expression levels of CD69 on MAIT cells were positively correlated with clinical and endoscopic disease activity in patients with UC. These findings support the idea that MAIT cells might be involved in inflammatory conditions, including AS.
In addition to IL-17, a promising therapeutic target of AS33, inflammatory cytokines including TNF-α are produced by MAIT cells. TNF-α blockers are effective in patients with AS whose disease remains active despite the use of at least 2 nonsteroidal antiinflammatory drugs over 4 weeks or 1 local corticosteroid injection34. The potent ability of MAIT cells to produce inflammatory cytokines, which are therapeutic targets of AS, suggests that these cells might contribute to the pathogenesis of AS through cytokine production.
Two mechanisms that underlie the activation of MAIT cells have been identified. MR1-restricted antigens originating from the vitamin synthesis pathway in microbes activate MAIT cells24. Similar to other innate-like lymphocytes, MAIT cells can be activated by cytokines such as IL-12 and IL-18 in the absence of exogenous antigens31,35,36. Elevated IL-12 levels37 and abnormalities in the intestinal microbiota38,39 were reported in AS and might be associated with disease activity. We showed that CD69 expression on MAIT cells was related to the severity of AS. Therefore, MR1 antigens produced by changes in the composition of the intestinal microbiota or inflammatory cytokines might activate MAIT cells in patients with an active disease.
We showed that MAIT cells are activated but have a reduced frequency in the peripheral blood of patients with AS. Although our present findings suggest the involvement of MAIT cells in AS, further studies to determine the mechanisms underlying the contribution of MAIT cells to disease development and progression will lead to a better understanding of AS pathogenesis. To examine the contribution of MAIT cells in AS, these findings need to be investigated in a longitudinal study with a larger cohort of patients and in relation to AS clinical features such as HLA-B27 positivity and the type of organ involvement. Studies using animal models including HLA-B27 transgenic mice are also informative to understand the involvement of MAIT cells in AS.
Acknowledgment
We thank the National Institutes of Health Tetramer Core Facility (contract: HHSN272201300006C) for the provision of CD1d tetramers.
APPENDIX 1.

Frequency of active caspase-3–positive MAIT cells and FAS expression on MAIT cells from patients with AS and HC. A. The frequency of active caspase-3–positive cells among MAIT cells in the peripheral blood of HC (n = 20) and patients with AS (n = 29) was analyzed by flow cytometry. B. MFI of FAS (CD95) on MAIT cells was analyzed in the same HC and patients with AS (Mann-Whitney U test). Each symbol represents the value of 1 individual. Box indicates the first and third quartile. Line in the box indicates the median. Whiskers indicate the minimum and maximum. MAIT: mucosal-associated invariant T; AS: ankylosing spondylitis; HC: healthy controls; MFI: mean fluorescence intensity;.
Footnotes
Supported by the Japan Society for the Promotion of Science (Grant-in-Aid for Scientific Research [B] 26293234 to S.M.).
- Accepted for publication May 27, 2016.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}